基于轉錄組的漆樹MYB轉錄因子的篩選及分析
2021-01-26 06:51:42謝冬冬王武萍何學高黃曉華
西北林學院學報 2021年1期
謝冬冬,王武萍,何學高,黃曉華*
(1.西北農林科技大學 林學院,陜西 楊陵 712100;2.中華全國供銷合作總社 西安生漆涂料研究所,陜西 西安 710000)
轉錄因子是真核生物細胞內通過與特定基因啟動子結合以調控下游基因的表達的一類蛋白[1]。通常含有寡聚化位點(oligomerization site)、轉錄激活區(activation domain)、DNA結合區(DNA-binding domain)以及核定位信號區(nuclear localization signal)等功能區域[2]。植物中轉錄因子類型多樣,常見的有WRKY類、ERF類、MYB類等,而MYB類轉錄因子是植物轉錄因子中數量較多的家族之一,其功能多樣[3]。MYB-DNA結合結構域(MYB-binding domain)是一段含有50~53個氨基酸殘基的不完全保守肽段[4]。該結構域中分布有3個起疏水核心作用的相對保守的色氨酸殘基(彼此間隔18或19個氨基酸殘基),維持MYB轉錄因子的構型[5]。MYB轉錄因子通常含有1~4個串聯的MYB-DNA 結合結構域,依據MYB轉錄因子含有的結合結構域數目的差異可將其分為4類,R1-MYB、R2R3-MYB、R1R2R3-MYB和R4-MYB[6]。其中R2R3類轉錄因子中R3結構域的第一個色氨酸殘基通常被亮氨酸、異亮氨酸或苯丙氨酸等所取代[7]。在植物中發現的第一個MYB轉錄因子是玉米穗基因C1編碼的參與花青素代謝合成的C-MYB-like[8]。現發現MYB類轉錄因子對植物的整個生命過程都具有重要的意義,包括形態建成[9]、生長發育[10]、對生物和非生物脅迫的應答[11-12]以及植物初生和次生代謝等[13]。
漆樹(Toxicodendronvernicifluum)為漆樹科(Anacardiaceae)漆屬(Toxicodendron)落葉喬木或小喬木,屬亞熱帶區系,新生代第三紀古老孑遺樹種[14],雌雄同株或異株,變異類型多,歷經7 000多萬a的演化,彌足珍貴[15]。但由于漆樹種植的存活率偏低和過度利用,我國目前保存的漆樹資源稀少,下降趨勢明顯,發展前景堪憂[16]。生漆是漆樹乳汁道中所分泌的次生代謝產物,是一種優良的天然涂料,生漆的產生和貯存的主要部位是漆樹各器官的韌皮部,以主干樹皮韌皮部為主要合成部位[17]。M.Zhaoetal[18]用光學顯微鏡(LM)和透射電子顯微鏡(TEM)研究了漆樹分泌細胞的結構和原始分泌物,發現高爾基體可以產生帶有單層膜的分泌顆粒并將其運輸到質膜中與細胞膜融合,結合細胞擴散到漆汁道并聚集形成生漆。生漆的主要成分漆酚占生漆含量的40%~80%,屬類黃酮類物質[19]。植物中類黃酮類物質是苯丙酸合成途徑的一個分支,其合成途徑主要受一些酶基因和轉錄因子的調控,參與植物黃酮類化合物生物合成的轉錄因子主要有MYB、bHLH和WD40重復蛋白[20]。因此,研究漆樹漆酚生物合成路徑,篩選并鑒定調控漆酚代謝的轉錄因子,有利于對漆樹的進一步開發和利用。MYB轉錄因子參與植物次生代謝調控,對漆樹MYB轉錄因子進行研究,有助于了解漆酚這一漆樹次生代謝產物的合成及調控機制。
1 材料與方法
1.1 材料
漆樹材料采自西北農林科技大學漆樹園。漆樹MYB轉錄因子源于實驗室已構建的漆樹轉錄組數據庫(登錄號:PRJNA587830)。從擬南芥信息資源(TAIR)數據庫(https://www.arabidopsis.org)下載擬南芥MYB轉錄因子家族氨基酸序列。
1.2 漆樹MYB轉錄本篩選
高通量測序工作由北京組學生物科技有限公司完成,綜合Pacbio SMRT三代技術和IlluminaX Ten二代測序技術的優點獲得漆樹轉錄組,經初步篩選獲得89條MYB基因序列,氨基酸序列的獲取通過基因ID進行搜索,然后在ORF Finder在線軟件(https://indra.mullins.microbiol.washington.edu/sms2/orf_find.html)的輔助下分析預測開放閱讀框(ORF)并獲得具有全長氨基酸序列的漆樹MYB轉錄因子家族蛋白基因序列。最后,將上述含有MYB功能域的序列提交至PlantTFcat(http://plantgrn.noble.org/PlantTFcat/)進行比對分析,進一步驗證其是否屬于MYB類轉錄因子[21]。
1.3 漆樹MYB轉錄因子的分類與結構域基序分析
運用在線軟件SMART(http://smart.embl.de/)將驗證正確的MYB蛋白序列進行批量搜索并分類[22],分類標準為其所含有的DNA-binding domain的數目。利用MEGA 5.1對漆樹中的R2R3-MYB類蛋白序列進行比對并保存比對結果。刪除不保守區域,利用weblogo(http://weblogo.berkeley.edu/logo.cgi)對R2R3-MYB的DNA domain進行結構域特征分析[23]。
1.4 漆樹MYB轉錄因子保守基序分析
運用The MEME Suite4.12.0(http://meme-suite.org/tools/meme)程序分析漆樹MYB家族蛋白的基序[24],基序寬度最小值設定為6、最大值為50,基序數量設為10,其余參數為默認值。
1.5 漆樹MYB轉錄因子蛋白的結構分析
選擇具有全長氨基酸序列的漆樹 MYB 轉錄因子家族蛋白,利用在線工具ProtParam(http://web.expasy.org/protparam/)對蛋白進行一級結構和理化性質分析。蛋白二級結構特征采用在線軟件SOPMA進行分析(https://npsaprabi.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_sopma.html)[25]。
1.6 漆樹MYB轉錄因子的GO注釋分析
使用WEGO在線注釋軟件(http://wego.genomics.org.cn/)對篩選獲得的漆樹MYB 轉錄因子進行功能注釋分析[26]。
1.7 漆樹MYB轉錄因子家族進化分析
為進一步探究漆樹MYB轉錄因子的功能,運用MEGA 5.1軟件采用鄰近法(Neighbor-Joining)構建包含125個擬南芥MYB轉錄因子及45個漆樹MYB轉錄因子(其中有3條序列由于同源性相差太大,剔除)的系統進化樹。其中Model設置為p-distance,Bootstrap值設為1 000,其他參數為默認值。最后,用在線軟件iTOL (https://itol.embl.de/)對構建好的進化樹進行優化。
1.8 漆樹MYB轉錄因子表達分析
基于漆樹根、莖和葉3個組織的轉錄組數據,取MYB基因家族各基因FPKM(fragments per kilobase of transcript per million mapped reads)值,采用工具ClustVis(https://biit.cs.ut.ee/clustvis/)對48個MYB轉錄因子在各組織中的表達量進行分層聚類及表達模式分析[27]。
2 結果與分析
2.1 漆樹MYB家族成員的獲得
經初步篩選,從漆樹轉錄組數據庫中獲得89條MYB基因序列,片段大小在555~5 226 bp。通過ORF預測全長氨基酸序列,具有完整ORF的序列有48條,序列平均長度為1 344 bp。其中,最長的序列由5 226個堿基組成(PB_c0_g11314),共編碼1 742個氨基酸;最短的序列含555個堿基(PB_c0_g2870),編碼185個氨基酸。
2.2 漆樹MYB蛋白序列的分類與結構域特征分析
對獲得的48條漆樹MYB蛋白序列運用SMART進行批量搜索,將不含或含有不完整 MYB-DNA binding結構域的序列以及冗余轉錄本序列舍去,分類依據為其所含有的DNA binding domain的數目。分別得到1條R3-MYB 序列,21條R2R3-MYB序列和26條R1-MYB序列。R2和R3是MYB識別DNA序列所必需的,氨基酸出現頻率越大在圖中對應位點的字母長度越高,表明氨基酸殘基越保守[1]。利用weblogo在線分析軟件對漆樹中的R2R3-MYB類轉錄因子的DNA結合結構域進行特征分析(圖1),結果表明,在長度約為51個氨基酸殘基的R2結構域中含有3個高度保守的色氨酸殘基(分別位于第4位、24位和44位),且每個色氨酸殘基有18~19個氨基酸相間隔。總體來看,漆樹MYB的R2R3結構域序列同已鑒定出的擬南芥、玉米和森林草莓等的序列高度相似,均具有特征性的氨基酸,R2和 R3結構域中存在大量保守氨基酸殘基,這些保守氨基酸殘基可能在構成MYB蛋白螺旋-轉角-螺旋(HTH)結構以及特異的DNA結合位點方面起著重要作用。
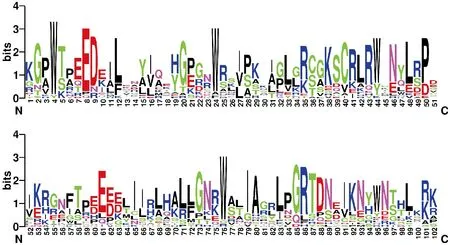
圖1 漆樹R2R3-MYB轉錄因子高度保守的DNA binding結構域Fig.1 Highly conserved DNA binding domain of T.vernicifluum R2R3-MYB transcription factors
2.3 漆樹MYB轉錄因子基序分析
通過MEME軟件分析了48個漆樹MYB轉錄因子的10個基序(圖2),結果表明,除PB_c0_g15467和PB_c0_g9179外,其他均具有基序1,具有基序2的有47個MYB蛋白。從漆樹MYB轉錄因子各自的基序及進化關系來看,同一進化分支的漆樹MYB蛋白保守基序種類相同或相近且位置大體一致,而不同進化分支中的漆樹MYB蛋白的保守基序和位置的差異則較大,同一進化分支的轉錄因子其功能可能相似。
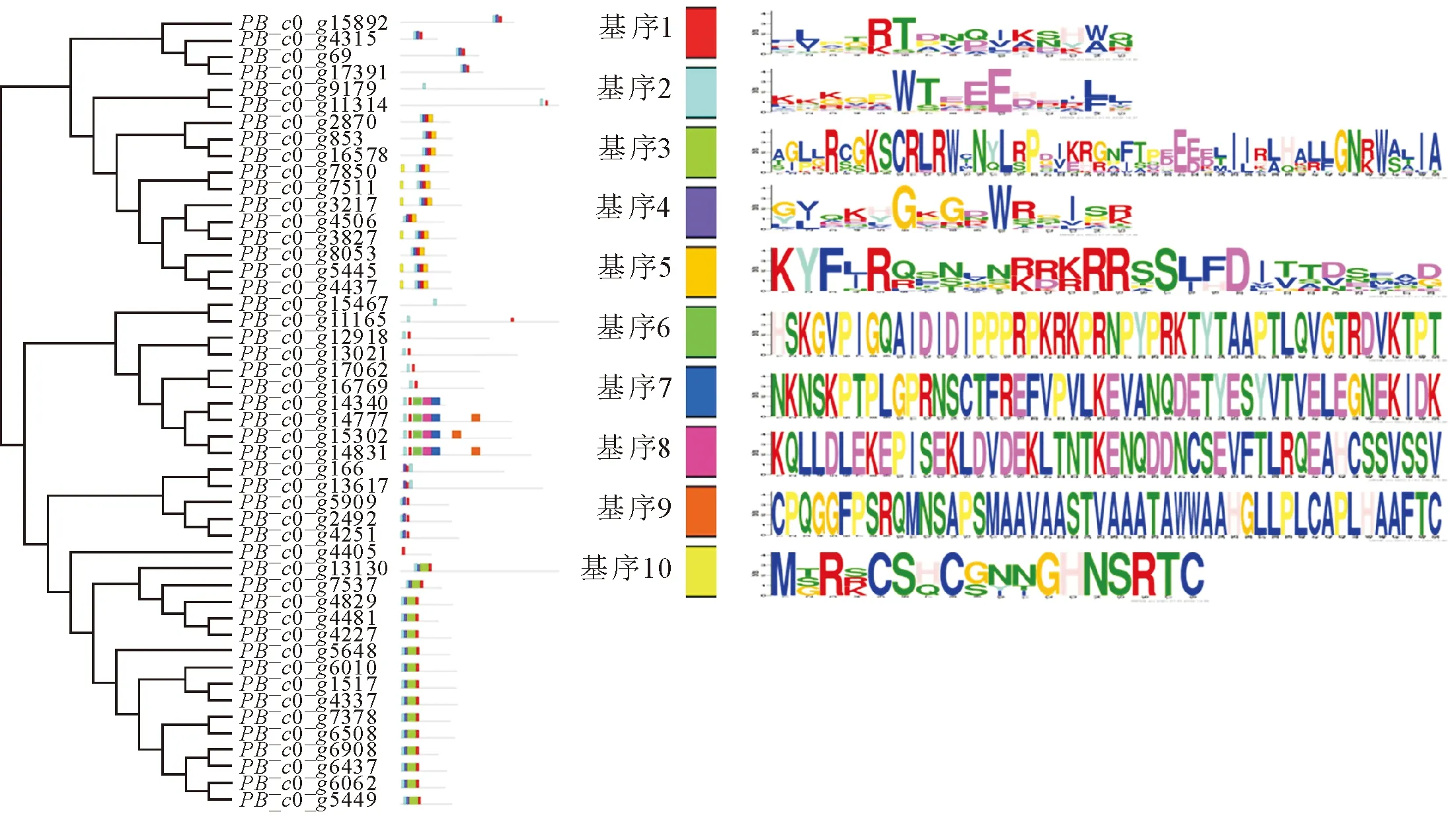
圖2 漆樹MYB轉錄因子基序分析Fig.2 Motifs analysis of T.vernicifluum MYB tanscription factors
2.4 漆樹MYB家族蛋白理化性質分析
ProtParam和SOPMA對漆樹MYB家族蛋白的理化性質分析結果(表1)表明,漆樹MYB家族蛋白平均氨基酸數目為447,最長的是PB_c0_g11314編碼蛋白,由1 741個氨基酸組成,最短的由184個氨基酸組成,即PB_c0_g4405編碼蛋白。48個漆樹MYB家族蛋白的平均相對分子質量為49 546.39。平均等電點(PI)為7.76,最大值為PB_c0_g4315編碼蛋白的10.05,最小值為PB_c0_g12918編碼蛋白的4.5,其中14個MYB家族蛋白平均等電點<7,偏酸性,剩余的34個等電點>7,偏堿性,說明漆樹MYB家族蛋白整體表現為偏堿性。漆樹MYB家族蛋白中,有44個蛋白為疏水性蛋白,平均疏水性值<-0.5,僅PB_c0_g8053、PB_c0_g7850、PB_c0_g2492和PB_c0_g4251編碼的蛋白>-0.5,具有一定的親水性。通過SOPMA對漆樹48個MYB家族蛋白進行二級結構預測,結果表明,在這48個MYB家族蛋白中,除PB_c0_g15467編碼蛋白外,其余47個均表現為無規則卷曲含量最高,其次是α螺旋和延伸鏈,β轉角的含量最低,故無規則卷曲和α螺旋是漆樹MYB家族蛋白二級結構的主要成分。
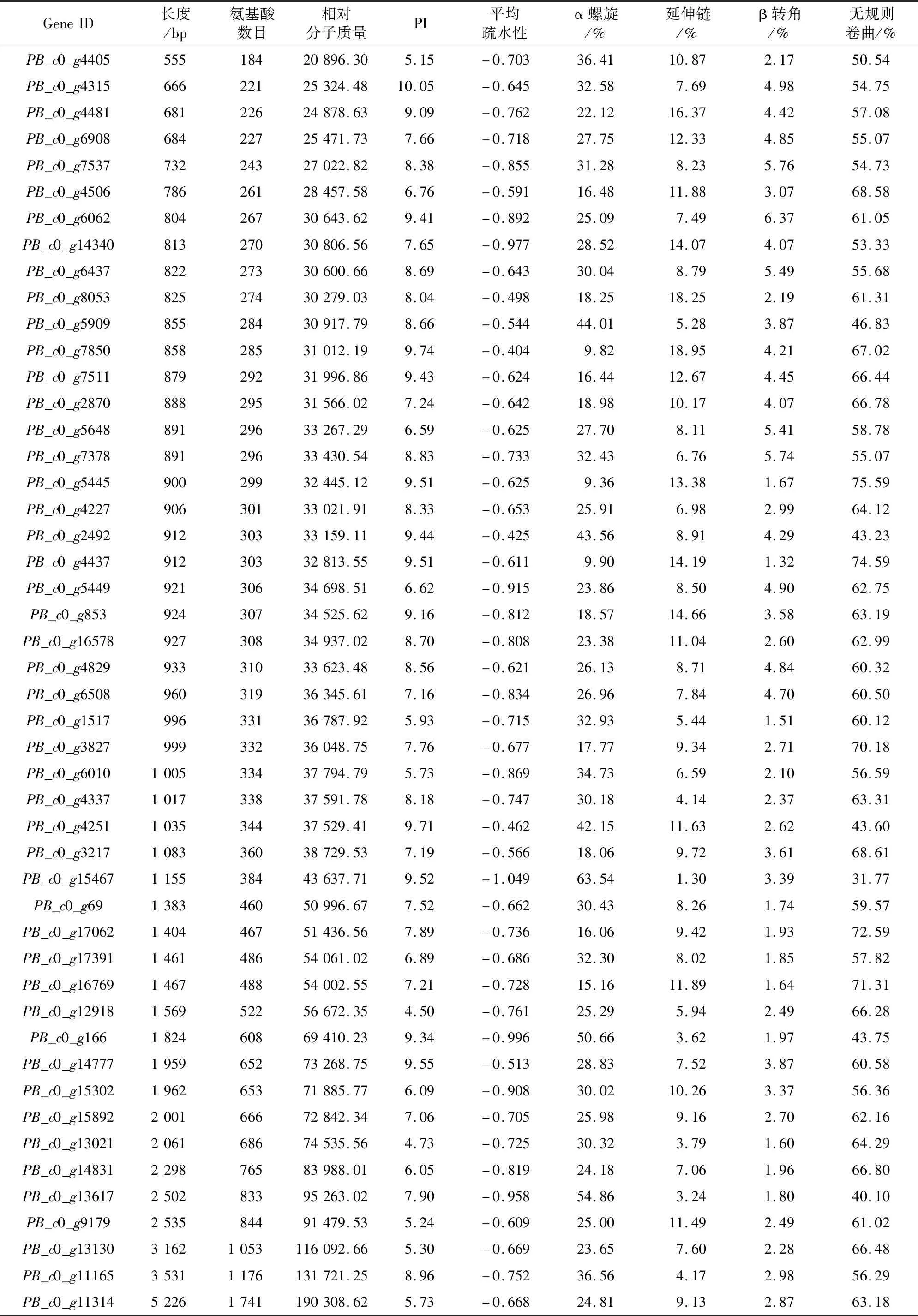
表1 漆樹MYB家族蛋白理化性質Table 1 Physicochemical properties of T.vernicifluum MYB family proteins
2.5 漆樹MYB轉錄因子的GO注釋分析
GO注釋分析表明(圖3),漆樹48條MYB轉錄因子序列富集為27個功能類別,分別有27、37條和29條注釋到細胞組分、分子功能和生物過程。細胞組分中,細胞部分(26個)、細胞(26個)和細胞器(26個)包含的序列較多,含蛋白質復合物(5個)和細胞器部分(4個)所含序列較少,膜部分、膜和膜封閉腔均只含有1個序列。分子功能中,具有結合活性的序列最多,為37個,其次為含12個序列的轉錄調節活性功能類別,而催化活性所含序列最少,僅有2個。生物過程中,代謝過程(27個)、生物調節過程(22個)、細胞過程(29個)、生物調節(22個)、刺激反應(11個)功能組中所含序列較多,細胞成分組織或生物發生(7個)、生物過程的正調節(4個)、免疫系統過程(2個)、信號(2個)、發育過程(2個)、生殖過程(2個)、再生產(2個)、多細胞生物過程(2個)、生物過程的負調節(4個)、節律過程(4個)、多細胞生物過程(2個)功能組中所含序列較少。從注釋結果來看,細胞部分在細胞組分中表現為主要富集,分子功能集中富集在結合活性上,生物過程主要富集在細胞過程。漆樹MYB轉錄因子發揮相應調控功能的方式是同 DNA 區域相結合,參與植物的細胞過程、代謝過程和生物調節等過程。
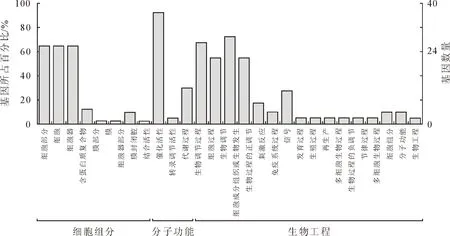
圖3 漆樹MYB轉錄因子功能注釋Fig.3 Function annotation of T.vernicifluum MYB tanscription factors
2.6 漆樹MYB轉錄因子的聚類分析
根據預測到的漆樹MYB蛋白和125個擬南芥MYB各亞家族蛋白質保守結構域氨基酸序列的相似性,系統發育樹利用MEGA 5.1采用Neighbor-Joining法進行構建。參考擬南芥MYB家族轉錄因子的亞家族分類方法[28],將漆樹MYB蛋白序列分為8個亞家族。由圖4可以看出,漆樹和擬南芥的部分MYB蛋白聚在同一亞組,高度同源。漆樹MYB家族蛋白中多數與擬南芥的MYB蛋白聚集在不同亞組,說明漆樹MYB家族蛋白與擬南芥MYB蛋白均具有較高的保守性。模式植物擬南芥R2R3-MYB家族蛋白中第S4、S5、S7亞族參與調控擬南芥的次生代謝產物生物合成過程,故與S4、S5、S7同一亞組的PB_c0_g6437、PB_c0_g6508和PB_c0_g7378、PB_c0_g4405、PB_c0_g12918和PB_c0_g13021編碼的漆樹MYB蛋白也可能參與調控漆樹的次生代謝過程。此外,有28個漆樹MYB轉錄因子單獨聚為1族,可能是在進化過程及環境選擇過程中產生了在序列上與擬南芥有較大差異的MYB轉錄因子,這些轉錄因子可能參與漆樹種某些特有的生物學過程的調控。漆酚是一種具有15~17個碳原子的不同飽和度長側鏈的單元酚、鄰苯二酚和間苯二酚的混合物,是兒茶酚的衍生物,可能通過簡單苯丙烷類化合物與長鏈脂肪酸反應得到[29]。擬南芥MYB家族蛋白中,AtMYB30、AtMYB60和AtMYB96在響應非生物脅迫中起著一定作用,且AtMYB30可通過調節長鏈脂肪酸的合成來響應病原體的侵襲[28],故同AtMYB30聚在同一亞組的漆樹MYB蛋白PB_c0_g6010、PB_c0_1517和PB_c0_g4337可能通過調節長鏈脂肪酸的合成進一步促進漆酚的形成。擬南芥MYB蛋白AtMYB11、AtMYB12和AtMYB111可調控類黃酮類物質的生物合成[30],故同這些蛋白處于同一分支的漆樹MYB蛋白PB_c0_g4405、PB_c0_g12918和PB_c0_g13021可能在黃酮類物質漆酚的生物合成中發揮重要調控作用。
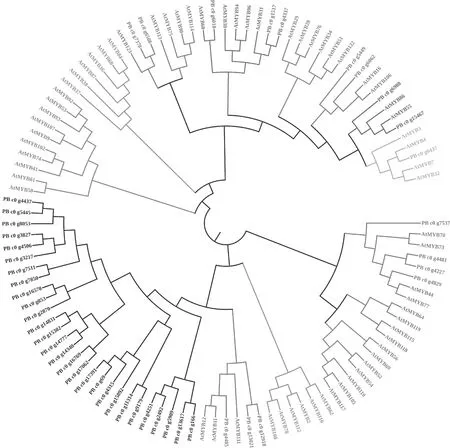
注:At代表擬南芥基因,PB代表漆樹基因。圖4 漆樹與擬南芥MYB家族蛋白聚類分析Fig.4 Cluster analysis of MYB protein from T.vernicifluum and Arabidopsis thaliana
2.7 漆樹MYB轉錄因子基因表達分析
基于漆樹轉錄組數據我們獲得了漆樹MYB轉錄因子在根、莖、葉各個組織中的基因表達量,以進一步了解漆樹MYB轉錄因子的生物學功能,應用ClustVis在線軟件對漆樹MYB轉錄因子在不同組織中的表達進行了聚類分析并繪制熱圖(圖5)。有11個MYB在葉片組織中的表達量較高,如PB_c0_g13130(28.27)、PB_c0_g6062(139.71)、PB_c0_g6437(64.15)、PB_c0_g6508(13.12)、PB_c0_g6908(51.43)、PB_c0_g7378(22.35)等;21個在莖中的表達量較高,其中表達量>10的有15個,PB_c0_g16578和PB_c0_g17062基因的表達量分別高達117.81和158.41,且PB_c0_g17062在3個組織中表達量都比較高,在莖中的表達量是所測基因中表達量最高的;16個MYB轉錄因子在根中表達上調,其中除PB_c0_g4481外各基因表達量均>10,而且基因PB_c0_g1517、PB_c0_g4337和PB_c0_g4437表達量分別高達95.53、151.03和93.88。以上不同MYB基因在不同組織中的差異表達情況表明MYB轉錄因子可能參與調控漆樹各個組織的發育。
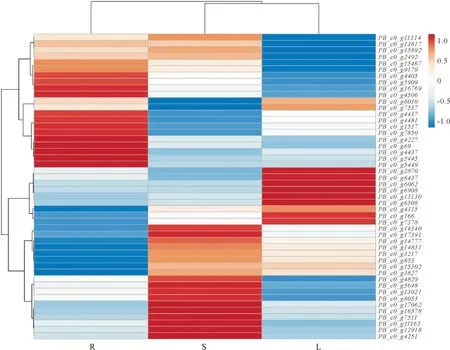
注:R.根;S.莖;L.葉。圖5 漆樹不同組織中MYB基因的表達譜Fig.5 Expression profiles of MYB genes between different tissues in T.vernicifluum
3 結論與討論
MYB轉錄因子是植物中最大的轉錄因子家族之一,在植物生長發育、次生代謝調控、脅迫應答等生命活動中起著調控作用[5]。擬南芥[31]、水稻[32]、玉米[33]、草莓[34]和桃[35]等植物中均已鑒定出MYB蛋白,且在基因組水平上對MYB家族基因進行了深入分析。
目前,轉錄組測序已成為研究植物形成與發育的分子機制的一種重要方法。姜福星等[36]應用高通量測序技術對瀘定百合的鱗莖進行了轉錄組分析,共獲得43 412條Unigene,為探究瀘定百合鱗莖形成和發育的分子機制、藥用成分以及百合的分子育種和研究開發提供了參考和依據。為培育品質優良的花椒無刺品種,篩選控制皮刺分化的關鍵基因,蔣弘剛等[37]基于二代高通量測序對花椒皮刺起始分化的莖尖節點組織進行轉錄組測序,最終獲得45 057條轉錄本序列,為尋找花椒皮刺分化的關鍵基因提供了豐富、可靠的基礎信息。本研究通過對漆樹轉錄組進行搜索,最終篩選鑒定出48個MYB轉錄因子,氨基酸數目為184~1 741,可能與可變剪切導致的不同基因編碼區和非編碼區在基因全長的比例呈現多態性有關[38]。對漆樹R2R3-MYB類轉錄因子的保守結構域分析表明其含有MYB家族典型的DNA-binding domain,包含102個左右的氨基酸殘基,與銀杏[1]R2R3結構域氨基酸殘基數目一致,而森林草莓的R2R3結構域包含107個左右的氨基酸殘基[34],大豆則包含104個左右的氨基酸殘基[5]。二級結構預測表明,無規則卷曲和α螺旋構成了漆樹MYB家族蛋白二級結構的主要成分。系統進化分析結果顯示,預測得到的漆樹48個MYB轉錄因子和擬南芥MYB轉錄因子被分為15個亞家族,漆樹28個MYB蛋白序列沒有與擬南芥的任何MYB亞家族聚到一起,這可能是漆樹進化過程中產生的調控其特有生物過程的轉錄因子。郭亞飛等[39]發現CsMYB123參與調控茶樹花青素的合成,該基因在各組織中均表達,且在新梢中高水平最高。田愛梅等[40]克隆了白芨MYB類轉錄因子PAP1基因并就該基因在白芨不同組織中的表達特征進行了分析,結果表明,基因BsPAP1在花中表達量較高,在塊根狀假鱗莖和嫩萌果中的表達較弱。李志根等[41]利用RACE技術從葡萄風信子花瓣中克隆了MYB轉錄因子基因MaMYB2并分析了其在不同組織部位中的表達模式,研究發現,MaMYB2可能參與葡萄風信子花青素積累的轉錄調控,且該基因在花中的表達高于根、莖和葉。月季‘紅勝利’花青素苷相關R2R3-MYB蛋白基因RhMYBs4-1和RhMYBs6-1在花瓣中高水平表達,在葉片和花藥中表達水平均較低[42]。對漆樹不同組織MYB的表達模式進行分析表明,漆樹MYB基因在各組織中均有表達且具有明顯的組織特異性,也有幾個基因在3種組織中表達水平相近,表明大部分漆樹MYB基因在特定組織中才會發揮作用,而有些基因在所有組織中均能發揮作用。
本研究在漆樹根、莖和葉轉錄組數據的基礎上,篩選鑒定得到48個MYB轉錄因子,對漆樹MYB轉錄因子家族的理化性質、蛋白功能結構域、系統進化關系以及不同組織的表達模式進行分析,初步鑒定了漆樹MYB轉錄因子家族。分析結果表明,漆樹MYB轉錄因子家族蛋白保守,R2R3-MYB類蛋白高度保守,且MYB轉錄因子在漆樹不同組織中表達模式各異。漆樹7 000萬a的生活史使其在生命進化過程中具有獨特的歷史地位,所產的生漆為自然界唯一的天然高分子涂料,環保,無污染。本研究為進一步探討漆樹MYB轉錄因子的生物學功能奠定了基礎。
