灘羊骨骼肌衛(wèi)星細胞體外培養(yǎng)及成肌和成脂誘導分化研究
2021-01-07 14:30:02徐小春陳文娟馬文平
家畜生態(tài)學報 2020年12期
徐小春,趙 瑞,陳文娟,馬文平,馬 森
(1.北方民族大學 生物科學與工程學院,寧夏 銀川 750021;2.河南農業(yè)大學 牧醫(yī)工程學院,河南 鄭州 450002)
骨骼肌衛(wèi)星細胞(Skelatal muscle satellite cells,SMSCs)是位于骨骼肌纖維基底板下的一小群肌原性干細胞[1],因直接參與骨骼肌分化而受到廣泛關注[2-4]。在肌發(fā)生過程中,刺激成肌細胞啟動表達肌源性分化特異性基因,如中間絲蛋白(desmin)[5]、配對盒基因7(Pax7)[5-6]、肌肉細胞生成素(MyoG)[7]、肌球蛋白重鏈(MHC)[8]。SMSCs活化后表達肌特異性轉錄因子Pax7和MyoD[9],分化后表達肌原蛋白desmin,參與運動,并可用于識別其他組織中的骨骼肌細胞[10]。因此,MyoG和MHC是骨骼肌發(fā)育的調控因子,可促進SMSCs向成熟肌細胞分化,并作為SMSCs分化的標志物。SMSCs是參與生長后肌肉再生的最重要的細胞類型,通過對SMSCs成肌分化的調控可以有效改善畜禽的肉品質。骨骼肌衛(wèi)星細胞是一種多功能干細胞,既可以向成肌細胞分化,也可以向脂肪細胞分化[11-12]。研究證明,過氧化物酶體增殖物激活受體γ(PPARγ)、CCAAT/增強子結合蛋白α(C/EBPα)是調節(jié)脂肪形成的主要轉錄因子,并且不同程度地調控脂肪的發(fā)育。脂肪酸合成酶(FAS)在調控脂肪酸合成及脂肪細胞分化過程中起著重要作用,因此常被作為檢測脂肪細胞分化的標志基因[13-14]。目前灘羊優(yōu)良肉質性狀的體內研究形成機制尚不清楚,因此獲取便捷的體外研究材料對深入揭示其優(yōu)良肉質性狀形成分子機制具有重要意義,而SMSCs正是一種理想的研究材料。本研究通過對灘羊SMSCs進行分離培養(yǎng)和鑒定,同時建立SMSCs成肌和成脂誘導分化體系的,以期為在體外條件下研究調控灘羊肌肉生長發(fā)育及脂肪沉積的分子機制提供模型。
1 材料與方法
1.1 試驗動物與主要試劑
選取10日齡健康灘羊公羔1只(寧夏鹽池縣春浩林草產業(yè)專業(yè)合作社提供),頸部處死后無菌條件下采取背最長肌組織,放于加雙抗的(100 IU/mL青霉素和100 μg/mL鏈霉素)無菌磷酸緩沖液(phosphate buffer saline,PBS)中備用。0.1%I型膠原酶(Gibco,美國)、胎牛血清(fetal bovine serum,F(xiàn)BS)(Gibco,美國)、馬血清(Hyclone,美國)、DMEM/F12培養(yǎng)基(Sigma,美國)、0.25%胰蛋白酶(Gibco,美國)、青鏈霉素(Gibco,美國)、OriCell SD大鼠脂肪間質干細胞成脂誘導分化培養(yǎng)基試劑盒(Cyagen Biosciences,中國)、油紅O染料(Sigma,美國)、二甲基亞砜(dimethyl sulfoxide,DMSO;Sigma, 美國)、RNA提取試劑盒(Takara,日本)、反轉錄試劑盒(Takara,日本)、實時熒光定量qPCR Mix(Takara,日本)、0.1%明膠(Sigma,美國)、0.1%多聚賴氨酸(Sigma,美國)、0.1%TritonX-100(Sigma,美國)、4%多聚甲醛(Solarbio,中國)、封閉用山羊血清(中杉金橋,中國)、MHC一抗(Bioss,中國)、Desmin二抗(FITC標記羊抗兔二抗)、3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴鹽(3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide, MTT(Sigma,美國)等。
1.2 骨骼肌衛(wèi)星細胞分離培養(yǎng)
采取腿部肌肉組織,用含有雙抗的PBS溶液沖洗5次,放入無菌培養(yǎng)皿中。剔除肉眼可見的血管和結締組織,用手術剪刀剪成約0.5~1.0 mm的組織塊。先加入0.1%I型膠原酶于37 ℃水浴鍋中消化組織50 min,然后加入0.25%胰蛋白酶(Gibco)消化組織10~20 min(振蕩1次/10 min),在消化完全后加入等體積的完全培養(yǎng)基(DMEM/F12+20% FBS+10%馬血清+100 IU/mL青鏈霉素)終止消化。將消化液過孔徑為200目的細胞篩,濾液1 000 r/min離心10 min,棄上清,得到細胞即為SMSCs。將細胞以1×106個/mL接種在包被的培養(yǎng)皿中(0.1%明膠+0.1%聚賴氨酸),在37 ℃,5% CO2培養(yǎng)箱中培養(yǎng)3 h后,將上清加入到新的培養(yǎng)皿中繼續(xù)培養(yǎng)3 h,后再次吸取上清到新的培養(yǎng)皿中,此時獲得細胞為含雜細胞較少的SMSCs。
1.3 骨骼肌衛(wèi)星細胞傳代培養(yǎng)
原代細胞培養(yǎng)匯合至80%~90%時,吸去培養(yǎng)液,用PBS沖洗3次,加入0.25%胰蛋白酶進行消化約3 min,顯微鏡下待細胞回縮并且少量細胞飄起時,加入等體積的完全培養(yǎng)液終止消化。收集細胞懸液,1 000 r/min離心10 min,棄上清,完全培養(yǎng)基重懸細胞,按1:2或1:3的比例進行傳代培養(yǎng),之后置于37 ℃、5% CO2培養(yǎng)箱中,每隔2 d換液1次,直到細胞生長匯合至90%,并鋪滿整個培養(yǎng)皿。
1.4 細胞生長曲線的測定
原代細胞培養(yǎng)匯合至80%~90%時,經胰蛋白酶消化后收集細胞,接種至96孔培養(yǎng)板內,每孔密度為1.0×105個/mL,隨機分為7組,每組3孔。采用MTT法測定細胞活力:每孔加入MTT 20 μL,37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)4 h,棄MTT溶液,加入DMSO 150 μL,震蕩10 min,在490 nm波長處讀取OD值,每隔48 h進行測定。設置空白調零,計算每孔吸光度值的平均值,繪制曲線,即為骨骼肌衛(wèi)星細胞的生長曲線。
1.5 骨骼肌衛(wèi)星細胞誘導分化
第二代細胞培養(yǎng)至80%匯合度時分別進行成肌和成脂誘導分化。將SMSCs接種于6孔細胞培養(yǎng)皿中,待SMSCs生長融合至80%時,PBS清洗細胞2次,6孔細胞培養(yǎng)皿中加入成肌分化培養(yǎng)基(98% DMEM/F12+2%馬血清)誘導SMSCs成肌分化,37 ℃、5% CO2培養(yǎng)箱中連續(xù)誘導SMSCs成肌分化8 d,每隔2 d更換新鮮培養(yǎng)液。
將SMSCs接種于6孔細胞培養(yǎng)皿中,待SMSCs生長融合至80%時,PBS清洗細胞2次,加入OriCell SD大鼠脂肪間質干細胞成脂誘導分化培養(yǎng)基A液72 h,后換以OriCell B液培養(yǎng)繼續(xù)培養(yǎng)24 h,重復上述處理兩次共8 d。
1.6 骨骼肌衛(wèi)星細胞的免疫熒光鑒定
利用細胞免疫熒光技術對成肌誘導分化4 d后的SMSCs標記蛋白MHC進行鑒定。將SMSCs接種至6孔板內,按上述流程誘導分化4 d后,吸除培養(yǎng)基,4%多聚甲醛室溫固定15 min,PBS清洗3次;0.25%TritonX-100通透15 min,PBS清洗3次;10%山羊血清封閉液封閉1 h,PBS清洗3次;MHC一抗(1:100稀釋)4 ℃孵育過夜;PBS清洗3次, FITC(1:100稀釋)標記的山羊抗兔二抗室溫孵育1 h;PBS清洗3次;DAPI室溫孵育15 min;共聚焦顯微鏡拍照。
1.7 油紅O染色法及鑒定
SMSCs成脂誘導成熟后,棄完全培養(yǎng)基,PBS洗2次。用4%多聚甲醛固定30 min,PBS洗2次,油紅O染色60 min,棄染色液,PBS洗2次,倒置顯微鏡下觀察。用60%異丙醇進行萃取,測定萃取液在510 nm波長處的OD值。
1.8 引物設計及合成
參照GenBank中Pax7、Desmin、MyoG、PPARγ、C/EBPα、FAS基因序列,用Primer Premier 5.0及NCBI軟件設計實時熒光定量PCR特異性引物,引物序列見表1。引物送至生工生物工程(上海)股份有限公司合成。

表1 qRT-PCR引物相關信息Table 1 Related information about the primers used for qRT-PCR
1.9 實時熒光定量PCR
分別在SMSCs成肌、成脂誘導第0、2、4、6、8 d提取總RNA,利用PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)逆轉錄試劑盒將RNA反轉錄為cDNA,以β-actin為內參基因,實時熒光定量RT-PCR法檢測脂肪細胞分化關鍵基因mRNA的相對表達水平。PCR反應體系20 μL:其中Mix 12.5 μL,上、下游引物各1 μL,模板cDNA 2 μL,ddH2O補足體系。PCR反應程序:95 ℃預變性3 min;95 ℃變性15 s,退火30 s,72 ℃延伸30 s,39個循環(huán),65~95 ℃熔解曲線5 s。
1.10 數據處理
數據均以“平均值±標準差”表示,采用GraphPad Prism 5進行統(tǒng)計分析,采用獨立樣本t檢驗進行組間差異分析,顯著性水平為P<0.05。
2 結果與分析
2.1 SMSCs的形態(tài)學觀察
新分離的SMSCs培養(yǎng)2 h后,大部分非特異性細胞已經附著。大多數非附著的SMSCs隨培養(yǎng)基轉移到新的明膠處理的培養(yǎng)板中以促進粘附生長。新分離的SMSCs呈球形,具有較強的折射率。9 h后,細胞開始附著在細胞培養(yǎng)皿上,貼壁細胞呈橢圓形、梭形或多角形(圖1A);2 d后細胞全部附著,細胞逐漸向梭形或多邊形延伸(圖1B);培養(yǎng)至4 d時,細胞形態(tài)與第二天相似,細胞數目增多(圖1C);培養(yǎng)至8 d時,細胞呈現(xiàn)長梭形且平行排列緊密(圖1D)。

圖1 SMSCs的原代培養(yǎng)(100×)A~D. SMSCs原代培養(yǎng)9 h、2 d、4 d和8 dFig.1 Primary culture of SMSCs (100×)A~D. Primary culture of SMSCs for 9 h, 2 d, 4 d, and 8 d, respectively
培養(yǎng)匯合至90%時傳代培養(yǎng),與原代細胞相比,傳代SMSCs的增殖能力明顯增強。在培養(yǎng)皿中,接種第二代細胞第2天,細胞生長旺盛,細胞形態(tài)正常(圖2A)。傳代第4、6天細胞呈現(xiàn)梭形及不規(guī)則三角形,可見部分融合細胞,折射率增加;在培養(yǎng)過程中,發(fā)生了更多的融合(圖2B、圖2C);培養(yǎng)至8 d時,細胞長梭形且平行排列緊密,呈長管狀,體積明顯大于單個SMSCs(圖2D)。

圖2 SMSCs的傳代培養(yǎng)(100×)A~D. 傳代培養(yǎng)2 d、4 d、6 d和8 dFig. 2 Subculture of SMSCs (100×)A~D. Subculture of SMSCs for 2 d, 4 d, 6 d, and 8 d, respectively
2.2 SMSCs的成肌誘導分化及免疫熒光鑒定
體外培養(yǎng)SMSCs至匯合度到60%~70%加入成肌誘導液,第2天開始有方向不一致的SMSCs相互融合,形成短小的肌管細胞(圖3A)。隨著時間的延長,細胞密度增大,細胞之間的融合更為廣泛,肌管細胞的形成明顯增加(圖3B)。誘導至第6天,肌管細胞之間也開始出現(xiàn)相互的連接和融合,連接成片狀、網狀(圖3C)。誘導至第8天,細胞呈長管狀,體積明顯大于單個骨骼肌衛(wèi)星細胞,各肌管細胞之間出現(xiàn)拉網和連接現(xiàn)象,最終彼此融合(圖3D)。在SMSCs成肌誘導分化第8天,利用細胞免疫熒光技術檢測MHC的表達情況,結果顯示在分化第8天可以明顯觀察到呈染色陽性的細胞核(圖3E)和肌管(圖3F)。
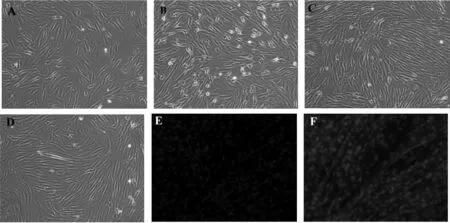
圖3 SMSCs的成肌誘導及鑒定(100×)A~D. 誘導分化2、4、6、8 d;E,F(xiàn). MHC免疫熒光鑒定(誘導分化第8天)Fig. 3 Induced differentiation of SMSCs towards myogenic cells and identification (100×) A~D. Induced differentiation of SMSCs towards myogenic cells at 2, 4, 6, and 8 d, respectively;E,F. MHC immunofluorescence identification (8th day post-induction)
2.3 SMSCs成脂誘導分化及油紅O染色
SMSCs在成脂分化培養(yǎng)基的作用下,隨著時間的增加,長梭形細胞縮短成不規(guī)則三角形及片狀(圖4A和圖4B),在誘導第6天時能觀測到有少量脂滴形成(圖4C),在第8天時細胞內沉積大量脂滴(圖4D)。油紅O染色后,細胞內脂滴被油紅O著色而成紅色,說明SMSCs已經被成功誘導分化為成熟脂肪細胞(圖4E(100×)和圖4F(200×)。
2.4 生長曲線
SMSCs接種后的潛伏期為0~4 d,此時完成細胞的貼壁和伸展。4~6 d進入對數生長期,細胞增殖迅速,第6天后開始進入到平臺期,細胞增殖緩慢。觀察細胞形態(tài)發(fā)現(xiàn),已有部分SMSCs開始相互融合形成散在的肌管細胞(圖5)。
2.5 SMSCs成肌、成脂誘導前后相關基因表達差異比較
SMSCs成肌誘導分化前后Pax7、MyoG和Desmin的表達量有顯著性變化(圖6A)。SMSCs成肌誘導第8天Pax7表達量顯著降低(P<0.05),為誘導第0天的0.08倍;SMSCs成肌誘導第8天MyoG和Desmin表達量顯著增加(P<0.05),分別是誘導第0天的11.03倍和5.48倍。SMSCs成脂誘導分化第8天與誘導第0天相比,PPARγ表達量增加,但差異不顯著(P>0.05);C/EBPα和FAS的表達量均顯著增加(P<0.05)(圖6B);SMSCs成脂誘導第8天C/EBPα和FAS表達量分別是誘導第0天的5.51倍和2.01倍。
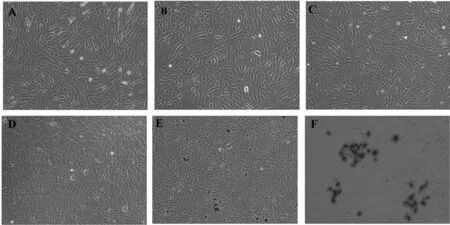
圖4 MDSCs的成脂誘導及鑒定A~D. 誘導分化2、4、6、8天;E,F(xiàn). 油紅O染色(誘導分化第8天)Fig. 4 Induced differentiation of MDSCs towards adipogenic cells and identificationA~D. Induced differentiation of MDSCs towards adipogenic cells at 2, 4, 6, and 8 d, respectively;E,F. Oil red O staining (8th day post-induction)
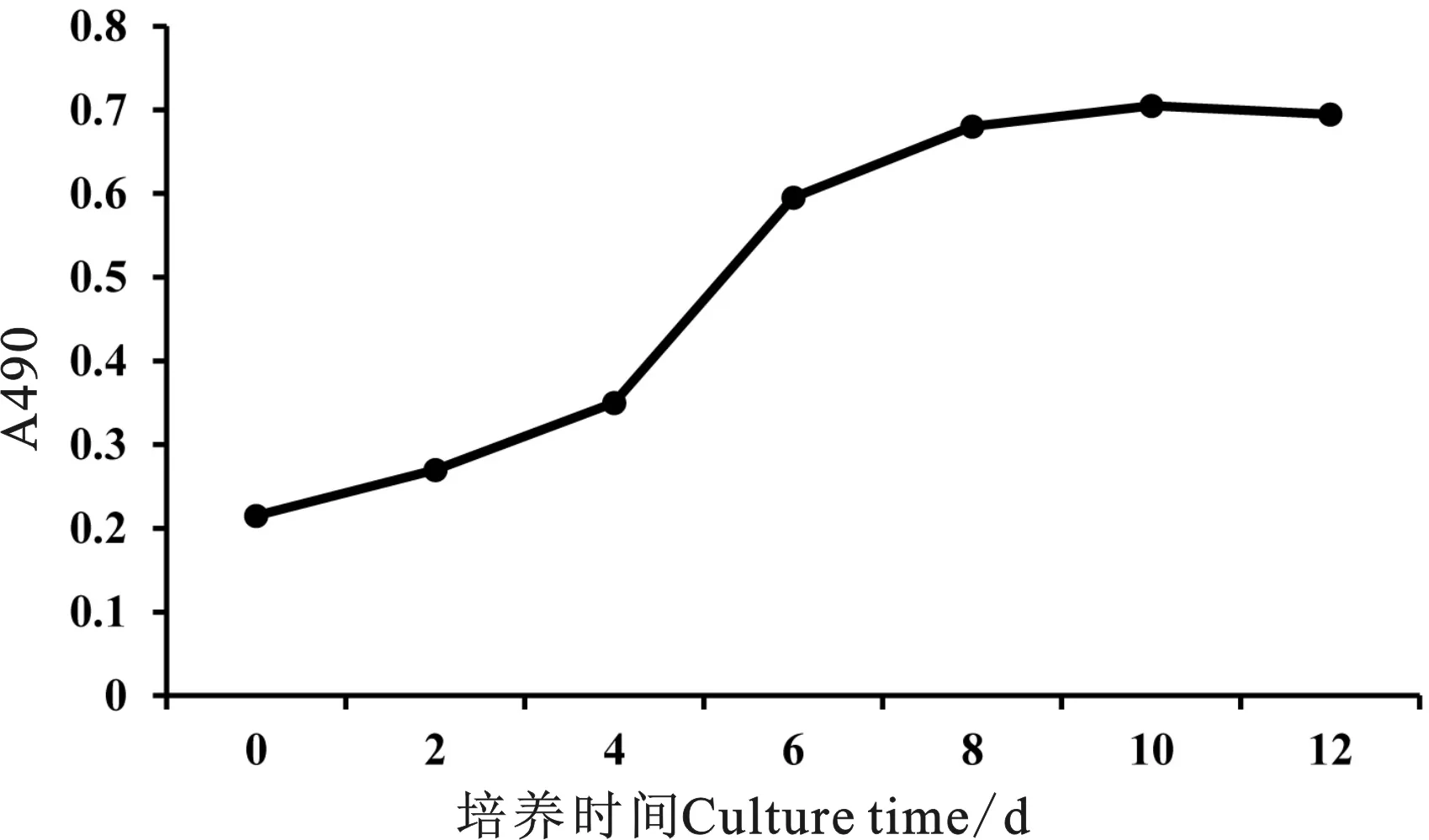
圖5 SMSCs生長曲線Fig. 5 The growth curve of SMSCs
3 討 論
3.1 SMSCs分離培養(yǎng)
SMSCs是位于肌膜和基底膜之間的一類肌源性干細胞,直接參與骨骼肌分化,是脊椎動物出生后調節(jié)肌肉再生長的最主要方式。一般情況下,SMSCs處于休眠狀態(tài),當肌肉受到損傷或者受到外界信號的刺激誘導后,肌肉組織的再生主要由肌衛(wèi)星細胞完成的。SMSCs在幼齡動物肌肉組織中占的比例較高,但是其總體數量和肌肉組織細胞占比會隨著動物年齡增加而不斷減少[15]。目前,羊、牛、豬、雞等畜禽的SMSCs的分離培養(yǎng)已經被報道。SMSCs的分離方法主要有組織塊培養(yǎng)法和酶消化法兩種,其中酶消化分離法主要有膠原酶法和胰蛋白酶法等[16-19]。本研究利用0.1%I型膠原酶和0.25%酶兩步消化法成功地分離了灘羊SMSCs,并對其進行了細胞學鑒定。鄭琪等[20]使用10%馬血清的培養(yǎng)基對安淮山羊骨骼肌衛(wèi)星細胞進行分離培養(yǎng);解一凡等[21]采用15%馬血清血清培養(yǎng)基對哈薩克羊骨骼肌衛(wèi)星細胞進行了分離培養(yǎng)。本研究采用10%馬血清的培養(yǎng)基對灘羊骨骼肌衛(wèi)星細胞進行了分離培養(yǎng),并進一步利用差速貼壁法對細胞進行純化。兩步消化法使得細胞充分釋放,差速貼壁法可以去掉成纖維細胞,獲得的骨骼肌衛(wèi)星細胞經過傳代后純度較高,增殖和分化能力較強。新分離的、未激活的SMSCs處于休眠狀態(tài),很難附著在未處理的培養(yǎng)皿底部[21],本研究采用0.1%明膠和0.1%聚賴氨酸對細胞培養(yǎng)皿進行包被,明膠和聚賴氨酸有助于綿羊SMSCs的附著。
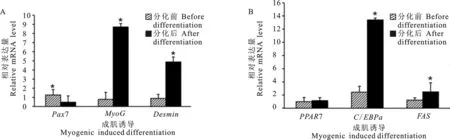
圖6 SMSCs成肌、成脂誘導前后相關基因的表達“*”表示2組數據差異顯著(P<0.05)Fig. 6 Expression of genes involved in myogenic and adipogenic potentials of SMSCs before and after induced differentiation“*” indicates significant difference between two data sets (P<0.05)
3.2 SMSCs的誘導分化及鑒定
SMSCs在未激活和已激活狀態(tài)表達不同的基因或蛋白質,其中一些已經被用作標記物利用免疫熒光、蛋白印記等技術對SMSCs進行分子鑒定或者細胞狀態(tài)的識別。目前,研究證實SMSCs表達骨骼肌標志性蛋白如MHC、Desmin、Pax7、MyoD、MyoG等[16,22-24],已經證實SMSCs能夠合成骨骼肌特異性的蛋白。例如,MHC作為表皮生長因子超家族的一員,已被證明在骨骼肌發(fā)育中發(fā)揮重要作用,常作為成肌標記基因。正常的成肌細胞中會表達MHC,利用細胞免疫熒光技術對MHC的表達進行檢測,可以說明該細胞是否為成肌細胞。本研究結果表明誘導分化的細胞中MHC的表達呈陽性,證實分離得到的SMSCs可誘導分化成為成肌細胞。SMSCs除了具有成肌潛力以外,還具有向成脂細胞和成骨的潛力。本研究采用成脂誘導劑誘導灘羊SMSCs分化為類脂肪細胞,油紅O染色鑒定發(fā)現(xiàn),誘導分化培養(yǎng)8d后細胞內有明顯脂肪滴形成,表明分離的灘羊SMSCs亦具有成脂細胞的潛力,可以作為細胞模型探究相應的調控機制。
3.3 SMSCs成肌、成脂誘導前后相關基因表達差異比較
SMSCs從靜息狀態(tài)到激活狀態(tài),再從激活狀態(tài)到成肌分化形成成熟的肌纖維受到有序復雜基因網絡的調控,精準控制SMSCs發(fā)揮其生物學功能。靜止和激活SMSCs最廣泛使用的標記物是Pax7,這是一種屬于參與肌肉發(fā)育的螺旋-轉-螺旋-配對盒家族的轉錄因子[25]。Myogenin(MyoG)是最后的MRFs之一,參與了發(fā)育和再生過程中肌肉分化的晚期階段,特別是在成肌細胞肌管和成熟纖維的形成中[26]。隨著MyoG的表達,細胞進入生肌分化階段,最后MHC基因表達,標志著生肌分化進入最后時期[27]。結蛋白(desmin)是中間絲蛋白的一種,是靜止的衛(wèi)星細胞和增殖的成肌細胞標志之一,在損傷后再生的肌纖維以及雞、大鼠和小鼠等的胚胎成肌纖維中表達[28]。肌細胞的生成和分化過程由Pax7和MRFs家族成員相互作用共同參與,Pax7在靜止的肌衛(wèi)星細胞中表達, 激活后則同時表達Pax7和MyoD。肌衛(wèi)星細胞激活后,Pax7正調控下游基因Myf5、MyoD、myogenin和MRF4,它們介導骨骼肌標志基因的表達并誘導肌衛(wèi)星細胞分化[29]。本研究發(fā)現(xiàn),SMSCs成肌誘導分化前后Pax7表達量顯著降低,而MyoG和Desmin的表達量表達量顯著增加。原代山羊肌衛(wèi)星細胞成肌分化過程中Pax7基因顯著下調,MyoG基因與Desmin基因顯著上調[30], 這與本試驗結果一致。靜息狀態(tài)下的肌衛(wèi)星細胞表達Pax7,進入活化階段后,Pax7表達下調,最終分化進入晚期階段,MHC開始表達,說明肌細胞逐漸停止增殖,并開始分化、融合。研究證明SMSCs能在體外分化為成脂細胞[31-32]。PPARγ和C/EBPα是參與脂肪細胞成熟的關鍵轉錄調控因子,C/EBPα被激活后能上調其他成脂分化相關基因的表達,從而促進細胞的成脂分化[30,33]。本研究發(fā)現(xiàn),SMSCs成脂誘導分化后C/EBPα和FAS的表達量顯著增加,說明C/EBPα激活后上調FAS的表達量,促進SMSCs的成脂分化。這說明通過改變誘導培養(yǎng)條件,骨骼肌衛(wèi)星細胞能夠分化成為脂肪細胞,并且成脂相關的基因PPARγ、C/EBPα和FAS的表達量均呈現(xiàn)了上升趨勢,此結果與細胞染色結果完全相符。這可能是由于部分細胞已分化為類脂肪細胞,成脂標志性基因開始特異性表達,也可能受到脂肪酸沉積的影響,同時也可能受到PPARγ和C/EBPα的調控。
4 結 論
本研究通過I型膠原酶和胰蛋白酶消化分離骨骼肌衛(wèi)星細胞,并利用差速貼壁法對細胞進行純化,所分離的細胞經過傳代后純度較高,增殖和分化能力較強。成肌和成脂誘導分化分別通過免疫熒光和油紅O染色鑒定所分離的細胞為骨骼肌衛(wèi)星細胞,并且具有成肌和成脂分化能力。MDSCs成肌誘導分化前后Pax7、MyoG和Desmin的表達量有顯著性變化,MDSCs成肌誘導第8天Pax7表達量顯著降低,MyoG和Desmin表達量顯著增加;MDSCs成脂誘導分化第8天與誘導第0天相比;C/EBPα和FAS的表達量顯著增加。灘羊骨骼肌衛(wèi)星細胞的成功分離培養(yǎng)為灘羊骨骼肌的發(fā)育及再生機制,提供了可靠的細胞模型。
