YAP和β-catenin在卵巢癌中的表達及臨床意義
2021-01-15 06:58:16李琳葉麗平
錦州醫科大學學報 2020年6期
關鍵詞:信號
李琳,葉麗平
(1.錦州醫科大學病理生理學教研室,遼寧 錦州 121000;2.錦州醫科大學附屬盤錦遼油寶石花醫院檢驗科,遼寧 盤錦 124010)
卵巢惡性腫瘤是婦科常見的惡性腫瘤之一,發病率僅次于子宮頸癌和子宮內膜癌而居第三位,但卻是導致女性癌癥死亡的首要原因。卵巢癌發病隱匿,具有異質性,目前仍缺乏有效的早期診斷和治療手段。研究表明,腫瘤的發生發展與細胞信號通路活性異常密切相關[1-2]。YAP(yes-associated protein,Yes相關蛋白),是Hippo信號通路的關鍵分子,參與調節細胞增殖、存活和分化,器官的發育、再生和干細胞等生物學功能,并與腫瘤的發生發展相關[3-4]。β-連環蛋白(β-catenin)是Wnt 信號通路的重要效應蛋白分子,在腫瘤中高表達并調節細胞的增殖、侵襲和凋亡等生命活動[5-6]。近年來越來越多的研究證實,Hippo/YAP通路與Wnt/β-catenin通路存在相互交叉作用,協同促進腫瘤的發生發展[7]。本研究應用免疫組化方法檢測了YAP和 β-catenin在卵巢癌中的表達情況,并分析了二者之間表達相關性及其各自與臨床病理因素的關系,旨在探討卵巢癌的發病機制,尋找有效的早診手段和分子靶向治療藥物。
1 材料和方法
1.1 組織來源
基于知情同意原則,收集盤錦遼油寶石花醫院住院卵巢癌手術切除的標本66例,所有病例在腫瘤切除前均未進行放、化療。根據年齡、病理類型、FIGO分期、分化程度、有無淋巴結轉移、遠處轉移進行分組。良性腫瘤組織標本30例,對照組30例取自同時期住院的卵巢切除患者。新鮮組織標本一部分凍存于-80 ℃冰箱備用,用于western blot,一部分經甲醛固定石蠟包埋,用于免疫組化染色。
1.2 免疫組織化學
30例正常卵巢組織、30例卵巢良性腫瘤、66例卵巢惡性組標本經10%中性甲醛固定,石蠟包埋,4 μm厚連續切片,常規脫蠟至水,3% H2O2室溫孵育5~10 min以滅活內源性過氧化物酶的活性。蒸餾水沖洗,PBS 浸泡 5 min×2次,抗原高壓熱修復,自然冷卻至室溫。 5%~10% 正常山羊血清封閉,室溫孵育 10 min,傾去血清,滴加一抗工作液,4 ℃孵育過夜。PBS 沖洗,5 min×3 次,滴加適當比例稀釋的生物素標記二抗,37 ℃ 孵育 10~30 min。PBS 沖洗,5 min×3次,DAB顯色,蘇木素復染,中性 樹膠封片,光鏡觀察。采用 0.01 M PBS液代替一抗作陰性對照。用已知YAP,β-catenin陽性的卵巢癌組織切片作為陽性對照。
請兩位病理科醫師雙盲閱片。每張切片隨機選取5個高倍視野,每個視野計數100個細胞,共計500個細胞。按陽性細胞數所占百分比評分:陰性為0分;陽性細胞數≤10%為1分;11%~50%為2分;51%~75%為3分;>75%為4分。再按染色強度評分:無色為0分;淡黃色為1分;棕黃色為2分;棕褐色為3分。按兩者乘積分數評分,0~2分為(-);3~4分為(+);5~8分為(++);9~12分為(+++)。
1.3 Western blotting 檢測YAP和β-catenin的蛋白表達
將盤錦遼油寶石花醫院婦科于2016年5月至2019年5月期間行卵巢手術切除的標本快速凍存,其中卵巢正常組織、良性腫瘤組織、卵巢癌組織各10例。加蛋白裂解液于放有剪碎組織試管內,超聲至勻漿狀態,在細胞蛋白提取完成后采用Bradford法進行測定。
采用10.0%的聚丙烯酰胺凝膠進行電泳以分離蛋白樣品,電轉移至PVDF膜,封閉后分別按1∶400和1∶1000的濃度加入YAP、β-catenin一抗和二抗,然后孵育2 h,雜交信號用ECL化學發光試劑盒進行檢測。X線曝光,顯影定影后掃描。各電泳帶的光密度值用Image Pro-plus圖像處理軟件分析,目的蛋白的相對表達水平通過計算目的片段與β-actin電泳帶密度的相對值來表示。
1.4 統計學分析
病理形態學資料采用SPSS 16.0統計軟件分析。χ2檢驗分析YAP,β-catenin表達的差異及其與病理臨床參數的關系。Pearson列聯系數分析YAP和β-catenin表達的相關性。P<0.05差異有統計學意義。
2 結 果
2.1 YAP和β-catenin在正常卵巢組織,良性卵巢腫瘤和卵巢癌中的表達情況
我們應用免疫組化的方法分別檢測了YAP和β-catenin在30例正常卵巢組織、30例良性卵巢組織和66例卵巢癌中的表達情況。我們發現YAP陽性表達主要定位細胞核,而β-catenin則主要定位于細胞膜。二者在惡性卵巢組織中的表達均明顯高于正常卵巢組織和良性卵巢腫瘤。YAP和β-catenin分別在30例正常卵巢組織中陽性表達率為10.0%(3/30)和16.7%(5/30),在30例良性卵巢組織中陽性表達率為33.3%(10/30)和40%(12/30),在66例卵巢癌中的陽性表達率為57.6%(38/66)和65.2%(43/66)。證明YAP和β-catenin均在卵巢癌中存在過表達,提示與卵巢的惡變有關,見圖1。
2.2 YAP、β-catenin的表達與卵巢癌患者臨床病理因素的關系
為了進一步明確YAP和β-catenin的臨床特點,我們分別統計分析了其表達與年齡,組織類型,FIGO分期,分化程度和淋巴結轉移等臨床病理因素的關系。結果發現 YAP和β-catenin均與FIGO分期有關,YAP在FIGOⅠ+Ⅱ期中陽性表達率為43.3%(13/30) 明顯低于Ⅲ+Ⅳ期中的陽性率69.4%(25/36),且有統計學差異(P<0.05)。同樣,β-catenin在FIGO Ⅲ+Ⅳ期中陽性表達率高于Ⅰ+Ⅱ期(86.1%(31/36)vs 40.0%(12/30),P<0.05)。此外,YAP在卵巢癌中過表達與淋巴結轉移有關(P<0.05),β-catenin在卵巢癌中過表達與分化程度有關(P<0.05)。二者的高表達均與年齡,組織類型無統計學差異,見表1、表2。
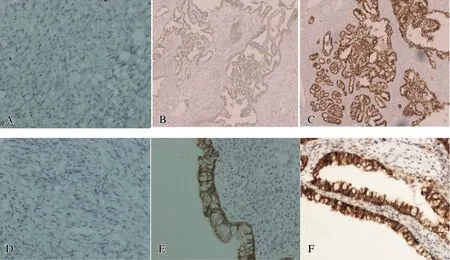
A:YAP在正常卵巢組織中的表達;B:YAP在良性卵巢腫瘤中的表達;C:YAP在卵巢癌中的表達;D:β-catenin在正常卵巢組織中的表達;E:β-catenin在良性卵巢腫瘤中的表達;F:β-catenin在卵巢癌中的表達
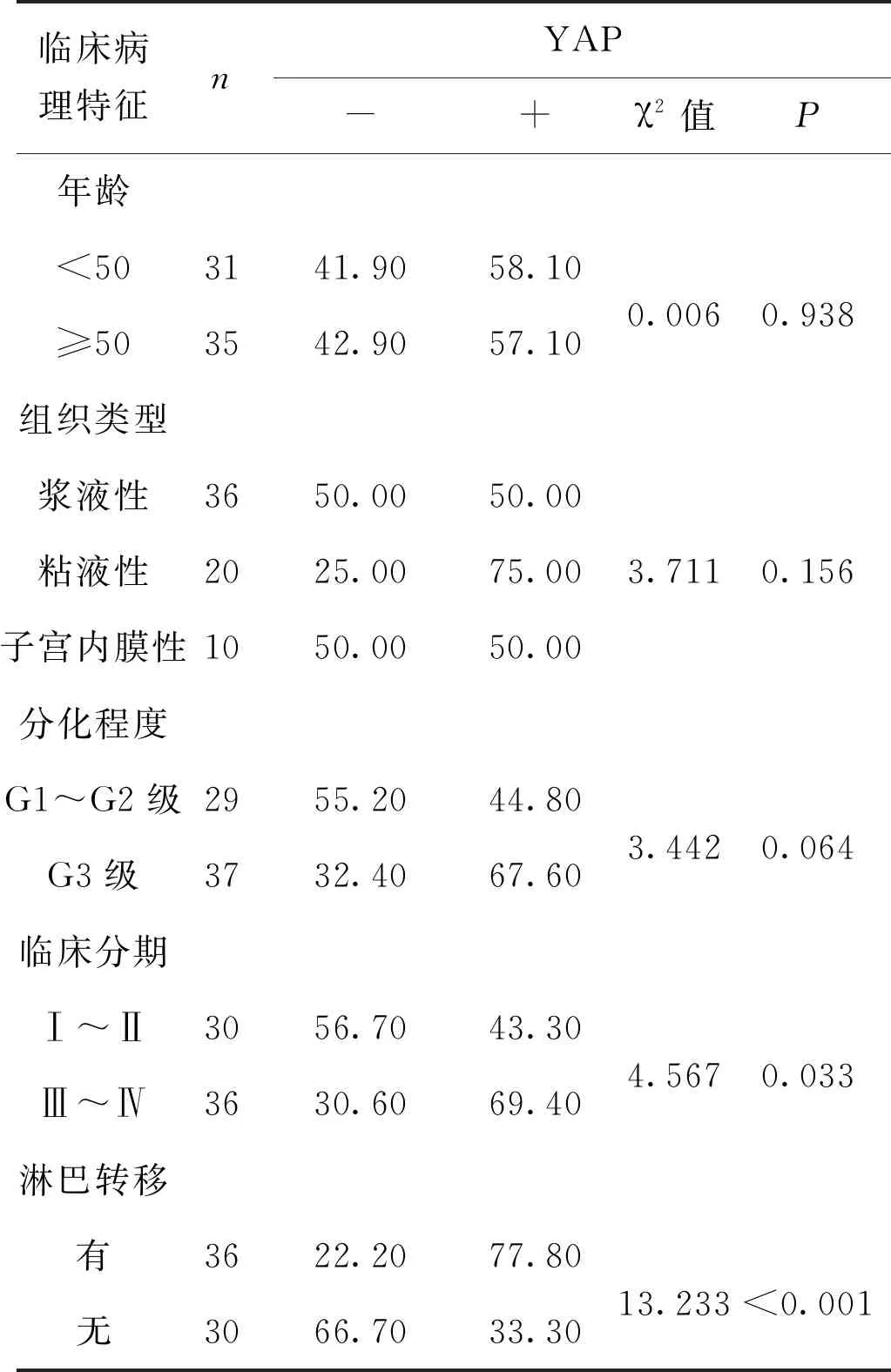
表1 YAP蛋白表達與卵巢癌臨床病理學參數關系(%)
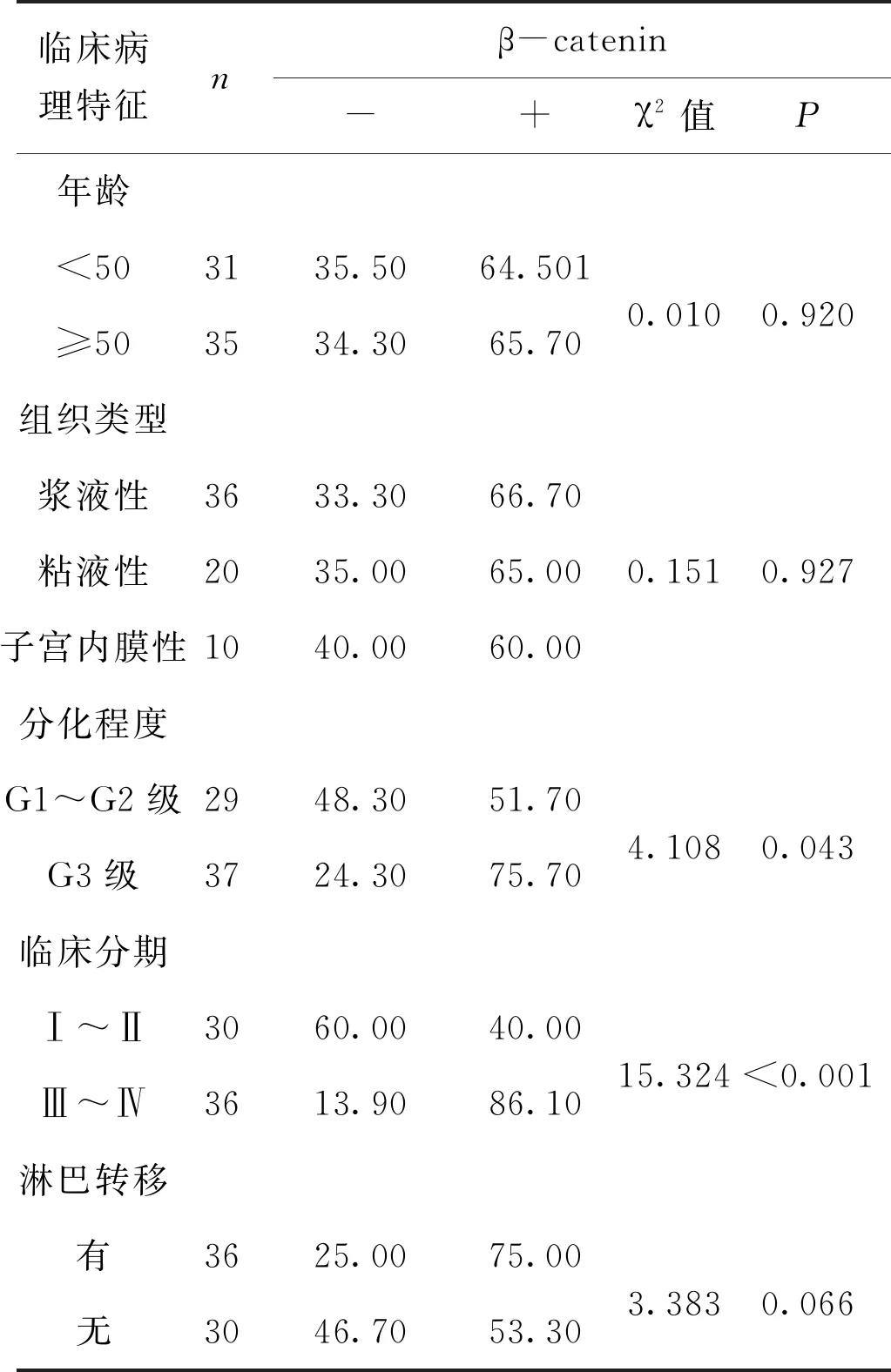
表2 β-catenin蛋白表達與卵巢癌臨床病理學參數關系(%)
2.3 卵巢癌中YAP和β-catenin的表達相關性
統計分析結果顯示66例卵巢癌患者中YAP 和β-catenin 表達均陽性者33 例,均陰性者18 例;YAP 表達陽性、β-catenin表達陰性者5例;YAP 表達陰性、β-catenin 表達陽性者10例,經過Pearson列聯系數分析:在66卵巢癌組織中,YAP 和β-catenin 的高表達呈顯著正相關(r=0.51,P<0.05),見表3。

表3 YAP、β-catenin二者表達的相關分析
2.4 Western blotting 檢測YAP、β-catenin的蛋白表達
YAP蛋白的分子質量為65 kDa,β-catenin的分子質量為92~95 kD,在正常卵巢組織、卵巢良性腫瘤組織和卵巢癌組織中均可見YAP、β-catenin的特異性條帶出現。
同正常卵巢組織組和卵巢良性腫瘤組相比,卵巢癌組織中的YAP和β-catenin的蛋白表達水平明顯更高,且差異有統計學意義(P<0.01),見表4、圖2。

表4 YAP、β-catenin在不同卵巢組織中的蛋白表達
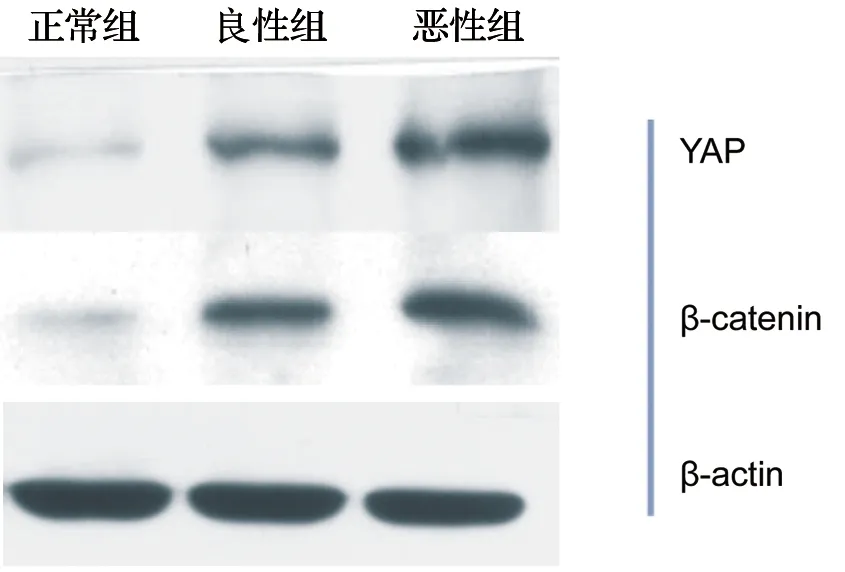
圖2 YAP、β-catenin在不同卵巢組織中的蛋白表達
3 討 論
近年來,卵巢癌發病率呈逐年上升趨勢,手術切除仍是治療的首選方式。然而,卵巢癌發病隱匿,患者就診時往往失去手術時機,目前仍未有有效的早診手段,導致卵巢癌仍是婦科腫瘤中死亡率最高的惡性腫瘤。伴隨基因組學的發展,從分子學探討腫瘤發病機制,為尋找有效的腫瘤診治靶點提供可能性。
腫瘤的發病機制是多因素,多階段,多基因之間的共同作用。研究證實在惡性腫瘤中存在多條信號通路的異常激活或失活。YAP定位于染色體11q22,該區域在人類多種腫瘤中被證實存在擴增或雜合型突變。Hippo信號通路的失活可使YAP在細胞漿積累,進入細胞核后與TEAD結合,從而調節目標基因的表達。研究表明,YAP在多種惡性腫瘤中表達異常,如結腸癌、肝癌、肺癌、前列腺癌、乳腺癌、口腔鱗癌、成神經管細胞瘤、食管鱗癌等,并調節細胞的惡性生物學行為[8]。β-catenin是Wnt信號傳導過程中的樞紐,當Wnt-1 信號與細胞膜上的受體蛋白Frizled(Ez)蛋白結合后,信號傳給dishevelled(DSH/Dvl),使其過磷酸化,DSH 的過磷酸化抑制了GSK-3β功能,使其不能磷酸化β-catenin 蛋白,導致胞漿內游離的β-catenin 增加,β-catenin進入胞核與轉錄因子LEF 和TCF形成復合物,促進如:c-myct 和cyclinD1 等下游靶基因的表達,從而影響細胞的增殖、分化、凋亡等行為[9]。已有研究證實Hippo/YAP通路與Wnt/β-catenin通路在肝癌、大腸癌、乳腺癌和膠質瘤中存在協同促進腫瘤的惡性進展的作用[7,10-11]71-79。但在卵巢癌中的作用研究尚未明確。
本研究證實YAP和β-catenin在卵巢癌中均存在過表達,提示與卵巢癌的發生和惡性進展有關。YAP在Hippo通路失活條件下,在細胞漿中聚集入核后發揮轉錄活化因子的作用。β-catenin在Wnt通路激活條件下,在細胞漿中聚集后入核亦發揮轉錄活化作用。二者均可以通過調節下游增殖、侵襲和凋亡相關因子的轉錄和表達而調控腫瘤細胞的惡性生物學行為。我們證實了YAP陽性表達主要定位于細胞核,而β-catenin主要定位于細胞膜。二者在卵巢癌中的陽性表達率均高于良性卵巢腫瘤和正常卵巢組織。在探討YAP和β-catenin在卵巢癌中高表達的臨床意義時,我們分析了二者過表達與卵巢癌臨床病理因素的關系即年齡、組織類型、FIGO分期、分化程度和淋巴轉移,結果發現二者均與FIGO分期有關(Ⅲ+Ⅳ期中的表達明顯高于Ⅰ+Ⅱ期)。其中YAP的過表達與卵巢癌淋巴結轉移有關,β-catenin的過表達與分化程度有關。
已有研究顯示,YAP和β-catenin定位于細胞核時能夠協同發揮轉錄活化的作用進而促進惡性腫瘤的發生發展。Hippo 信號通路活的激活能夠抑制Wnt信號通路,而Wnt信號通路的激活亦能抑制Hippo 信號通路的活性。在腸癌中Hippo 信號通路活性下調而Wnt信號通路活性上調,磷酸化YAP能夠促進β-catenin定位于細胞漿[12]。在膠質瘤中,YAP可以通過GSK3β調節β-catenin的表達進而影響腫瘤的增殖,β-catenin亦能調節YAP。在肝癌中[13],Wnt/β-catenin信號通路異常激活時,發現YAP亦起到關鍵促癌因子作用[14]。值得一提的是,我們在本研究中證實了YAP1 和β-catenin 在卵巢癌中高表達且二者表達呈正相關性,這與上述研究報道結果一致。提示我們YAP和β-catenin在卵巢癌中亦發揮協同促進癌癥惡性進展的作用,為進一步闡明Hippo 與Wnt 信號通路在卵巢癌的協同作用提供實驗基礎。但是,二者是如何發揮協同作用,仍需進一步研究證實。
綜上所述,YAP和β-catenin 分別作為Hippo和Wnt 信號通路中的關鍵分子在卵巢癌中高表達,并且均與臨床FIGO分期有關,重要的是二者表達呈正常相關。我們的研究證實了YAP和β-catenin在卵巢癌中的協同促癌作用,為探討卵巢癌發病分子機制提供理論研究基礎,為臨床卵巢癌早期診治提供依據。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
