山桐子果實發育過程中內含物、內源激素及光合特性的變化*
2021-01-05 02:47:18房麗莎徐自恒耿曉東蔡齊飛翟雯婧周海清王艷梅
林業科學 2020年11期
關鍵詞:生長
房麗莎 徐自恒 劉 震 李 志 耿曉東 蔡齊飛 翟雯婧 周海清 王艷梅
(1. 河南農業大學 鄭州 450002; 2. 河南伏牛山國家級自然保護區黑煙鎮管理局 南陽 474550)
近些年,我國年食用植物油消費量在3.8×107t左右,排名世界第一,而國產食用油自給率僅占31%,這就意味著我國約69%的食用油依賴進口。新型、特色或功能性食用植物油的開發生產有助于滿足對食用植物油不斷增長的需求,而木本油料是食用植物油的開發生產重要來源(吳晶晶, 2020)。
山桐子(Idesiapolycarpa)是大風子科(Flacourtiaceae)山桐子屬(Idesia)落葉喬木。果實(漿果)含油率高且營養豐富,其種子、果肉、果實的含油率分別可達28.17%、48.35%、39.40%,不飽和脂肪酸含量可達90.68%、86.66%、88.67%,亞油酸分別在80%、60%、70%以上(代莉, 2014)。山桐子果實高產高質,可作為開發高檔食用植物油的優良原料,有利于維護國家食用油戰略安全; 食用山桐子油可預防多種心腦血管疾病; 山桐子果實可入藥,其深加工提取的亞油酸和共軛亞油酸是醫藥和保健食品的良好原料(代娟等, 2016); 山桐子油不但可用作制造肥皂、制漆及防水涂料等,也可被加工利用制備生物柴油和潤滑油; 榨油后的油餅可以加工做飼料,也可提取驅蟲劑。山桐子作為木本油料樹種,抗性強、耐貧瘠,適應范圍廣,具有“不與人爭糧、不與糧爭地”的優勢,在食品、醫藥、化工及林業等多領域具有巨大的的開發潛能及應用前景。
光合作用不僅是植物生長發育和干物質積累的基礎,而且與果實的生長發育和產量有著密切的關系(王瑞等, 2013)。研究果實生長發育過程中光合生理的變化,掌握果實不同生長發育時期對光環境的適應變化規律,可為后期采取合理的栽培管理措施及充分利用光合潛能實現優質增產的目的提供理論依據(彭邵鋒等, 2012)。在黃連木(Pistaciciachinensis)和白檀(Symplocospaniculata)果實發育的前期,光合產物主要以可溶性糖和淀粉的形式積累,而在果實發育中后期轉化為蛋白質和脂肪(齊昆等, 2011; 劉倩倩等, 2017)。超氧化物歧化酶(SOD)、過氧化物酶(POD)及過氧化氫酶(CAT)等抗氧化酶對抵抗外界環境傷害起到積極作用(楊盛等, 2019)。內源激素對果實的生長發育及其品質具有重要的調控作用(張淑文等, 2019; Mcateeetal., 2013)。在果實發育前期,GA3與ZR對果實的膨大生長有一定影響; 在果實發育后期,低濃度的IAA、GA3和高濃度的ABA會造成落果; 另外ABA能促進花色苷的合成從而影響果實的變色(吳尚等, 2017; Jiaetal., 2011)。
目前,山桐子的研究主要集中在生物學特性(劉芙蓉等, 2017)、苗木繁育與栽培(楊恩讓等, 2013)、油脂提取工藝(華婉等, 2016)和生物活性成分測定等(代娟等, 2016),對山桐子果實發育機制研究甚少。本研究測定山桐子果實生長發育過程中內含物、內源激素及葉片的光合變化,為山桐子果實發育機制及調控技術研究提供依據。
1 材料與方法
1.1 試驗材料
以2013年栽植于河南省鄭州市河南農業大學林學試驗站(113°38′E,34°47′N)的4年生成都種源山桐子為試驗材料,株行距為4 m×4 m。
1.2 果實內含物及內源激素的測定
2017年以栽植于2013年的苗木為材料,根據果序長度、果實橫縱徑及顏色變化將果實生長過程分為果實發育的快速生長期(Ⅰ)(5月26日—7月6日)、生長緩慢期(Ⅱ)(7月6日—8月16日)、顏色變化期(Ⅱ)(8月17日—10月15日)和成熟期(Ⅳ)(10月16日起)(圖1)。
于6月16日、7月16日、9月16日和10月16日,在山桐子標準木的東、西、南及北方向隨機各采摘無病蟲害大小基本一致的20粒果實。采摘后用液氮速凍,然后置于-80 ℃條件下保存,用于抗氧化酶、內含物及內源激素測定。
SOD活性、POD活性、CAT活性、丙二醛(MDA)含量測定參照高俊鳳(2000),測定方法分別采用氮藍四唑光化還原法(NBT)、愈創木酚法、紫外吸收法、硫代巴比妥酸(TBA)法; 可溶性蛋白含量測定采用考馬斯亮藍G-250染色法(路文靜等, 2012),可溶性糖的測定采用蒽酮法(李合生, 2000)。IAA、ABA、GA3及ZR含量的測定使用高效液相色譜儀(Agilent1260 HPLC,Agilent Company,USA),參照Li等(2011)的方法。每次取樣0.5 g,并重復3次。

圖1 山桐子果實生長發育過程Fig.1 The fruit growth and development of Idesia polycarpa
1.3 光合指標測定
1.3.1 葉片光合參數測定 在6月18日、7月15日、9月18日和10月18日(晴朗、無風),使用LCpro-SD便攜式光合儀,以標準木主莖新梢向陽中部健康的成熟葉片為測試葉,從早上8:00至下午6:00每隔2 h測定1次凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)及胞間CO2濃度(Ci)。每次重復測定3次。
1.3.2 葉片光響應曲線和CO2響應曲線測定 光響應曲線采用LCpro-SD光合儀自身配備的紅藍光源為光源,光合有效輻射(PAR)均設置為0、50、100、200、400、600、800、1000、1 200和1 500 μmol·m-2s-1,共10個光強梯度,參比室CO2濃度600 μmol·mol-1進行測定,得到所對應的Pn值。
CO2響應曲線以光合儀自身配備的紅藍光源為光源,以CO2小鋼瓶作為氣源,設定各種源CO2濃度梯度均為: 0、50、100、200、400、600、800、1 000、1 200和1 400 μmol·mol-1,PAR設定為1 000 μmol·m-2s-1,得到所對應的Pn值。
1.4 數據分析
采用Microsoft Excel 2010進行數據整理,SPSS 21.0數據分析軟件對相關指標進行單因素方差分析(One-way ANOVA),運用Duncan新復極差進行多重比較,采用直角雙曲線修正模型(葉子飄, 2012)擬合光響應曲線和CO2曲線,Pearson相關性檢驗分析內含物、內源激素以及光合參數的關系,并采用OriginPro 9.0制圖。
2 結果與分析
2.1 山桐子果實不同發育階段抗氧化酶和內含物變化
SOD是植物體內清除自由基的最關鍵酶類之一,為典型的誘導酶,外部環境的改變能影響其活性水平。由圖2可知,在果實生長發育過程中,SOD活性呈“下降-上升”趨勢,在快速生長期(Ⅰ)SOD活性較高,為127 U·g-1min-1,可能是受到高溫或缺水等不良環境影響; 在果實成熟期(Ⅳ)也處于較高水平,為121 U·g-1min-1,可能與環境及果實發育進程有關。POD和CAT是清除H2O2的關鍵酶。POD活性呈“下降-上升”的趨勢,CAT活性呈“上升-下降”趨勢,但果實成熟期(Ⅳ)與顏色變化期(Ⅱ)差異不顯著(P>0.05),二者共同作用清除H2O2。保護酶系統以SOD為主,在顏色變化期(Ⅱ)SOD活性最低,可能誘發膜脂過氧化作用,導致MDA的增加。可溶性糖含量和可溶性蛋白變化趨勢基本一致,呈“上升-下降”趨勢。
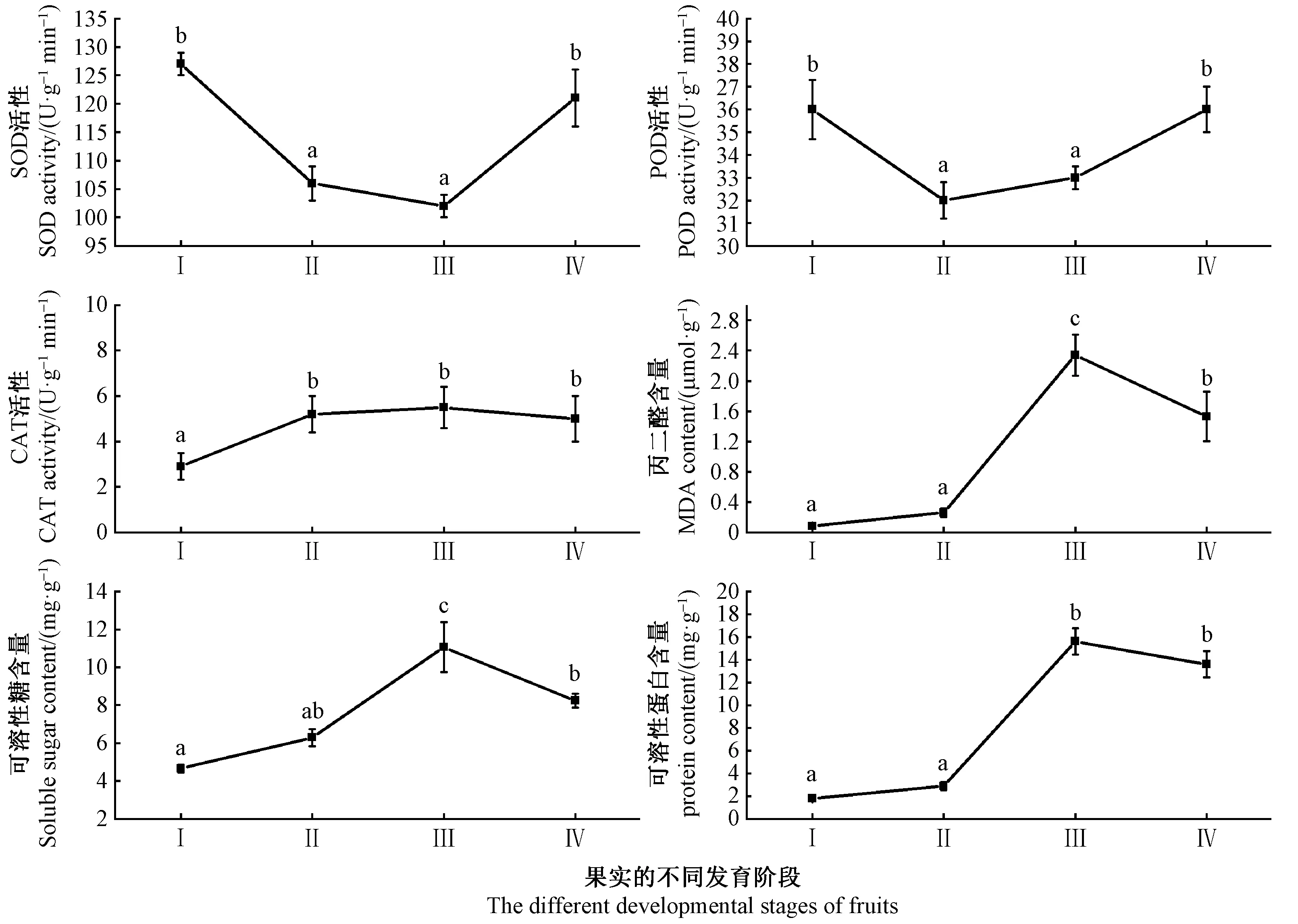
圖2 山桐子果實的不同發育階段抗氧化酶和內含物的變化Fig.2 The changes of the activity of antioxidant enzymes and contents at different fruit developmental stages of I. polycarpa
2.2 山桐子果實不同發育階段內源激素變化
由圖3可知,ABA含量從快速生長期(Ⅰ)到生長緩慢期(Ⅱ)有小幅度下降,但二者差異不顯著(P>0.05); 之后ABA含量處于上升趨勢,在果實成熟期(Ⅳ)ABA含量達到最高,且差異顯著(P<0.05),此時期果實已經成熟,這說明高含量的ABA起促進果實成熟的作用。從快速生長期(Ⅰ)到顏色變化期(Ⅱ)GA3含量升至最高41.05 μg·g-1,差異顯著(P<0.05),到果實成熟期(Ⅳ)GA3含量緩慢下降,差異不顯著(P>0.05); IAA含量呈“下降-上升”趨勢,在快速生長期(Ⅰ)IAA含量達到最高,顏色變化期(Ⅱ)IAA含量降至最低0.01 μg·g-1,且差異顯著(P<0.05),IAA含量變化與果實的生長發育規律相符; 快速生長期(Ⅰ)到生長緩慢期(Ⅱ)ZR含量下降至最低0.05 μg·g-1,與其他發育時期相比差異顯著(P<0.05),在果實顏色變化期(Ⅱ)又迅速上升; 在果實成熟期ZR含量升至最高,這可能與后期成熟果實中種子同化物的積累有關。
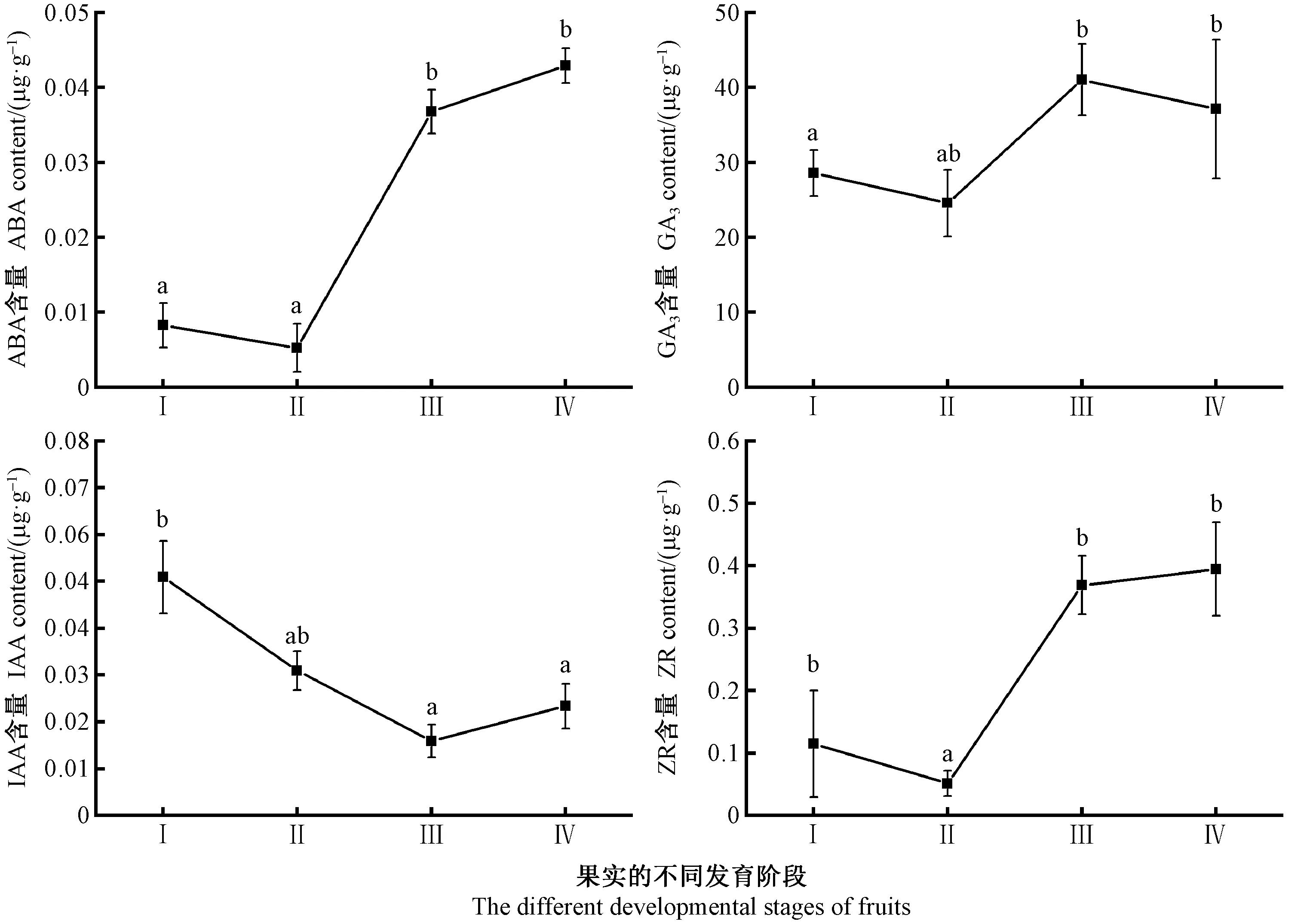
圖3 山桐子果實的不同發育階段內源激素變化Fig.3 The changes of content of endogenous hormones during different fruit developmental stages of I. polycarpa
2.3 山桐子不同發育階段光合特性分析
2.3.1 光合參數分析 由圖4可知,山桐子凈光合速率(Pn)在果實不同發育階段的日變化曲線呈不對稱的雙峰變化趨勢,在10:00達到最大峰值為9.4 μmol·m-2s-1,之后Pn逐漸下降,有明顯的光合“午休”現象,這可能是由于中午的光強過高,造成葉片氣孔關閉; 在16:00時又出現1個高峰,之后繼續下降。在果實生長緩慢期(Ⅱ)Pn值最高,在果實成熟期(Ⅳ)Pn值最低,主要是由于此時期的葉片進入衰老階段,光合強度下降導致。蒸騰Tr總體變化趨勢為先上升后下降的倒“V”形,由于上午溫度低、光照弱、空氣濕度大,Tr較低; 中午隨著溫度上升、光強增大,Tr在果實發育各階段均在14:00達到最大值,之后又隨著溫度和光強的下降,Tr降低。在生長緩慢期(Ⅱ)Tr值整體趨勢最高,在果實成熟期(Ⅳ)Tr值整體趨勢最低。葉片胞間CO2濃度(Ci)變化趨勢基本一致,果實發育的同一時期Ci日變化趨于平穩。在果實成熟期(Ⅳ)日變化Ci值最大,在果實生長緩慢期(Ⅱ)日變化Ci值最小。山桐子葉片氣孔導度(Gs)隨時間變化呈現出雙峰曲線的變化規律,Gs在10:00時出現最大值為0.3 μmol·m-2s-1,之后下降,這可能是由于中午具有較高的光強和溫度,Tr達到最大,為防止水分過度散失,植物會關閉氣孔。在果實快速生長期(Ⅰ)Gs高于其他時期,在果實顏色變化期(Ⅱ)和成熟期(Ⅳ)Gs值差異不明顯。
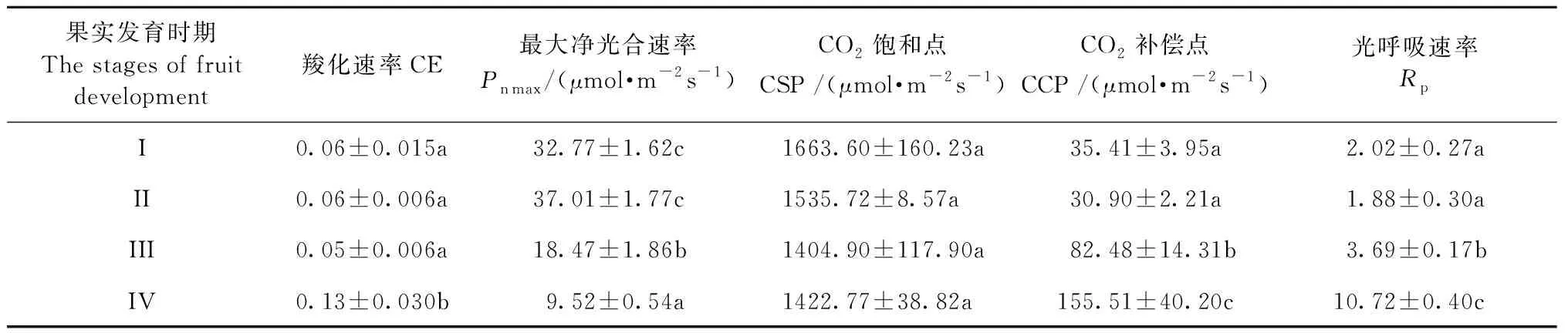
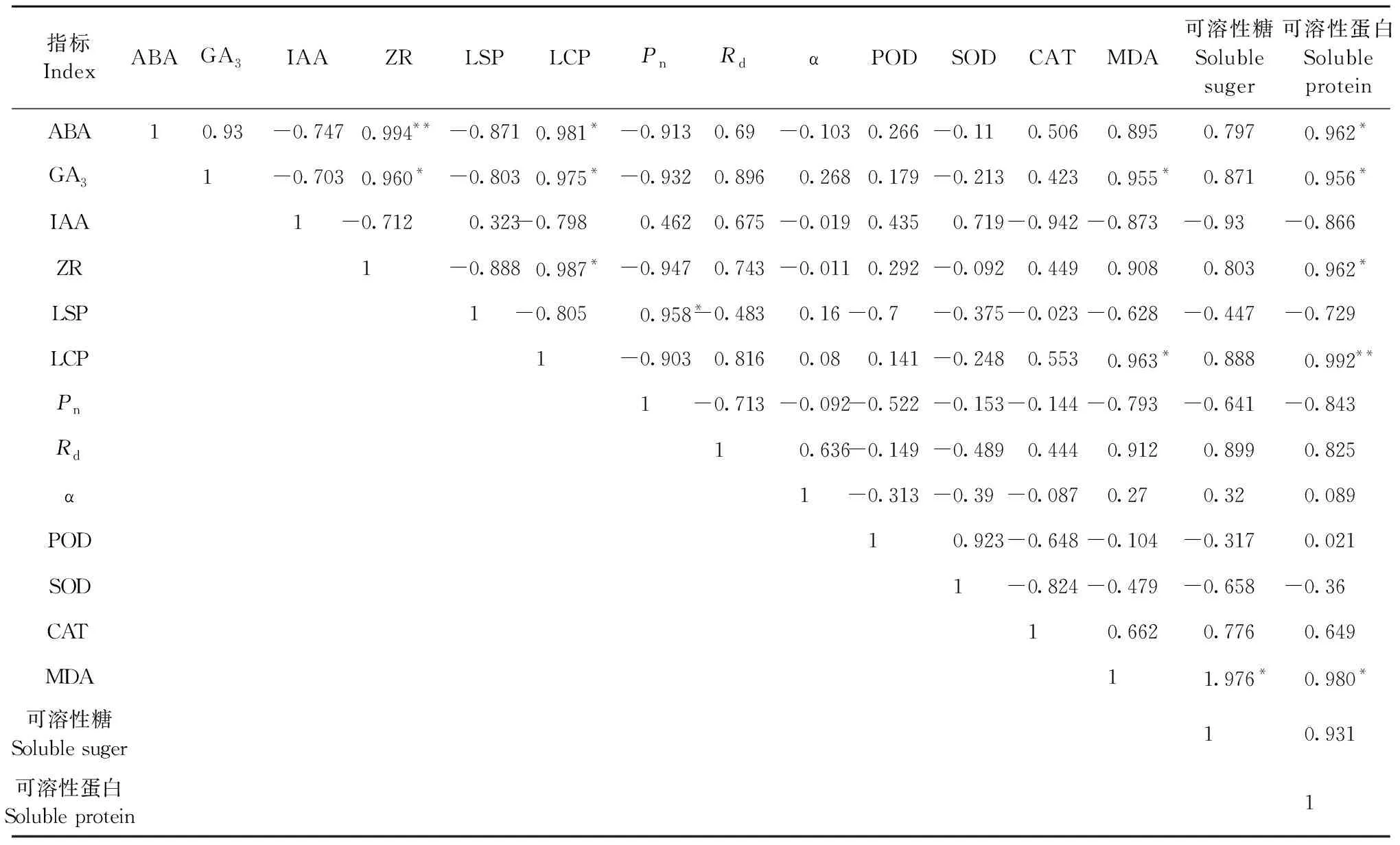
2.3.2 光響應曲線分析 由圖5可知,山桐子果實在不同發育階段光響應曲線變化趨勢基本一致,在光合有效輻射(PAR)≤200 μmol·m-2s-1時,Pn隨著PAR增加呈線性增加; 在200 μmol·m-2s-1 2.3.3 葉片CO2響應曲線 由圖5可知,山桐子CO2響應曲線變化趨勢基本一致,Ci≤ 800 μmol·mol-1時,Pn隨著Ci的增大幾乎呈直線趨勢; 800 μmol·mol-1 圖4 山桐子果實不同發育階段的不同光合參數日變化Fig.4 Diurnal variation of photosynthetic parameters in different fruits developmental stages of I. polycarpa 表1 山桐子果實發育不同階段光響應曲線特征參數①Tab.1 The light response curve characteristic parameters during different development stages of I. polycarpa 表2 山桐子果實發育不同階段CO2響應曲線特征參數Tab.2 The light response curve characteristic parameters during different development stages of I. polycarpa 山桐子果實不同發育階段的內含物、內源激素和光合參數兩兩進行相關性分析。由表3可知,可溶性蛋白與ABA、GA3和ZR呈顯著正相關; MDA與LCP呈顯著正相關; 可溶性蛋白與LCP呈極顯著正相關; ABA、GA3和ZR與LCP呈顯著正相關。 表3 山桐子不同發育時期內含物、內源激素及光合參數相關性①Tab.3 The correlation analysis between inclusions,endogenous hormone and photosynthetic parameter during different fruit developmental stages of I. polycarpa SOD、POD及CAT是植物體內維護細胞微環境重要的抗氧化酶,SOD通過歧化反應將對植物有害的超氧化自由基轉化為O2和毒性較低的H2O2,然后通過POD、CAT作用分解為H2O,其活性的高低意味著對膜系統保護能力的強弱(許娟等, 2015)。山桐子果實快速生長期SOD和POD活性較高,可能是由于果實膨大速度較快,細胞透性增大,易受環境影響,誘導抗氧化酶升高,阻止膜脂過氧化作用,維持細胞微環境。這與杏(Armeniacavulgaris)果實發育前期抗氧化酶的變化研究結果一致(李萍等, 2013)。壺瓶棗(Ziziphusjujuba‘Huping’)的半紅期果皮MDA明顯升高,抗氧化酶顯著降低,破壞了細胞膜系統,最終引起棗裂果(丁改秀等, 2014); 艷紅桃(Amygdaluspersica‘Yanhong’)果實MDA含量在成熟期達到最高,有助于果實成熟軟化(張蘭蘭等, 2012)。在山桐子果實發育后期MDA含量明顯上升,表明此時果實內可能含有較高活性氧,有助于乙烯的生物合成,為山桐子果實成熟奠定基礎。 不同的植物果實中油脂和糖類、蛋白質生物合成之間的關系不但受遺傳因素的影響,而且受發育階段的影響(劉倩倩等, 2017)。果實快速生長需要消耗大量糖類,山桐子果實快速生長期可溶性糖含量低,這與油茶(Camelliaoleifera)幼果發育的可溶性糖變化一致(張凌云等, 2013)。文冠果(Xanthocerassorbifolia)油脂快速累積之前,可溶性糖水平較高,隨后其含量隨著油脂的快速累積幾乎呈線性下降,脂肪含量與糖類含量變化呈負相關而與蛋白質含量變化呈正相關,油脂合成主要來源于葉片光合作用的可溶性糖(趙娜等, 2015)。山桐子的油脂積累主要發生在7月中旬到9月下旬(祝志勇等, 2010),在果實生長緩慢期和顏色變化期油脂逐步積累,而可溶性糖含量顯著升高,表明山桐子油脂積累方式可能與文冠果有差異。 在果實的生長發育過程中,內源激素對果實生長、油脂的合成和果實著色有一定的調控作用(廖明安等, 2009)。IAA能夠促進細胞分裂、縮短細胞分裂周期并參與果實膨大生長(翟開恩等, 2015), 在梨(Pyrus)、沙棘(Hippophaerhamnoides)果實發育前期和中期,IAA含量保持在較高水平(Zhangetal., 2011; 杜維等, 2018),這與山桐子果實的IAA變化一致,促進了果實的膨大生長。ZR會抑制細胞壁酶活性,影響果實種子中同化物的積累和胚乳發育(周蕾等, 2006),表現出細胞壁緊湊和果實膨大的現象(楊曉婉等, 2014),山桐子果實快速生長期的ZR含量顯著高于生長緩慢期,這有利于促進山桐子果實的膨大生長。GA3不僅能有效誘導漿果的增大,并且可通過降低花色素苷的合成,抑制漿果的成熟,防止果實的脫落(Zeretal., 2012; Mesejoetal., 2016)。對赤霞珠葡萄(Vitisvinifera‘Cabernet Sauvignon’)和樹莓(Rubuscorchorifolius)的研究發現GA3在果實發育前期保持較高濃度,促進果實膨大,增加單果質量,在果實成熟期GA3含量明顯降低(高江曼等, 2017; 高慶玉等, 2015)。而山桐子果實成熟期GA3雖呈下降趨勢,但GA3含量仍保持較高濃度,這與不易落果的楊梅(Myricarubar)果實成熟期較高的GA3含量一致(張淑文等, 2019)。山桐子在果實成熟后,果實長時間不脫落,并且有的果實可掛于枝端至翌年3月或4月,這可能與其GA3含量較高有關。ABA參與果實成熟的啟動調控,ABA亦能促進果實花色素苷的積累從而影響果實的顏色變化,進而促進果實的成熟(Shenetal., 2014; Hao etal., 2014),且高濃度的ABA能促進油脂(杜維等, 2018)和可溶性糖含量的積累(趙曉曉等, 2019),因此山桐子果實變紅、成熟、油脂增多及可溶性糖含量上升可能與ABA含量顯著上升有關。相關性分析表明ABA、GA3與可溶性蛋白均呈顯著正相關,可能ABA、GA3在調控果實的變紅、成熟以及油脂的增多過程中需要大量的酶類參與,而可溶性蛋白作為植物體內多種酶的主要組成部分,調控多種生理生化代謝過程(冷熹鳴等, 2019)。 植物果實發育所需的營養物質主要來源于葉片的光合作用。山核桃(Caryacathayensis)果實膨大期,Pn和Tr達到最高(Xuetal., 2016); 高產油茶無性系(XL67)的Pn和Tr的變化一致,較高的Tr使葉片免于高溫和強光的灼傷,維持葉片在高溫下進行光合作用(彭邵鋒等, 2012)。本研究中山桐子果實緩慢生長期的Pn在日變化過程中達到最高,此時光照強度和溫度都達到全年最高,Tr也達到最高,保證山桐子在強光和較高的溫度下進行光合作用。在果實顏色變化期和成熟期Pn相對下降,此時期果實ABA的濃度較高,研究表明較高的ABA濃度能夠抑制葉片的生長(Daviesetal., 2003); 同時山桐子果實成熟過程中乙烯(ET)含量會增多,ET不僅具有信號傳導的作用,還能夠與脫落酸(ABA)共同調控氣孔開度,促進葉綠素降解等作用,從而使葉片光合速率下降,光合產物生成減少(張小娟等, 2017)。這可能也是在果實顏色變化期和成熟期山桐子葉片的光合能力減弱的重要原因之一。 光響應曲線中,LSP和LCP反映植物的需光性和弱光的利用能力,較高的LSP和較低的LCP表明植物光能利用范圍大,具有較高的生態適應性(黃秦軍, 2013)。Pnmax較高表明植物的光能利用效率高,本文中山桐子果實生長緩慢期(Ⅱ)LSP和Pnmax最高、Rd和LCP最低,表明山桐子在高溫和強光下保持較高的光能轉化率以及較寬的光能利用范圍,這與張磊等(2014)研究結果一致,說明山桐子對光照的適應性相對較強。葉片光合生理指標影響果實的品質,相關研究表明Pn與果實的可溶性糖呈顯著正相關,Pn越高,光合產物越多,積累更多的可溶性糖(申強等, 2020)。在本研究中Pn與可溶性糖無顯著正相關關系,可能是由于果實的發育和葉片的光合都處于動態的變化,存在相關性但不顯著。LCP與ABA和GA3呈顯著正相關,說明葉片的LCP的變化顯著影響果實中ABA和GA3的變化,可能是由于LCP越高,弱光利用能力越差,光合產物向果實的輸入降低,而果實發育后期需要充足的碳源物質,高濃度的ABA和GA3通過調控糖代謝相關的酶活性,從而提高果實中的可溶性糖的含量(沙建川等, 2019)。CO2是綠色植物進行光合作用的重要原料,其通過提高Rubisco酶結合位點的羧化速度和抑制呼吸作用來增高凈光合速率(李麗霞等, 2016)。CSP表征對高CO2的利用能力,CCP表征對低濃度CO2的同化能力,Rp代表光呼吸速率的大小(葉子飄, 2010)。果實生長緩慢期(Ⅱ)較高的Pnmax和CSP以及較低的CCP和Rp表明此時期有較寬的CO2利用區間和較高的CO2同化能力。 適當的水分能保持較高的光合作用,而且漿果的生長期水分虧缺會降低含糖量和可溶性固形物,影響果實的品質(陳婧等, 2013; 牛最榮等, 2020); 當土壤含水量在46.3%~81.9%,山杏(Prunussibirica)就維持較高的Pn和CE,沒有明顯的CO2飽和抑制現象,當土壤含水量低于或高于這個范圍,就會明顯的降低光合作用(吳芹等, 2013)。李晶等(2018)認為在葡萄果實成熟期水分虧缺可以提高還原糖和可溶性固形物,并降低可滴定酸,提高葡萄的品質; 而對油料樹種油茶的油脂積累期適當的水分灌溉可以提高果實的含油率(徐猛, 2014)。因此在田間栽培管理時,山桐子果實生長期應及時灌溉補充水分,防止光合效率下降,提供充足的光合產物來保證果實的品質,而對于果實發育后期是否需要水分灌溉,還需要進一步的試驗證明。 山桐子果實發育過程,SOD、POD及CAT協同發揮作用,以維持活性氧的形成和清除之間的平衡,保證山桐子果實在不同時期的正常生長發育。ABA、IAA、GA3、ZR含量的變化動態與果實的發育規律相吻合,對果實的發育進程起積極的調控作用。在山桐子的生長緩慢期有較大的光能和CO2利用范圍以及較高的凈光合速率,但此時高溫干旱,氣孔導度增加,蒸騰速率也達到最高,應進行及時灌溉,以便維持較高的光合速率,為山桐子提供更多的光合產物來保證果實的優良品質。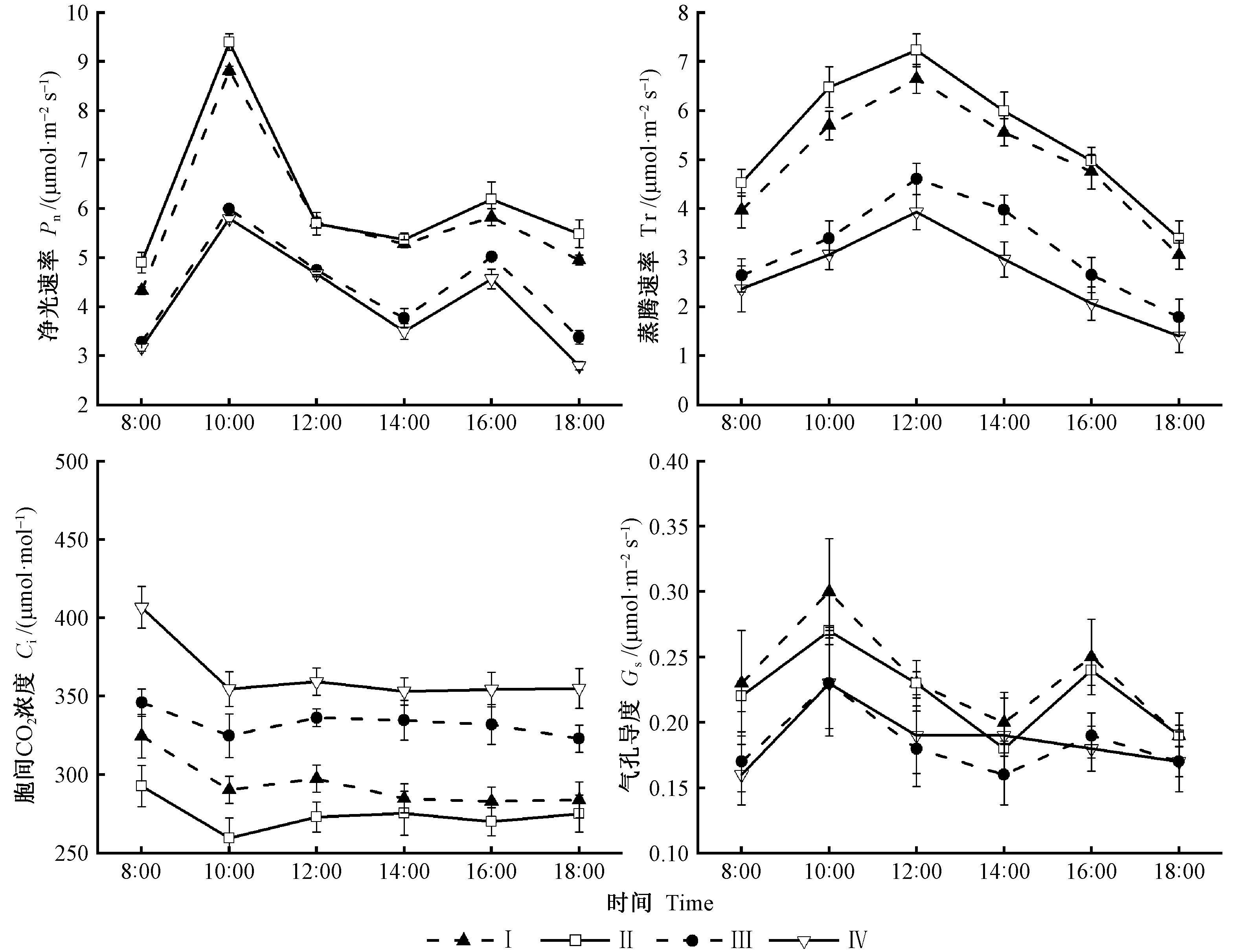

2.4 山桐子果實內含物、內源激素和光合參數的相關性
討論
3.1 果實不同發育階段內含物的變化
3.2 果實不同發育階段內源激素的變化
3.3 果實不同發育階段光合特性的變化
4 結論
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00小讀者(2021年2期)2021-03-29 05:03:48少兒美術(2020年3期)2020-12-06 07:32:54現代裝飾(2020年11期)2020-11-27 01:47:48中學生天地(A版)(2020年3期)2020-04-10 10:57:45故事作文·高年級(2020年3期)2020-03-17 09:24:33瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16華人時刊(2019年13期)2019-11-17 14:59:54NBA特刊(2018年21期)2018-11-24 02:48:04文苑(2018年22期)2018-11-19 02:54:14
