基于線粒體標記的滁州鯽遺傳多樣性和遺傳結構
2021-01-13 08:23:00胡玉婷江河段國慶凌俊潘庭雙周華興汪煥
安徽農學通報 2021年24期
胡玉婷 江河 段國慶 凌俊 潘庭雙 周華興 汪煥
摘 要:為了解滁州鯽遺傳種質資源現狀,采用線粒體細胞色素b和控制區為分子標記,研究了滁州鯽保種群體和野生群體的遺傳多樣性和遺傳結構。研究獲得了長度分別為990bp的Cyt b部分序列和907bp的控制區全序列。2個群體相比較,保種群體遺傳多樣性略低于野生群體。總體而言,基于線粒體細胞色素b和控制區均顯示滁州鯽的遺傳多樣性偏低,但2個群體在2種分子標記中均不存在顯著的遺傳分化;不同序列間比較,2個群體的控制區不僅A+T含量更高,還具有更高的單倍型多樣性和核苷酸多樣性,表明控制區更適于研究滁州鯽的群體遺傳變異。
關鍵詞:滁州鯽;細胞色素b;控制區;遺傳多樣性
中圖分類號 S917.4 文獻標識碼 A 文章編號 1007-7731(2021)24-0012-04
Genetic Diversity and Genetic Structure of Chuzhou Crucian Carp based on Mitochondrial Markers
HU Yuting et al.
(Fishery Institute of Anhui Academy of Agricultural Sciences, Anhui Province Key Laboratory of Aquaculture & Stock Enhancement, Hefei 230031, China)
Abstract: In order to understand the genetic resources of Chuzhou crucian carp, mitochondrial cytochrome b and control region were used as molecular markers to study the genetic diversity and structure of the conserved and wild crucian carp populations. A partial sequence of 990 bp of Cyt b and a full sequence of 907 bp of control region were obtained.? The genetic diversity of the conserved population was slightly lower than that of the wild population, but there was no significant genetic differentiation between the two populations with two molecular markers. In general, the genetic diversity of Chuzhou crucian carp was low with two molecular markers.? The comparison between different sequences showed that the control region not only had higher A+T content, but also had higher haplotype diversity and nucleotide diversity, indicating that the control region was more suitable for studying population genetic variation of Chuzhou crucian carp.
Key words: Chuzhou crucian carp; Mitochondrial cytochrome b; Control region; Genetic diversity
滁州鯽(Carassius auratus),因產于安徽省滁州市滁河而得名,尤其盛產于滁州市西澗湖中[1],多年來該湖中鯽魚自然生息繁衍,最高年產量達10000kg,捕撈個體一般在500g以上,最大個體達2kg以上。滁州鯽以個體大,味道鮮美、色澤亮麗深受消費者青睞。早在明朝,《滁州志》就有記載:“滁州西澗鯽魚因其味美、功能表花,尤膾炙人口;肉肥厚細嫩,味甘香鮮醇、烏背金鱗,銀光閃閃,更名聞遐邇”。食者無不贊不絕口,因而價格也遠高于一般鯽魚。20世紀90年代以來,省內外學者開展了滁州鯽相關基礎研究,發現滁州鯽是生物進化過程中形成的地方性天然三倍體鯽魚種群,有雌核發育特性,系全國六大自然三倍體鯽魚品系之一,具有色澤亮麗、食性雜、生長快、病害少的特點[2-4],有較高的選育種價值和廣闊的推廣前景,2017年滁州鯽獲批為中華人民共和國地理標志產品,但滁州鯽資源保護不夠完善,品種選育還有很大潛力。
近年來,隨著滁州鯽養殖規模的不斷擴大,對滁州鯽苗種的需求量也越來越大,在實際生產過程中存在種質退化和品種混雜等問題,從而降低了其優良生產性能。而各種自然和人為因素導致其自然棲息地如西澗湖野生資源銳減,從而限制了滁州鯽繁育、養殖產業發展。目前滁州鯽的資源開發利用僅限當地極少數周邊地區。究其原因,有關滁州鯽的研究遠不夠充分,且主要集中在傳統的形態學、繁殖、養殖等基礎生物學方面[5-9],對其野生甚至保種群體的遺傳多樣性和遺傳結構現狀缺乏研究,限制了滁州鯽的資源保護與開發利用。本研究以滁州鯽保種和原產地西澗湖野生群體為對象,通過采用群體遺傳學廣泛使用的線粒體細胞色素b和控制區(D-loop)基因為分子標記,分析其遺傳多樣性和遺傳結構,為滁州鯽進一步的種質資源保護和開發利用提供科學依據。
1 材料與方法
1.1 材料 2021年6—8月,滁州鯽樣品分別采自滁州市農業科學研究院水產科研基地和西澗湖調查樣本,測量體重后,每個群體隨機取30尾樣品魚,剪取魚體右側背部肌肉保存于無水乙醇中備用。
1.2 方法
1.2.1 基因組的提取、PCR擴增與測序 采用動物基因組提取試劑盒(Universal Genomic DNA Extraction Kit),按照提示的操作步驟提取基因組DNA。Cyt b擴增和測序引物均為L14724和H15915[10],D-loop區擴增引物為CR1和CR2[11]。PCR擴增采用常規方法。擴增產物經1%的瓊脂糖凝膠電泳檢測后,委托生工生物工程(上海)股份有限公司純化測序。
1.2.2 序列與群體遺傳分析 使用軟件Seaview[12]和Bioedit[13]對相關序列進行比對、拼接,并經人工校正。采用軟件DNAsp 5.0[14]計算多態位點數目、單倍型數目、單倍型多樣性(haplotype diversity,Hd)和核苷酸多樣性(nucleotide diversity,Pi)。利用Mega 4.0軟件[15]計算序列堿基組成、序列變異率、轉換與顛換比值、群體內和群體間遺傳距離。由Arlequin 3.5軟件[16]計算群體分化指數(Fst)。群體間基因流(Nm)由公式Nm=(1/Fst-1)/2[17]計算得出。
2 結果與分析
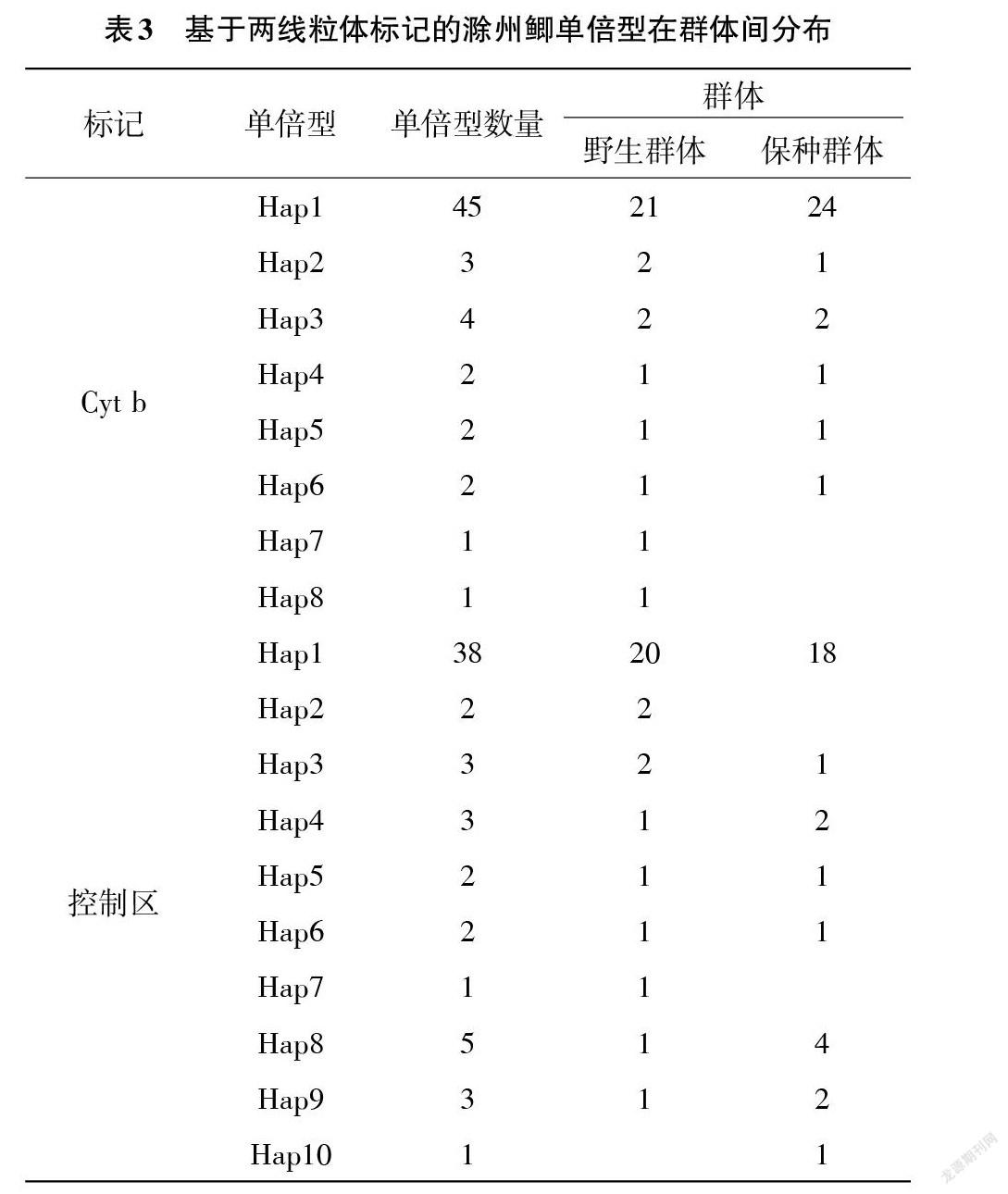
2.1 序列分析 經整理后得到長度一致為990 bp的線粒體Cyt b基因同源序列,包括從其起始端第16位至1015位,所有序列無插入和缺失。60條序列共檢測到6個多態位點,多態率較低(0.61%),包括簡約信息位點4個,單突變位點2個。線粒體Cyt b序列變異主要發生在密碼子第3位,僅少數發生在第1位和第2位,且全為同義突變,符合魚類線粒體中具有較慢的氨基酸變化速率的特征[18]。獲得所有樣本的線粒體控制區全序列長度一致為907bp,共檢測到8個多態位點(多態率僅0.88%),包括簡約信息位點7個,單突變位點1個。
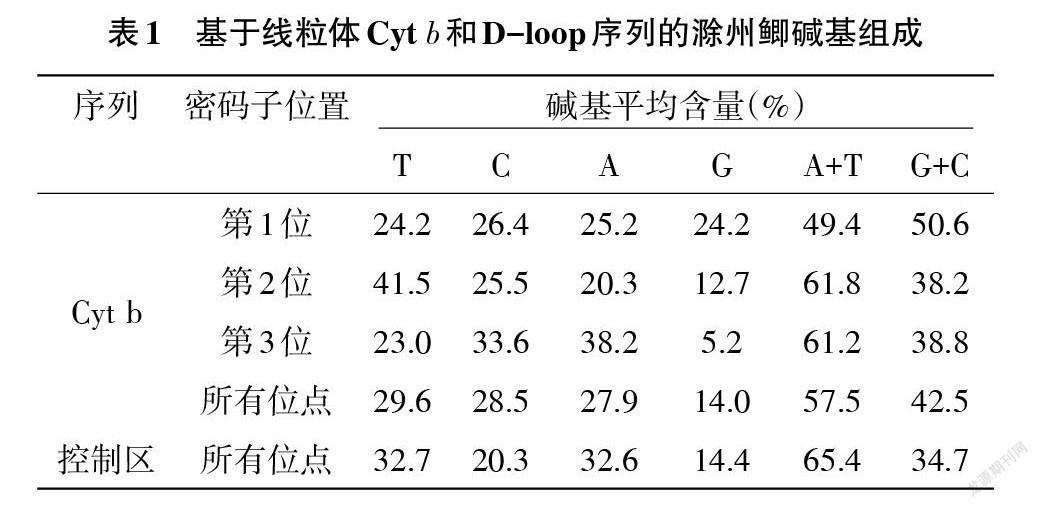
基于線粒體Cyt b和D-loop序列的滁州鯽2個群體的堿基組成見表1。相同序列的兩群體間堿基組成一致,具有明顯的AT偏好(A+T含量57.5%)和反G偏倚(G含量14.0%),符合脊椎動物線粒體基因堿基組成的普遍特征[19]。但Cyt b密碼子不同位次上堿基組成差異很大,從第1位的均衡到2、3位堿基G的含量越來越低。而與Cyt b序列比較,控制區具有更明顯的AT偏好,A+T含量高達65.4%。
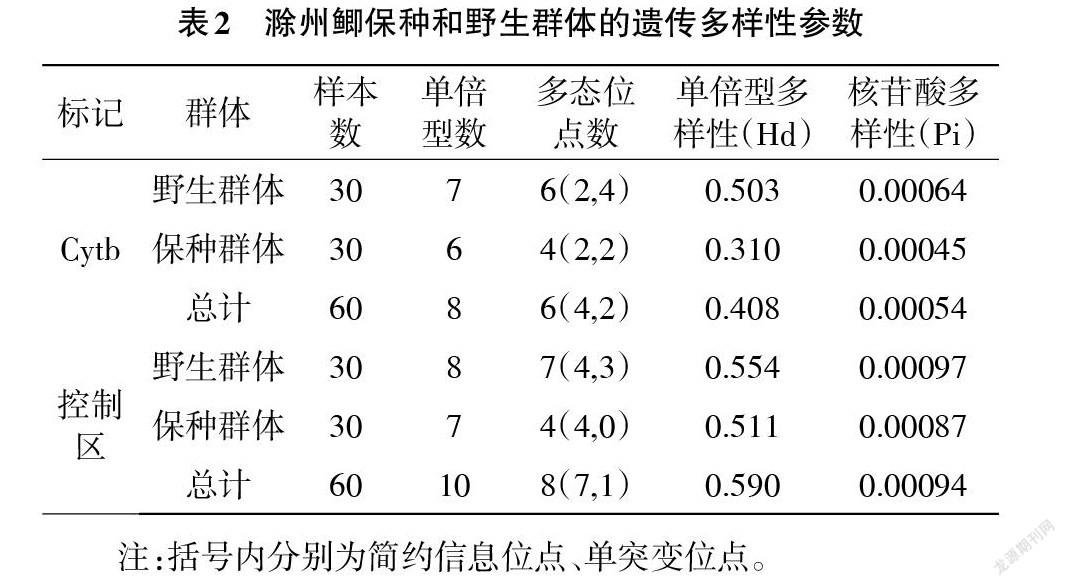
2.2 群體遺傳多樣性和遺傳結構 滁州鯽2個群體的遺傳多樣性參數見表2。除基于Cyt b的保種群體和總群體的單倍型多樣性較低,其他群體單倍型多樣性較高(>0.5),基于2個線粒體標記的所有群體核苷酸多樣性均較低(<0.01)。而所有群體的核苷酸多樣性均較低。2個群體間比較,基于2個標記的野生群體的單倍型多樣性和核苷酸多樣性均遠大于保種群體,顯示野生群體遺傳多樣性高于保種群體。不同標記間比較,基于D-loop區序列的所有遺傳多樣性參數均明顯大于Cyt b序列。
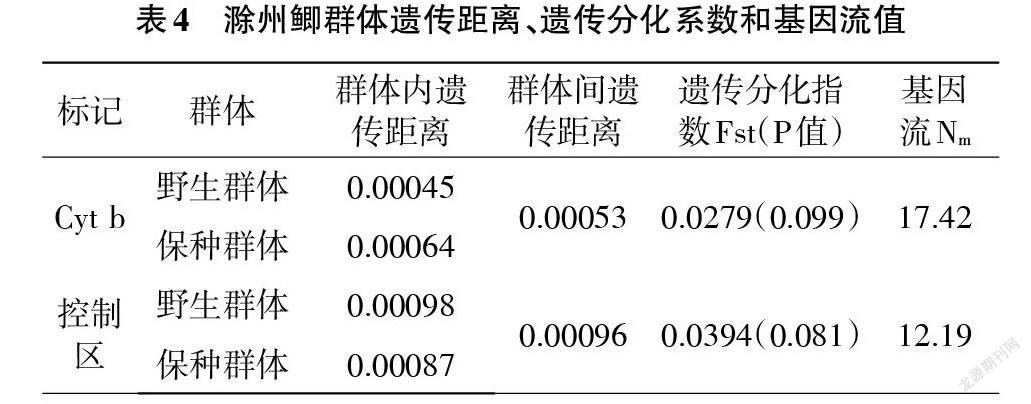
基于線粒體Cyt b和D-loop區的滁州鯽群體間遺傳距離均較小(<0.001),且與2個群體內的遺傳距離相差不大。基于2個標記的保種群體和野生群體間遺傳分化指數Fst均較小(<0.05),顯示群體間無顯著遺傳分化。
3 討論與結論
遺傳多樣性是生物多樣性的重要組成部分,是物種或群體長期進化的產物,也是其生存適應和發展進化的前提,其遺傳多樣性大小代表生物適應環境變化能力的強弱,既可揭示物種或群體過去的進化歷程,也能分析其進化潛力和未來命運。單倍型多樣性(Hd)和核苷酸多樣性(Pi)是衡量群體遺傳多樣性高低的重要參數,相比較單倍型多樣性,核苷酸多樣性更能體現遺傳多樣性的狀況。研究結果顯示:基于線粒體Cyt b和D-loop2種分子標記的滁州鯽保種群體、野生群體和總群體的單倍型遺傳多樣性(0.310~0.590)偏低或略高,而核苷酸多樣性(0.00045~0.00097)則全部偏低(<0.05)。這表明,滁州鯽的遺傳多樣性總體偏低,并低于其他地區的鯽魚群體,如低于基于線粒體COI序列的西江鯽8個群體的遺傳多樣性(Hd:0.714-0.917,Pi:0.0027-0.0087)[20]、基于線粒體控制區的都柳江鯽的遺傳多樣性(Hd:0.893,Pi:0.0063)[21]、基于線粒體Cyt b序列和D-loop區野生鯽魚的遺傳多樣性(Hd:0.933,Pi:0.00262、0.01127)[22]。滁州鯽遺傳多樣性較低的原因除了其天然分布范圍較窄、野生種群較小的緣故之外,主要是受到人類活動的影響:滁州鯽的主要棲息地西澗湖位于城區邊緣,有發電廠利用其水發電,導致西澗湖水位晝降夜升,而鯽魚把受精卵產于水草,但由于白天水位下降且氣溫升高使其難以孵化。
滁州鯽保種群體遺傳多樣性低于野生群體,可能與奠基者效應導致的種群遺傳多樣性水平較低有關。這種保種或養殖群體遺傳多樣性低于野生群體的現象在許多魚類研究中均有報道,如歐陽美等[23]基于線粒體Cyt b基因序列對長江中上游草魚野生和養殖群體遺傳多樣性進行了比較研究,結果顯示草魚群體多樣性水平大小依次是:野生群體>原種場群體>苗種場群體>放流群體。為提高保種群體的遺傳多樣性,可加大從野生群體采集更多單倍型個體,更新保種群體親本。
基于2種分子標記的滁州鯽單倍型間最大變異率很低(0.61%、0.88%),顯示為種內級別的差異。與線粒體Cyt b序列比較,兩群體的控制區不僅A+T含量更高,還具有更高的單倍型多樣性和核苷酸多樣性,表明控制區更適于研究滁州鯽的群體遺傳變異。群體遺傳結構上,當遺傳分化指數Fst在0至0.05 間為無分化,0.05至0.15 間為中度分化,0.15至0.25 間為高度分化[23]。基因交流程度是影響群體遺傳結構的重要因素,當群體間基因流Nm≥4時,基因流則成為主導因素,可防止群體間遺傳漂變[24]。本研究基于Cyt b序列和D-loop的滁州鯽兩群體間遺傳差異分析結果中Fst均小于0.05、Nm均遠大于4,表明2個群體間基因交流頻繁、遺傳差異較小,群體間無顯著遺傳分化。這可能與保種群體均取自野生群體并定期更新有關,保種群體繁育的后代放流到西澗湖,因而2個群體間存在定期的基因交流,遺傳差異不明顯。
綜上,滁州鯽群體包括保種群體和野生群體的遺傳多樣性較低,保種和野生群體間無顯著遺傳分化,但保種群體遺傳多樣性略低于野生群體,可通過從野生群體采集更多單倍型個體更新保種群體進而提高其遺傳多樣性水平,研究可為滁州鯽的資源保種和開發利用提供科學依據。
參考文獻
[1]張克儉,萬全,李公行,等.滁州鯽的形態學和血清蛋白電泳譜型[J].水產學報,1996,20(4):352?356.
[2]張克儉,萬全,李公行,等.滁州鯽染色體組型的研究[J].中國水產科學,1995,2(4):8?15.
[3]徐廣友,萬全,袁兆祥,等.滁州鯽雌核發育的細胞學研究[J].安徽農業大學學報,2009,36(4):533?537.
[4]尹永波,崔迎松,彭福,等.滁州鯽生物學特性及其苗種培育技術[J].中國水產,2005,(4):43.
[5]馬明發,張克儉,凌武海,等.三種群銀鯽形態特征的比較研究[J].河北漁業,2009(4):17?19.
[6]馬明發,袁兆祥,凌武海,等.滁州鯽生長特性的初步研究[J].水產養殖,2010,31(5):16?20.
[7]袁兆祥,凌武海,汪永忠,等.滁州鯽魚人工繁育與苗種培育技術研究[J].水產養殖,2010,31(3):2?4.
[8]凌武海,任信林.天然滁州鯽氨基酸成分分析與營養價值評價[J].水產養殖,2011,32(1):50?52.
[9]陳可東,萬全,蔡玉龍.滁州鯽夏花魚種適宜蛋白質需求量的研究[J].水利漁業,2004,24(2):23?24.
[10]Xiao W H,Zhang Y P,Liu H Z. Molecular systematics of Xenocyprinae (Teleostei:Cyprinidae):Taxonomy,biogeography,and coevolution of a special group restricted in East Asia[J]. Molecular Phylogenetics and Evolution,2001,18(2):163?173.
[11]劉良國,楊品紅,王文彬,等.洞庭青鯽與其他六個鯽魚群體線粒體DNA控制區的比較分析[J].水生生物學報,2010,34(2):378?38.
[12]Galtier N,Gouy M,Gautier C. SeaView and Phylo_win:two graphic tools for sequence alignment and molecular phylogeny[J]. Computer Applications in the Biosciences,1996,12:543?548.
[13]Hall,T.A. BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT [J]. Nucleic Acids Symp Ser,1999,41:95?98.
[14]Librado P,Rozas J. DnaSP v5:A software for comprehensive analysis of DNA polymorphism data [J]. Bioinformatics,2009,25:1451?1452.
[15]Kumar S,Dudley J,Nei M et al. MEGA:A biologist-centric software for evolutionary analysis of DNA and protein sequences [J]. Briefings in bioinformatics,2008,9(4):299?306.
[16]Excoffier L,Lischer HEL. Arlequin suite ver 3.5:A new series of programs to perform population genetics analyses under Linux and Windows [J]. Molecular Ecology Resources,2010,10:564?567.
[17]Hudson RR,Slatkint M,Maddison WP. Estimation of levels of gene flow from DNA sequence data [J]. Genetics,1992,(132):583?589.
[18]Kocher TD,Thomas WK,Meyer A. Dynamics of mitochondrial DNA evolution in animals:amplification and sequencing with conserved primers [J]. Proceedings of the National Academy of Sciences of the United States of America,1989,86:6196?6200.
[19]Cantatore P,Roberti M,Pesole G,et al. Evolutionary analysis of cytochrome b sequences in some perciformes:Evidence for a slower rate of evolution than in mammals[J]. Journal of Molecular and Evolution,1994,39(6):589?597.
[20]匡天旭,帥方敏,陳蔚濤,等.西江鯽的遺傳多樣性與群體結構[J].南方水產科學,2018,14(5):29?35.
[21]袁振興,代應貴,劉偉,等.都柳江鯉、鯽和草魚種群mtDNA控制區及遺傳多樣性分析[J].基因組學與應用生物學,2017,36(5):1926?1934.
[22]鄧朝陽.鯽4個群體線粒體Cytb序列和D-loop區的比較分析[J].現代農業科技,2015,(9):277?279,283.
[23]歐陽美,張曉宇,張富鐵,等.基于線粒體Cytb基因序列的長江中上游草魚野生和養殖群體遺傳多樣性比較研究[J].淡水漁業,2021,51(4):65?74.
[24]Wright S. Evolution and the genetics of populations. vol. 4. Variability within and among natural populations [A].University of Chicago Press, Chicago. 1978, 580.
(責編:王慧晴)
