單嘧磺酯鈉誘導芥菜型油菜雄性不育的作用效果
2021-01-27 07:00:32杜春蕾高虎虎馬懷英胡勝武
西北農業學報 2021年2期
杜春蕾,高虎虎,馬懷英,胡勝武,郭 媛
(西北農林科技大學 農學院,陜西楊凌 712100)
油菜是重要的經濟作物之一,含有豐富的油脂和蛋白質,是食用油和工業油的重要來源[1]。芥菜型油菜(BrassicajunceaL.,2n=36,AABB)由二倍體親本白菜(BrassicarapaL.,2n=20,AA)與黑芥(BrassicanigraL.,2n=16,BB)自然雜交后異源多倍化進化而來[2]。芥菜型油菜生育期長、成熟期晚,主根發達且入土深,具有天然黃籽、抗旱、耐瘠等優點,且芥菜型油菜所榨菜油香味濃郁,深受消費者喜愛[3]。由于極端災害天氣頻發,抗逆性的好壞對油菜品種來說格外重要[3],芥菜型油菜抗逆性較強,是非常具有應用潛力的種質資源。與其他大田作物相比,油菜雜種優勢顯著,利用雜種優勢是提高油菜產量的重要途徑[4]。中國油菜雜種優勢利用的研究起步較早,雜交油菜的選育和應用都處于世界先進水平。油菜雜種優勢利用途徑主要包括細胞質雄性不育(Cytoplasmic male sterility,CMS)、細胞核雄性不育(Genic male sterile,GMS)、生態型不育(Ecological male sterility,EMS)、自交不親和(Self incompatibility,SI)、化學雜交劑(Chemical hybridization agent,CHA)誘導的雄性不育[4]。其中,化學雜交劑誘導的雄性不育又稱化學殺雄,相對于其他雜種優勢利用途徑,化學殺雄具有兩系育種、組合自由、可多代利用及育種周期短的優點[5]。
自20世紀50年代初報道了馬來酰肼(植物生長調節劑,Maleic Hydrazide)可誘導玉米產生雄性不育后[6],化學雜交劑的研究飛速發展,接連發現NAA(萘乙酸,Naphthaleneacetic acid),2,4-D(2,4-二氯苯氧乙酸,2,4-Dichlorophenoxyacetic acid)等植物生長調節劑可誘導不同作物雄性不育[7-8]。但這些植物生長調節劑大多誘導效率低,對植株的生長和雌蕊的發育有負面效應,無法應用于生產實踐[9]。1970年后,相繼篩選出殺雄劑1號[10]、殺雄劑2號[11]、TO3[12]、殺雄劑3號[13]等用于水稻的化學雜交劑,黃雪清等[13]對不同水稻品種施用殺雄劑3號,90%的供試品種殺雄效果在97.2%以上。小麥中也篩選出RH531[14]、WL84811[15]、SQ-1[16]、GENESIS[17]等殺雄效果不錯的化學雜交劑,高慶榮等[17]使用GENESIS誘導小麥,殺雄效果達到95%,且具有一定的廣譜性。Song等[16]發現SQ-1對不同小麥品種的殺雄效果均到達95%以上。目前,油菜上公開報道的化學殺雄劑主要有甲基坤酸鹽類(殺雄劑1號[18],殺雄劑2號[19])、KMS-1[20],磺酰脲類(苯磺隆[21]、酰嘧磺隆[22]、氯磺隆[23]、EXP[24]、EN[25]、SX-1[26]、ESP[27]等),咪唑啉酮類[23](咪草煙、甲氧咪草煙),以及磺酰胺類[23](氟唑磺隆)等。芥菜型油菜雜種優勢的主要利用途徑是細胞質雄性不育(Cytoplasmic male sterility,CMS)[28]。印度學者報道了較多適用于印度的CMS[29],國內應用的不育胞質類型主要是hau CMS[30],由于可利用的芥菜型油菜不育種質單一,芥菜型油菜的雜種優勢利用受到了限制。化學雜交劑制種可以克服不育系對基因型的依賴性,是解決芥菜型油菜不育種質單一的有效途徑。Singh等[31]研究表明2% Surf Excel可誘導芥菜型油菜100%不育,且對產量影響較小,但由于缺乏后續研究,未能廣泛應用。Yu等[22]發現酰嘧磺隆可誘導芥菜型油菜雄性不育,但育性不穩定。井苗[32]以0.3μg/mL濃度的苯磺隆溶液處理芥菜型油菜可誘發100%全不育,但育性在后期不穩定。在眾多類型的化學雜交劑中,由磺酰脲類除草劑研發得到的化學殺雄劑效果好、制種產量高且成本低廉。磺酰脲類化合物作用于植物乙酰乳酸合成酶(ALS),通過抑制支鏈氨基酸的合成來抑制細胞分裂。動物體內不存在該化合物的靶標酶,因此磺酰脲類除草劑低毒、高效。單嘧磺酯鈉(Monosulfuron Ester Sodium,MES)是由李正名院士課題組研發的磺酰脲類除草劑,該藥是中國創制的具有自主知識產權的除草劑新品種[33]。Cheng等[34-35]研究發現0.05~0.10μg/mL MES可較好地誘導甘藍型油菜雄性不育,其機理研究表明用MES處理甘藍型油菜后得到的差異表達蛋白能夠引起代謝紊亂,從而引起絨氈層發育異常和小孢子不育。Li等[36]利用0.1μg/mL的MES誘導甘藍型油菜雄性不育后進行轉錄組分析,發現大部分差異表達基因定位于質體和線粒體中,且主要富集于糖類、脂質代謝、細胞運輸等通路。
本試驗以3份不同芥菜型油菜自交系為材料,對MES的殺雄效果及其農藝性狀的影響進行研究,旨在找到MES殺雄有效質量濃度,并通過細胞學觀察來初步揭示MES化學殺雄的機理。
1 材料與方法
1.1 材 料
供試的芥菜型油菜自交系材料共3份,分別為關中芥、L2598、芥菜野生型,以上材料均為西北農林科技大學胡勝武老師課題組收集。試驗于2016-2017年在西北農林科技大學農作物標本區(34°16′N,108°E)進行。利用隨機區組進行設計,分別設置3個重復,每小區包括4行,每行大約20棵植株,行長為2 m,行距為0.50 m,株距為0.10 m。土壤肥力中等,按照油菜大田常規管理,并于2017年5月收獲。
1.2 噴藥處理
供試除草劑原藥單嘧磺酯鈉(Monosulfuron Ester Sodium,MES,C14H13N4O5SNa),由南開大學李正名院士惠贈。稱取0.01 g MES,加入200 mL蒸餾水,配成200 mL 50 mg/L的MES母液存于4 ℃。取50 mg/L的母液0.75 mL、1 mL分別配置500 mL的0.075 mg/L、0.1 mg/L的藥液。第1次噴藥在油菜現蕾期最大花蕾長度為1~2 mm時,用小型手動噴霧器將MES均勻噴施于葉面,單株平均用藥量為8~10 mL。第2次噴藥在第1次噴藥后7~10 d進行。同時設置清水對照。
1.3 農藝性狀的調查與統計分析
盛花期對各株系各濃度材料育性進行考察,噴藥處理后在盛花期每3 d調查1次育性變化。統計全不育、半不育和可育等各類植株數量。測定自交和異交結實率,評估雌蕊與雄蕊育性。記錄整個花期所持續的時間,分別包括初花期、盛花期和終花期。此外,分別在每個小區隨機采取10朵花,對其花器官特征,包括花冠大小、花瓣長和寬、雌蕊、花藥和花絲長度等進行測量;在成熟期分別在各小區內隨機抽取10棵植株進行考種,考察性狀分別包括:株高、分枝部位、一級分枝數、二級分枝數、主序長度、主序角果數、全株角果數、每角粒數、角果長度等產量相關性狀。
用Win-Excel軟件對芥菜型油菜育性、花器官、農藝性狀等數據進行處理,利用SPSS(version 23.0,IBM SPSS Statistics,Chinese)軟件對數據進行方差分析,使用Duncan’s多重比較法分析不同處理間的顯著性差異。
1.4 花粉活力和花藥敗育的細胞學觀察
在油菜初花期隨機采取對照和不同質量濃度(0.075 μg/mL、0.1 μg/mL)MES處理的植株主花序(L2598)并立即用卡諾氏液(乙醇∶冰醋 酸=6∶1)固定2 h,然后用分級乙醇系列(100%、95%、85%、75%和70%)脫水,每次約 1~2 h,在4 ℃下儲存,直至使用。用鑷子輕輕擠壓花藥,將花粉粒釋放到醋酸洋紅溶液中,利用OLYMPUS BX51(Olympus,Japan)顯微鏡觀察花粉活力以及花藥敗育時期[37]。
取對照和0.1 μg/mL MES處理的植株(L2598)主花序,將其花藥切成小塊,并立即浸入4 ℃的磷酸鹽緩沖液(PBS,0.1 mol/L,pH 6.8)中,將其浸入由4%戊二醛組成的溶液中直至使用。分別用PBS(0.1 mol/L,pH 6.8)沖洗5、10、15、20和30 min后,將標本置于1%四氧化鋨 (0.2 mol/L PBS,pH 6.8)中于4 ℃固定2 h。然后用上述PBS緩沖液再次洗滌樣品。然后,使用30%、50%、70%、80%、90%和100%梯度乙醇系列分別將花藥脫水15 min,然后在樹脂(倫敦樹脂公司,雷丁,英國)∶酒精為1∶1混合物中滲入過夜,之后再在純樹脂中兩次(分別為7 h和3 h)將樣品放入明膠膠囊中,裝滿樹脂并在60 ℃孵育48 h以進行聚合。使用Leica EM UC7超薄切片機(Leica,Nussloch,Germany)將半薄切片切成 1 μm厚的切片,用1%甲苯胺藍溶液染色,最后在OLYMPUS BX51顯微鏡明場視野下觀察拍照[38]。
2 結果與分析
2.1 MES誘導芥菜型油菜雄性不育效果
在殺雄試驗中,不同濃度的MES均能引起3個芥菜型油菜材料不同程度的不育。雖然3個株系在抗藥性上存在差異,但引起3個材料不育的濃度并不存在差異。表1結果表明,低質量濃度處理下L2598和芥菜野生型出現了可育花粉,自交結實率分別為7.26%和1.51%。高質量濃度處理下3個品種的結實率均為0,關中芥和芥菜野生型的不育株率達到100%,但是有較為嚴重的藥害并伴有死蕾現象,以關中芥最為嚴重,L2598的不育株率達到96.30%。
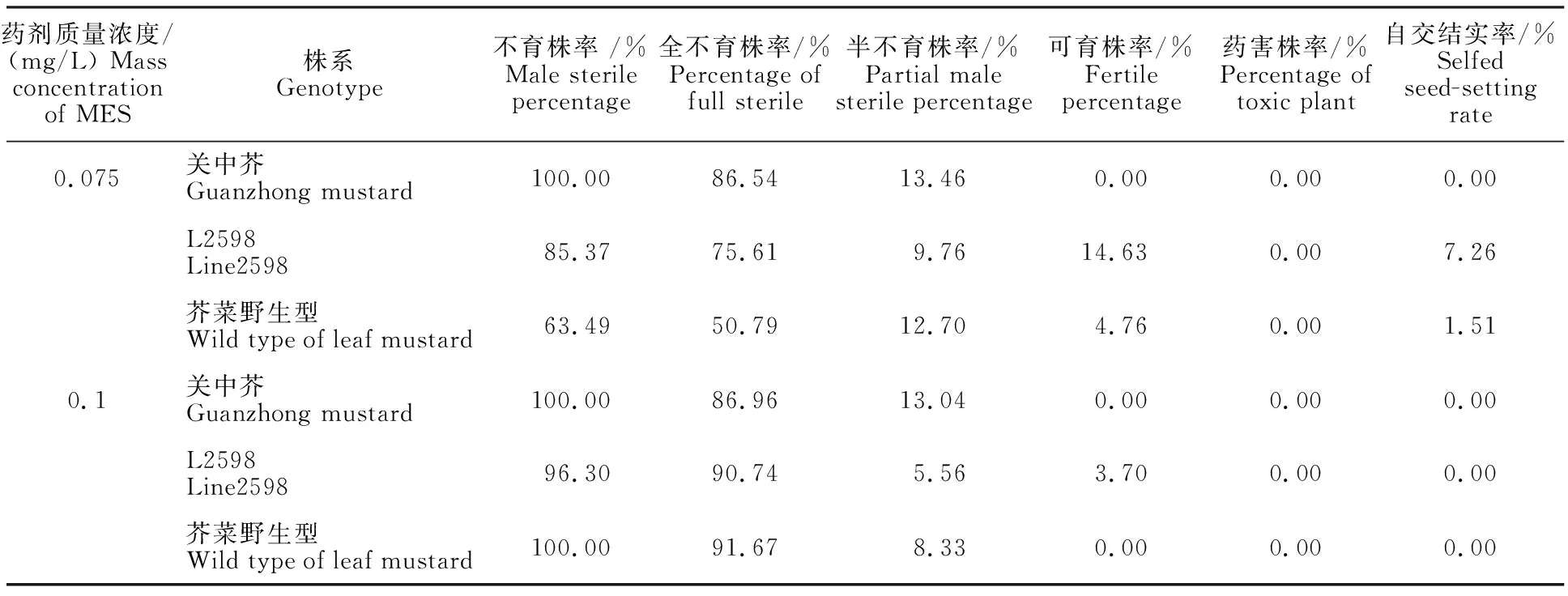
表1 MES對芥菜型油菜殺雄效果
2.2 MES誘導芥菜型油菜雄性不育對其農藝性狀的影響
化學雜交劑的篩選在于確立能夠誘導完全不育的最低濃度,從而將藥害降到最低。評估一個優良的化學雜交劑的標準主要包括兩方面,其一,能誘導完全不育;其二,對主要農藝性狀沒有影響或影響較小。
本試驗對MES各質量濃度處理后的L2598植株的農藝性狀進行了統計。如表2所示,低質量濃度(0.075 mg/L) MES處理后,L2598各農藝性狀與對照相比無顯著差異。高質量濃度(0.1 mg/L)處理下MES對L2598的全株角果數和角果長度有顯著影響,對每角粒數有極顯著的抑制作用,但是噴藥處理對株高、分支部位、分枝數、主花序長度和角果數沒有顯著影響。

表2 MES處理下芥菜型油菜主要農藝性狀的比較
2.3 MES誘導芥菜型油菜雄性不育對其植株生長發育的影響
優良的化學雜交劑應在保證不育的同時,應將對植物生長發育的影響降到最低。MES在引起不育的同時,對植物的生長發育也有影響。本研究發現,在有效殺雄濃度下L2598生長發育受到抑制。研究發現抽薹時,經MES處理的植株其節間的伸長受到抑制,且處理后,花期推遲,高濃度處理比低濃度處理花期推遲更多。
此外,噴藥處理還對花器官的形態產生較大影響。首先是噴藥處理后的主花序和雌蕊略微變短(圖1-A和1-B)。與可育材料雄蕊相比,不育花藥在早期發白,至后來逐漸干枯出現干癟、瘦小、有些成三角形。高濃度處理下雄蕊縮短,花絲不能伸長,花藥干癟,無可育花粉,花藥敗育很徹底(圖1-C)。其次噴藥處理還引起了雄蕊發白,萼片、花柄、花軸和花瓣褪色,以高濃度更為顯著(圖1-D)。噴藥處理還引起了(1)花蕾和莖稈發紫;(2)角果變短;(3)葉片不均勻泛黃,葉脈失綠,部分葉片卷曲,略有畸形(圖1-E)。
將MES對花發育的影響進行統計(表3),發現各項與花發育相關的指標均有顯著差異,經MES處理的油菜花器官的各個組成部分均顯著變小,以高質量濃度的更為顯著。
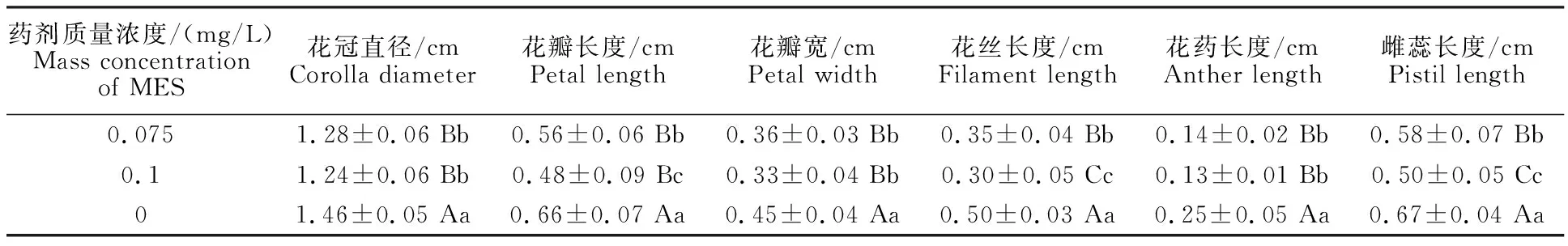
表3 MES處理下對芥菜型油菜花器結構特征的分析
2.4 MES誘導芥菜型油菜雄性不育的細胞學 觀察
2.4.1 壓片觀察 取L2598的花粉亞歷山大染色后進行壓片觀察,可育小孢子細胞核可被染成紫紅色,不育小孢子不能著色(圖2)。在0.075 mg/L和0.1 mg/L兩種MES質量濃度下都未見可育孢子,且不育孢子在體積上均小于對照(圖2-A,圖2-B,圖2-C)。在油鏡下進一步觀察發現不育小孢子表現出較為嚴重的細胞壁畸形,其中0.1 mg/L處理下尤為嚴重(圖2-D),畸形小孢子的數量相對更多。
為進一步探究MES誘導芥菜型油菜花粉不育的過程,對噴施清水的對照和0.1 mg/L MES處理植株花粉粒(L2598)用醋酸洋紅染色后進行壓片觀察(圖3)。醋酸洋紅染色后,可育小孢子為鮮紅色,不育小孢子呈鐵銹色或不能著色。壓片結果顯示在花粉母細胞時期(圖3-A和圖3-B),噴藥處理后的花粉粒已經出現異常。雖然MES處理過的花粉顆粒在細胞形態上完整,與對照無明顯差異,但細胞著色整體較淺。至四分體時(圖3-C和圖3-D),MES誘導的花粉顆粒出現部分四分體不能著色,且所有小孢子著色淺。四分體之后不育小孢子呈鐵銹色,可育小孢子為鮮紅色(圖3-E~J)。總的來說,MES處理后,小孢子早期細胞基本完整,晚期出現空的小孢子,且所有小孢子都出現一定的畸形。
2.4.2 半薄切片觀察 為探究MES引起芥菜型油菜敗育的機理,對噴施清水對照和0.1 mg/L MES處理的L2598全不育株花藥進行半薄切片試驗,用苯胺藍染色后置于光學顯微鏡下觀察。
半薄切片的結果顯示,在造孢細胞時期(圖4-A和圖4-B),噴藥處理與對照差異并不明顯,整體著色都較淺,處理的絨氈層并無異常。至花粉母細胞時期(圖4-C和圖4-D),MES處理后與對照相比著色顯著加深,此外在形態上二者差異不明顯,但在絨氈層細胞形態上存在差異。對照絨氈層緊貼內壁,形狀規則,而經MES處理的絨氈層輪廓不清晰。發育至四分體時期(圖4-E和圖4-F),MES處理出現很大差異。對照四分體著色很深且外圍胼胝質著色較淺,而MES處理過的四分體著色很淺,胼胝質相對深一些。MES處理的絨氈層幾乎不能被染色,細胞呈畸形狀態。至單核早期(圖4-G和圖4-H),對照和MES處理小孢子都已經釋放,絨氈層也已經開始降解,但MES處理的小孢子明顯染色較淺,且絨氈層幾乎沒有內容物。單核晚期(圖4-I和圖4-J),對照小孢子發育正常,染色較深,而MES處理的絨氈層完全降解、小孢子沒有內容物。
綜上所述,認為噴施單嘧磺酯鈉引起芥菜型油菜花粉敗育發生在四分體至單核晚期。觀察到噴施MES后,在四分體時期處理的絨氈層發育異常,這可能是引起小孢子敗育的主要原因。
3 討 論
化學雜交劑制種可以克服不育系對基因型的依賴性,是解決目前國內油菜應用不育胞質類型單一的重要途徑之一。在油菜雜交種實際生產中,開發更多高效、低毒、穩定,同時具有中國自主知識產權的化學雜交劑是目前化學殺雄育種技術的重要任務[30]。
中國早期開發的殺雄效果較好的赤霉素,殺雄效果達到70%~100%,但殺雄效果穩定性差;甲基砷酸鋅的殺雄效果為80%~100%[18-19],但該類CHAs含有毒砷元素。由磺酰脲類除草劑研發得到的化學殺雄劑EN[25]、ESP[27]、EXP[24]、SX-1[26]、苯磺隆[21]等在甘藍型油菜中的殺雄效果較好,不育率達到90%以上,而且具有成本低廉、低毒的優點,但大都包含受國外知識產權保護的成分。
目前,芥菜型油菜中磺酰脲類化學殺雄劑的相關研究較少,Yu等[22]發現酰嘧磺隆可以誘導芥菜型油菜雄性不育,但育性不穩定;井苗[32]發現0.3 μg/mL的BHL溶液可誘導芥菜型油菜100%全不育,但后期育性不穩定。本研究以3份芥菜型油菜自交系為材料,對現蕾期葉面噴施MES誘導芥菜型油菜雄性不育的作用效果進行了研究。試驗結果表明,單株受藥量在8~10 mL時,0.075 mg/L和0.1 mg/L兩個MES質量濃度梯度處理下均能有效誘導芥菜型油菜不同程度的不育。0.075 mg/L的MES可以誘導關中芥完全不育,0.1 mg/L MES處理后3個品種的不育株率可以達到96.30%~100%。與前期開發的多種化學雜交劑相比,MES的殺雄效率高、殺雄效果穩定,并且低毒、環保,擁有自主知識產權。
邢澤農[39]對MES誘導甘藍型油菜雄性不育進行了研究,結果發現0.1 μg/mL MES處理對株高、單株果粒數、角果粒數沒有顯著影響,0.2 μg/mL MES處理后植株株高顯著降低,單株角果數和全株粒數上升,角粒數變化不大。Yu等[40]發現葉面噴施0.10 μg/mL MES對甘藍型油菜的株高、角粒數有一定的負面影響,對單株角果數,全株粒數有一定的增加作用,但對單株產量沒有明顯影響,并且不同基因型油菜之間的反應存在差異。本研究中,MES處理對農藝性狀的影響因品種而異,0.1 mg/L質量濃度下,關中芥植株藥害大并伴有死蕾現象,而L2598的植株藥害輕,全株角果數呈上升趨勢,每角粒數降低,角果長度變短。Cheng等[34]研究發現噴施MES對甘藍型油菜植株的花瓣和雌蕊沒有受到明顯影響。本研究中,兩種濃度MES處理后的油菜植株花期均推遲,植株的花序和雌蕊略微變短;雄蕊花絲明顯變短,花藥變短、干癟;葉片隨MES處理濃度增加而變小。此外,發現MES處理后的植株花冠直徑、花瓣長度和寬度均顯著變小,高濃度處理下減小幅度更為顯著。這與Yu等[40]用苯磺隆處理后的甘藍型油菜所表現出的開花時間延遲,開花時間縮短,以及植物高度和花器官尺寸減少是一致的。
化學雜交劑誘導植株雄性不育研究中絨氈層液泡化、異常膨大、提前或是延遲解體等現象較為常見,且異常的小孢子細胞在花粉發育的各個時期不斷產生和積累,導致成熟期花粉全部敗育[24,32,41]。本研究選取0.01 μg/mL MES處理的L2598進行細胞學觀察,結果發現小孢子在花粉母細胞時期就已經出現異常,早期細胞基本完整,晚期逐步出現空的小孢子,且所有不育小孢子皆出現較為嚴重的細胞壁畸形;MES處理植株的的絨氈層在造孢細胞時期與對照差異不明顯,花粉母細胞時期二者開始存在輕微差異,絨氈層輪廓模糊,發育至四分體時期絨氈層細胞呈不規則膨大,單核早期時絨氈層幾乎沒有內容物且延遲降解。筆者認為噴施MES導致花粉敗育發生在四分體至單核晚期之間,在這段時期內不斷有敗育花粉產生,直至成熟期全部花粉敗育。化學雜交劑誘導的雄性不育是從多方面、多途徑來誘導敗育花粉不斷產生,最終導致雄性不育,本研究中植株絨氈層發育異常且延遲降解,可能是引起小孢子敗育的主要原因。
本研究旨在不影響芥菜型油菜農藝性狀的前提下找到較好殺雄效果的MES濃度。研究發現葉面噴施0.075 mg/L的MES可誘導關中芥完全不育,且對主要農藝性狀影響較少。0.1 mg/L的MES殺雄效果最好,但對農藝性狀的影響較顯著,關中芥死蕾現象嚴重,不宜在雜種優勢利用中推廣。任何一種化學殺雄劑都有適宜的使用濃度及其使用方法,油菜中磺酰脲類殺雄劑的有效殺雄濃度為0.02~20 μg/mL[21,24-27,31-32]。Cheng等[35]用MES在49個甘藍型油菜上試驗表明,單核期用0.05~0.10 μg/mL MES處理產生的不育株率為99.3%~100%。張寶娟等[42]對甘藍型油菜噴施0.075~0.1 μg/mL的苯磺隆得到94%~100%的不育株率。Yu等[21]發現苯磺隆、EXP[24]誘導甘藍型油菜雄性不育的最適質量濃度分別為0.2 μg/mL和0.25 μg/mL。劉絢霞等[25]認為甘藍型油菜用0.5~0.8 μg/mL EN處理殺雄效果最好,全不育株率在95%以上,不育株率為100%。嚴自斌等[27]研究表明0.3~ 0.6 μg/mL的ESP誘導甘藍型油菜雄性不育的效果最佳,不育株率達98%。本研究中MES誘導芥菜型油菜雄性不育的最適質量濃度為 0.075~0.1 μg/mL,較MES或其他磺酰脲類化學雜交劑誘導甘藍型油菜雄性不育的最佳殺雄濃度范圍更小,且細微濃度差異即可造成殺雄效果和農藝性狀的顯著差異,說明芥菜型油菜可能對MES更加敏感,而且不同品種自交系的芥菜型油菜對MES的敏感性存在差異。本研究探討了MES誘導芥菜型油菜雄性不育的作用效果,并對其不育花藥敗育的特征進行了觀察,從而為確定MES誘導芥菜型油菜雄性不育的最佳殺雄濃度提供了有價值的參考。
