朱砂葉螨幾丁質(zhì)脫乙酰基酶TecCDA3基因克隆及表達(dá)特征分析
2021-01-08 07:04:26宋莉紅卜春亞
北京農(nóng)學(xué)院學(xué)報(bào) 2021年1期
丁 超,宋莉紅,卜春亞
(北京農(nóng)學(xué)院生物與資源環(huán)境學(xué)院/農(nóng)業(yè)農(nóng)村部華北都市農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,北京 102206)
朱砂葉螨(Tetranychuscinnabarinus)別名紅蜘蛛,俗稱“紅蛐”,屬于蜱螨目葉螨科,為多食性害螨,主要寄主是瓜類、豆類等植物,廣泛分布于中國(guó)各地[1]。因其具有較強(qiáng)的繁殖能力且近親交配、生存領(lǐng)域小、世代周期短、對(duì)環(huán)境適應(yīng)性強(qiáng)、且時(shí)常接觸殺蟲(chóng)劑,導(dǎo)致其產(chǎn)生抗藥性比其他農(nóng)作物害蟲(chóng)更為嚴(yán)重[2]。目前全球市面上最主流的防治方法是依靠化學(xué)農(nóng)藥進(jìn)行化學(xué)防治,其具有見(jiàn)效快、成本低、面積廣等優(yōu)點(diǎn),常用的有炔螨特、阿維菌素、氟蟲(chóng)脲、噠螨靈、吡蟲(chóng)啉等[1];該方法有一定的缺點(diǎn),如易殘留、毒性大,對(duì)人體產(chǎn)生危害,噴灑的同時(shí)對(duì)周圍環(huán)境產(chǎn)生破壞。同時(shí)由于朱砂葉螨等害蟲(chóng)具有易產(chǎn)生抗藥性的特點(diǎn),極易造成后期難以殺死,需要加大農(nóng)藥的劑量,從而造成惡性循環(huán)。
幾丁質(zhì)脫乙酰基酶(CDA)是參與昆蟲(chóng)幾丁質(zhì)代謝的關(guān)鍵酶類,其主要作用是將幾丁質(zhì)脫乙酰基化形成殼聚糖,這種修飾可能有助于增強(qiáng)多種蛋白與幾丁質(zhì)特異性結(jié)合的親和力[3]。幾丁質(zhì)是昆蟲(chóng)生長(zhǎng)發(fā)育過(guò)程中不可或缺的多糖,對(duì)昆蟲(chóng)的外骨骼、表皮、圍食膜的形成起到重要作用。幾丁質(zhì)脫乙酰基酶在昆蟲(chóng)的蛻皮和圍食膜的形成過(guò)程中起到關(guān)鍵作用[4-6]。
自1984年初次在魯氏毛霉(Mucorroxianus)的提取物中發(fā)現(xiàn)幾丁質(zhì)脫乙酰基酶以來(lái),人們又陸續(xù)在昆蟲(chóng)、真菌、細(xì)菌中發(fā)現(xiàn)幾丁質(zhì)脫乙酰基酶的存在[7-10]。Guo等[11]在2005年首次在昆蟲(chóng)粉紋夜蛾的中腸中發(fā)現(xiàn)并鑒定其幾丁質(zhì)脫乙酰基酶,發(fā)現(xiàn)其在沒(méi)有幾丁質(zhì)結(jié)合區(qū)域的情況下有很強(qiáng)的幾丁質(zhì)結(jié)合活性并與圍食膜的特性相關(guān)。后來(lái),Arakane等[12]在赤擬谷盜中發(fā)現(xiàn)多種幾丁質(zhì)脫乙酰基酶,通過(guò)對(duì)其性質(zhì)和結(jié)構(gòu)劃分出詳細(xì)的幾丁質(zhì)脫乙酰基酶家族(GroupⅠ-GroupⅤ),GroupⅠ、GroupⅡ有3個(gè)保守區(qū),GroupⅢ、GroupⅣ有幾丁質(zhì)結(jié)合結(jié)構(gòu)域和乙酰基催化結(jié)構(gòu)域,GroupⅤ只具有乙酰基催化結(jié)構(gòu)域[13]。并通過(guò)RNAi的手段驗(yàn)證部分幾丁質(zhì)脫乙酰基酶對(duì)昆蟲(chóng)生長(zhǎng)發(fā)育的影響。
目前研究表明哺乳動(dòng)物和高等植物中并沒(méi)有幾丁質(zhì)脫乙酰基酶,故而可以利用幾丁質(zhì)代謝相關(guān)生物防治藥物去調(diào)控昆蟲(chóng)的生長(zhǎng)發(fā)育和繁殖,從而達(dá)到防治害螨的目的,且對(duì)環(huán)境和人類的副作用小,符合綠色農(nóng)業(yè)的理念。該研究旨在克隆朱砂葉螨TecCDA3基因并分析其表達(dá)特征,為闡明朱砂葉螨幾丁質(zhì)脫乙酰基酶家族奠定堅(jiān)實(shí)基礎(chǔ)。
1 材料與方法
1.1 供試用昆蟲(chóng)與試劑
供試用昆蟲(chóng)為北京農(nóng)學(xué)院農(nóng)業(yè)農(nóng)村部華北都市農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室培養(yǎng)的敏感品系朱砂葉螨,通過(guò)人工種植的白花蕓豆進(jìn)行飼養(yǎng),未經(jīng)任何特殊處理,溫度26 ℃±1 ℃,相對(duì)濕度50%±10%,光照/黑暗為16 h/8 h,此為朱砂葉螨的最適生長(zhǎng)環(huán)境。EasyPure?RNA Kit、TransScript?One-Step gDNA Removal and cDNA Synthesis SuperMix購(gòu)自北京全式金生物技術(shù)有限公司;X5 High-Fidelity DNA Polymerase PCR Mix、M5 HiPer pTOPO-Blunt Cloning Kit、M5 HiPer Top10 Competent Cell購(gòu)自北京聚合美生物科技有限公司;FastPure Gel DNA Extraction Mini Kit、FastPure Plasmid Mini Kit購(gòu)自南京諾唯贊生物科技有限公司;TB Green?Premix Ex TaqTM Ⅱ(Tli RNaseH Plus)購(gòu)自大連寶生物(Takara)公司。
1.2 朱砂葉螨幾丁質(zhì)脫乙酰基酶TecCDA3基因克隆
1.2.1 朱砂葉螨的培養(yǎng) 朱砂葉螨有4個(gè)生長(zhǎng)時(shí)期,分別為卵時(shí)期、幼螨時(shí)期、若螨時(shí)期、成螨時(shí)期。用細(xì)毛刷挑取約1 000頭生長(zhǎng)狀況良好的朱砂葉螨雌成螨到一盆新白蕓豆葉上,將其放回養(yǎng)螨室進(jìn)行為期24 h的產(chǎn)卵,使用體式顯微鏡觀察產(chǎn)卵數(shù)量,若產(chǎn)卵數(shù)量較多且密集則將葉片上的雌成螨盡數(shù)挑下,使之只剩卵于其葉上,繼續(xù)觀察,約3 d朱砂葉螨由卵發(fā)育至幼螨,約5 d發(fā)育至若螨,約9~10 d發(fā)育至成螨。不同時(shí)期的螨均通過(guò)自然發(fā)育得到。將同一時(shí)間段所有螨均用細(xì)毛刷挑至0.02%的吐溫20溶液中進(jìn)行細(xì)分,卵與幼螨使用0.175 mm篩子篩得幼螨,幼螨與若螨使用0.225 mm篩子篩得若螨,若螨與成螨使用0.3 mm篩子篩得成螨。過(guò)篩后用清水輕輕沖凈螨身上的吐溫溶液,將篩子倒扣于干燥的干凈吸水紙上吸干水分,通風(fēng)干燥約2 min,待螨可以活動(dòng)后用細(xì)毛刷收集最少10 mg同一時(shí)期的螨至1.5 mL離心管中,4個(gè)時(shí)期共計(jì)4個(gè)離心管。
1.2.2 朱砂葉螨總RNA提取 將1.2.1中的4個(gè)離心管快速置于液氮中,急速降溫約30 s,用電動(dòng)研磨器將組織研磨成粉末,參照EasyPure?RNA Kit說(shuō)明書(shū)對(duì)朱砂葉螨進(jìn)行總RNA提取。
1.2.3 第一鏈cDNA合成 參照TransScript?One-Step gDNA Removal and cDNA Synthesis SuperMix說(shuō)明書(shū)對(duì)朱砂葉螨總RNA進(jìn)行反轉(zhuǎn)錄生成cDNA。
1.2.4 引物設(shè)計(jì) 根據(jù)前期獲得的朱砂葉螨轉(zhuǎn)錄組數(shù)據(jù),通過(guò)NCBI的Blast找到高同源性物種二斑葉螨,以其幾丁質(zhì)脫乙酰基酶基因?yàn)閰⒄眨O(shè)計(jì)5′引物和3′引物進(jìn)行同源克隆,TecCDA3-F為5′-AATCACCACTGAATTTAGCCAAT-3′,TecCDA3-R為5′-TTGGTCAAAAGTGAGATTACTTG-3′。
1.2.5 基因擴(kuò)增 以朱砂葉螨cDNA為模板進(jìn)行PCR擴(kuò)增(50 μL),體系參照X5 High-Fidelity DNA Polymerase PCR Mix說(shuō)明書(shū)。PCR擴(kuò)增程序?yàn)轭A(yù)變性95 ℃,3 min;變性95 ℃,10 s;退火54 ℃,15 s;延伸72 ℃,1 min;34個(gè)循環(huán);大延伸72 ℃,5 min。用1%瓊脂糖凝膠電泳對(duì)PCR產(chǎn)物進(jìn)行檢驗(yàn),對(duì)符合大小的產(chǎn)物進(jìn)行純化,純化后連接至TOPO載體上,轉(zhuǎn)化至Top10感受態(tài)細(xì)胞中,菌液PCR進(jìn)行驗(yàn)證,對(duì)陽(yáng)性克隆進(jìn)行質(zhì)粒提取并送交上海生工生物公司進(jìn)行測(cè)序。
1.3 朱砂葉螨幾丁質(zhì)脫乙酰基酶TecCDA3基因特征分析
通過(guò)NCBI的ORF Finder(http://www.ncbi.nlm.nih.gov/home/analyze)進(jìn)行基因開(kāi)放閱讀框的推導(dǎo);將朱砂葉螨CDA的CDS區(qū)域翻譯成氨基酸序列后,通過(guò)NCBI的Blast工具(https://blast.ncbi.nlm.nih.gov/Blast.cgi)軟件進(jìn)行氨基酸序列同源性比對(duì),使用MEGA7.0系統(tǒng)發(fā)育樹(shù)的構(gòu)建,使用ML法進(jìn)行發(fā)育樹(shù)構(gòu)建,使用Bootstrap=200的方法進(jìn)行結(jié)果數(shù)據(jù)的計(jì)算;通過(guò)SMART(https://smart.embl-heidelberg.de)進(jìn)行蛋白功能域的預(yù)測(cè);使用ProtScale(http://web.expasy.org/protscale)進(jìn)行蛋白疏水區(qū)的分析。
1.4 朱砂葉螨幾丁質(zhì)脫乙酰基酶TecCDA3在不同生長(zhǎng)時(shí)期的表達(dá)規(guī)律
1.4.1 不同生長(zhǎng)時(shí)期朱砂葉螨的RNA提取和cDNA合成 使用全式金EasyPure?RNA Kit試劑盒分別對(duì)各個(gè)時(shí)期的朱砂葉螨進(jìn)行RNA提取,然后通過(guò)全式金TransScript?One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒以提取的RNA為模板反轉(zhuǎn)錄合成cDNA。
1.4.2 實(shí)時(shí)熒光定量PCR 用Beacon Designer 7軟件設(shè)計(jì)TecCDA3基因的RT-qPCR引物,同時(shí)設(shè)計(jì)一對(duì)β-actin基因?yàn)閮?nèi)參,引物見(jiàn)表1。
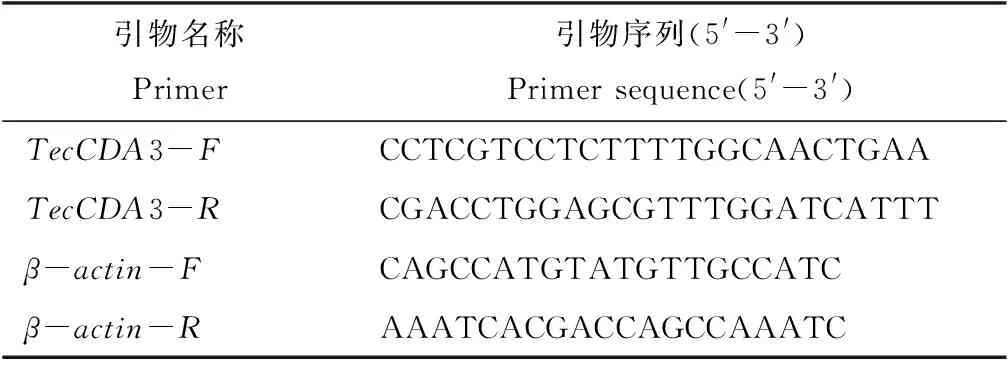
表1 TecCDA3熒光定量引物設(shè)計(jì)Tab.1 Design of RT-qPCR primers for TecCDA3
分別以朱砂葉螨4個(gè)時(shí)期的cDNA為模板,參照Takara公司的TB Green?Premix Ex TaqTM Ⅱ(Tli RNaseH Plus)說(shuō)明書(shū)對(duì)熒光定量體系進(jìn)行配置,PCR反應(yīng)條件為二步法PCR。95 ℃預(yù)變性30 s,95 ℃變性5 s,60 ℃退火延伸30 s,共40個(gè)循環(huán)。單次試驗(yàn)中每個(gè)樣本設(shè)置3次重復(fù),同時(shí)設(shè)置陰性對(duì)照,陰性對(duì)照重復(fù)2次。程序設(shè)置使用相對(duì)定量△△Ct法,CT值使用2-△△Ct法計(jì)算其相對(duì)表達(dá)量,設(shè)定EGG時(shí)期為參照,相對(duì)表達(dá)量為1,使用SPSS軟件對(duì)數(shù)據(jù)進(jìn)行顯著差異性分析,采用單因素方差分析(ANOVA)并采用Tukey's HSD算法進(jìn)行分析(P<0.05)。
2 結(jié)果與分析
2.1 朱砂葉螨幾丁質(zhì)脫乙酰基酶TecCDA3基因克隆
以朱砂葉螨cDNA為模板擴(kuò)增得到與預(yù)期大小相一致的基因,大小約為1.6 kb。通過(guò)NCBI的ORF Finder工具分析TecCDA3的開(kāi)放閱讀框,見(jiàn)圖1。TecCDA3包含一個(gè)長(zhǎng)度為1 611 bp的開(kāi)放閱讀框(Open Reading Frame,ORF),共編碼536個(gè)氨基酸,以5′端的起始密碼子ATG開(kāi)始編碼蛋白,TAA為其終止密碼子,蛋白分子量為62.07 kD。命名為TecCDA3(Genbank:MK779705)。
2.2 TecCDA3氨基酸序列同源比對(duì)分析
使用NCBI的Blast工具進(jìn)行篩選,得到多組高同源性物種的氨基酸序列,用MEGA7.0軟件對(duì)二斑葉螨(Tetranychusurticae)、赤擬谷盜(Pseudostellacrassipes)、黑腹果蠅(Drosophilamelanogaster)的CDA3氨基酸序列與朱砂葉螨TecCDA3進(jìn)行同源比較,得到圖2。比對(duì)結(jié)果顯示朱砂葉螨幾丁質(zhì)脫乙酰基酶與二斑葉螨具有最高的同源性;使用SMART在線工具對(duì)朱砂葉螨TecCDA3的氨基酸序列進(jìn)行蛋白結(jié)構(gòu)域分析,TecCDA3含有幾丁質(zhì)結(jié)合結(jié)構(gòu)域(ChBD),低密度脂蛋白受體結(jié)合結(jié)構(gòu)域(LDLa)和多糖多乙酰基催化結(jié)構(gòu)域(Polysaccharide deacetylase)。Group Ⅱ型幾丁質(zhì)脫乙酰基酶均含有這3種催化結(jié)構(gòu)域,并且呈現(xiàn)高度的保守性。TecCDA3蛋白從第12個(gè)氨基酸到第31個(gè)氨基酸之間有明顯跨膜結(jié)構(gòu),共計(jì)20個(gè)氨基酸,表明其為跨膜蛋白,但是其并不含有信號(hào)肽位點(diǎn)。
2.3 TecCDA3的疏水性分析
通過(guò)ProtScale在線軟件在線對(duì)朱砂葉螨幾丁質(zhì)脫乙酰基酶的疏水性進(jìn)行分析預(yù)測(cè),結(jié)果如圖3。當(dāng)Score>0的得分時(shí)表示該區(qū)域?yàn)槭杷裕?dāng)Score<0的得分時(shí)表示該區(qū)域?yàn)橛H水性,得分絕對(duì)值越高表明相對(duì)的親疏水性越高。TecCDA3的氨基酸序列中均含有親水區(qū)域和疏水區(qū)域,且分布總體上來(lái)說(shuō)較為均勻,但是可以明顯看出處于Score<0的得分區(qū)域要多于Score>0的得分區(qū)域,故而總體上來(lái)說(shuō)屬于親水蛋白,計(jì)算平均疏水性為-0.403,與預(yù)測(cè)的結(jié)果一致,為親水性蛋白。
2.4 分子進(jìn)化樹(shù)構(gòu)建
發(fā)育樹(shù)主要選取包括朱砂葉螨在內(nèi)的多種物種的幾丁質(zhì)脫乙酰基酶,如圖4。通過(guò)發(fā)育樹(shù)的構(gòu)建可以看出,TecCDA3與同屬蜱螨目的二斑葉螨的幾丁質(zhì)脫乙酰基酶親緣性最高,由于高度的序列相似性,其遺傳距離十分相近。命名為TecCDA3,除此之外可以看出朱砂葉螨的幾丁質(zhì)脫乙酰基酶與蜱螨目的物種親緣關(guān)系最近,如柑橘全爪螨(Panonychuscitri)、二斑葉螨(Tetranychusurticae),它們處于同一條分支上;與黑腹果蠅(Drosophilamelanogaster)、赤擬谷盜(Triboliumcastaneum)等相對(duì)遺傳距離較近;與家蠶(Bombyxmori)、中華稻蝗(Oxyachinensis)等相對(duì)遺傳距離較遠(yuǎn)。通過(guò)構(gòu)建的發(fā)育樹(shù)可以明顯看出幾丁質(zhì)脫乙酰基酶基因家族的分布,為GroupⅠ至GroupⅤ,其中TecCDA3屬于Group Ⅱ。
2.5 TecCDA3基因在朱砂葉螨不同生長(zhǎng)時(shí)期的表達(dá)規(guī)律
將得到的數(shù)據(jù)通過(guò)2-△△Ct法計(jì)算朱砂葉螨在4個(gè)生長(zhǎng)時(shí)期相對(duì)表達(dá)量,結(jié)果通過(guò)Origin軟件進(jìn)行作圖,見(jiàn)圖5。朱砂葉螨幾丁質(zhì)脫乙酰基酶在4個(gè)生長(zhǎng)時(shí)期(卵時(shí)期、幼螨時(shí)期、若螨時(shí)期、成螨時(shí)期)的mRNA豐度的相對(duì)差異,該試驗(yàn)使用的內(nèi)參基因?yàn)棣?actin,其作為看家基因在細(xì)胞中的表達(dá)量較為穩(wěn)定。TecCDA3在朱砂葉螨各個(gè)生長(zhǎng)時(shí)期均有表達(dá),但是表達(dá)量之間具有顯著差異,TecCDA3在成螨時(shí)期表達(dá)量達(dá)到最高,在幼螨時(shí)期表達(dá)量最低,在卵時(shí)期的表達(dá)量要高于若螨時(shí)期,整體呈現(xiàn)出一種先下降后上升的表達(dá)趨勢(shì)。
3 討 論
目前在中華稻蝗(O.sinensis)、赤擬谷盜(T.castaneum)、意大利蜂(I.bee)、黑腹果蠅(D.melanogaster)、岡比亞瘧蚊(A.gambiae)、柑橘全爪螨(C.citris)、家蠶(B.mori)、云山卷葉蛾(S.budworm)、褐飛虱(Nilaparvatalugens)、柑橘木虱(D.citri)等多種昆蟲(chóng)中均發(fā)現(xiàn)了幾丁質(zhì)脫乙酰基酶。其中赤擬谷盜最高的含有9種亞型基因,但并不是所有昆蟲(chóng)都具備全部的9種亞型基因。這9種亞型被分為5個(gè)家族,這5個(gè)家族中目前研究最多的是Group Ⅰ,該家族基因被證實(shí)具有參與昆蟲(chóng)體內(nèi)幾丁質(zhì)代謝的作用,影響昆蟲(chóng)胚胎表皮的形成和氣管幾丁質(zhì)的排列[14]。對(duì)其余的4個(gè)家族目前研究較少。
幾丁質(zhì)脫乙酰基酶TecCDA3為親水性蛋白,具有一個(gè)跨膜結(jié)構(gòu),無(wú)信號(hào)肽。由于不具備信號(hào)肽可能造成無(wú)法引導(dǎo)蛋白到達(dá)細(xì)胞含不同膜結(jié)構(gòu)的亞細(xì)胞器內(nèi),導(dǎo)致其無(wú)法加工。發(fā)現(xiàn)其氨基酸序列中含有3種不同的結(jié)構(gòu)域,分別為幾丁質(zhì)結(jié)合結(jié)構(gòu)域(ChBD)、多糖多乙酰基催化結(jié)構(gòu)域(Polysaccharide deacetylase)和低密度脂蛋白受體結(jié)合結(jié)構(gòu)域(LDLa),且具有高度的保守性。分子進(jìn)化樹(shù)分析可知,朱砂葉螨與二斑葉螨的親緣性最高。根據(jù)幾丁質(zhì)脫乙酰基酶具備的3個(gè)保守區(qū)域數(shù)目以及同源比對(duì)結(jié)果將TecCDA3歸類為Group Ⅱ家族。目前的研究表明赤擬谷盜TcCDA3在成蟲(chóng)的胸肌中特異性表達(dá)[13]。NlCDA3為褐飛虱的腸道特異性幾丁質(zhì)脫乙酰基酶,在褐飛虱的各個(gè)生長(zhǎng)發(fā)育階段均有較高的表達(dá)[15]。柑橘木虱DcCDA3在其外表皮和3日齡若蟲(chóng)中含量最高,且表達(dá)水平與20-hydroxyecdysone的催化有關(guān)[16]。該研究的TecCDA3與北京農(nóng)學(xué)院農(nóng)業(yè)農(nóng)村部華北都市農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室先前研究的與控制蛻皮有關(guān)的TecCDA1和TecCDA2之間的表達(dá)量存在著顯著差異,TecCDA1和TecCDA2均在卵時(shí)期有最高或較高的表達(dá)量,而TecCDA3則在卵時(shí)期相對(duì)表達(dá)量較低,在成螨時(shí)期表達(dá)量最高,這可能暗示其功能上的差異,其對(duì)朱砂葉螨是否具有影響蛻皮功能還有待進(jìn)一步探究。該試驗(yàn)結(jié)果豐富朱砂葉螨幾丁質(zhì)脫乙酰基酶基因家族,為基于靶向殺蟲(chóng)基因的篩選提供理論和實(shí)踐依據(jù)。
