林下植被清除對不同徑級中齡杉木生長速率的影響機制*
2021-01-05 02:47:16陳奕帆付曉莉王輝民戴曉琴陳伏生卜文圣
林業科學 2020年11期
陳奕帆 付曉莉 王輝民 戴曉琴 寇 亮 陳伏生 卜文圣
(1.中國科學院地理科學與資源研究所 生態系統網絡觀測與模擬重點實驗室千煙洲生態站 北京 100101; 2.中國科學院大學資源與環境學院 北京 100049; 3.江西農業大學林學院 南昌 330045)
杉木(Cunninghamialanceolata)是我國南方最重要的常綠針葉樹種之一,具有3 000多年的培育歷史,在我國亞熱帶地區廣泛分布,具有較好的耐用性和較高的單位產量,是重要的用材林樹種(Shietal., 2010)。過去20~30年,杉木人工林多以高密度、短輪伐期經營為主,經營重點集中在中小徑材上。隨著人們對多樣化木材材種需求的升高,大徑材供需矛盾凸顯(葉功富等, 2006)。因此,研究杉木中齡林的科學經營方式對充分發揮中齡杉木的生長潛力、培育杉木大徑材具有重要意義。
林下植被清除是人工林經營管理中的一項重要措施(Lebeletal., 2008),其目的是為了提高木材質量和產量(Stokesetal., 2014)。林下植被清除主要用于幼林撫育階段,因為在幼林階段,林下植被與喬木對水分、養分的競爭尤為激烈,清除林下植被對喬木生長最有利(Stokesetal., 2014)。隨著喬木的生長,高大喬木本身會阻礙林下植被發展(Busseetal., 1996),清除林下植被對喬木生長的促進作用可能會減弱(Busseetal., 1996; Zhangetal., 2013; Stokesetal., 2014)。但已有研究多關注林下植被清除對林分水平喬木生長的影響(Busseetal., 1996; Takahashietal., 2003; Takahashietal., 2006; Zhangetal., 2013; Stokesetal., 2014),而同一林分中相同林齡不同徑級喬木對同一管理措施的響應機制仍不明確。
影響植物個體生長的機制有化學內穩性、資源利用效率、碳分配模式等(Evans, 1989; Jacobetal., 1991; Lietal., 2009; Garrishetal., 2010; Xieetal., 2018; 任書杰等, 2012)。氮(N)、磷(P)是保持植物結構、維持植物生理代謝功能的重要元素,是生產力形成的重要限制因素(Jacobetal., 1991; Lietal., 2009; 任書杰等, 2012)。N、P元素可通過影響CO2同化速率和水分利用效率(WUE)、光合氮素利用率(PNUE),進而影響植物生長(Evans, 1989; Garrishetal., 2010; 李勇, 2011)。N/P既可用來判斷林木生長過程中土壤N的供應狀態(Herbertetal., 2003),還能反映植物個體、群落及生態系統的養分限制情況(Gusewell, 2004; Zhangetal., 2004)。此外,碳水化合物是樹木細胞結構的主要組成成分,也是植物生長代謝過程中的重要能量來源(鄭云普等, 2014)。碳水化合物在植物體內的存在形式主要包括結構性碳水化合物和非結構性碳水化合物(NSC)。其中,結構性碳水化合物(木質素、纖維素等)作為植物細胞組成成分主要用于植物生長,而NSC是植物生命代謝的重要碳庫,為植物各項生理功能的正常發揮提供物質基礎(潘慶民等, 2002)。在植物生長過程中,大部分光合產物不會直接用于新組織形成和代謝過程,而是以NSC的形式儲存在植物體內,在植物需要時被消耗以維持植物正常生長和代謝(Kozlowski, 1992)。NSC含量的大小能夠反映植物抵御外界逆境的能力(Xieetal., 2018)。在干旱脅迫時,氣孔的關閉減少碳固定,使樹木依賴儲存的NSC進行代謝活動來抵抗逆境和維持滲透壓(Mcdowell, 2011)。
本研究以中齡杉木人工林為對象,依托林下植被管理實驗平臺,研究不同徑級中齡杉木化學計量比、碳的光合同化速率、水和氮的利用效率及碳分配模式(如結構性碳與NSC間的分配,NSC在不同器官間的分配),探討不同徑級中齡杉木生長速率對林下植被管理措施的響應機制,為中齡杉木的科學管理策略提供理論支撐。
1 研究區概況
樣地位于江西省泰和縣中國科學院千煙洲石溪林場(115°03′29.9″E,26°44′29.1″N)。氣候屬于典型的亞熱帶大陸性季風氣候,年均氣溫約17.1 ℃,年降水量1 377.4 mm(Yangetal., 2015)。研究區在7-10月存在著明顯的季節性干旱(Yangetal., 2017)。土壤為典型低鹽基濕潤弱育土,土壤母質為紅色砂巖和泥石風化物(Kouetal., 2015),pH值為4.87,有機碳、總氮、可溶性有機碳和有效氮含量在自然條件下約為19.1 g·kg-1、1.4 g·kg-1、32.9 mg·kg-1和15.3 mg·kg-1(Yangetal., 2019)。樣地坡度約15°,土壤厚度50~100 cm。
2 研究方法
2.1 樣地設置
本試驗依托2013年建立的杉木人工林林下植被管理實驗平臺,杉木林于1998年種植。2013年的本底調查顯示: 林分平均樹高10.2 m,胸徑9.8 cm,郁閉度0.75,林下植被以芒萁(Dicranopterisdichotoma)、鉤藤(Uncariarhynchophylla)、南燭(Vacciniumbracteatum)、鹽膚木(Rhuschinensis)、紅花檵木(Loropetalumchinense)、短柄枹(Quercusserrata)、大青(Clerodendrumcyrtophyllum)、杜鵑花(Rhododendronsimsii)為主(Yangetal., 2019),林下植被約為6 236 kg·hm-2。本試驗包括2個處理: 保留林下植被(CK),清除林下植被(UR,刈割并移出樣地)。每個處理有6塊樣地重復,樣地大小15 m×30 m,樣地隨機排列。2013年1月移除UR樣地林下植被,每個月對樣地進行維護,清除UR樣地中萌生的草本及灌木。至2016年,UR樣地林下植被僅有少量草本及零星灌木。2016年,林分密度為2 143棵·hm-2,平均樹高為13.7 m,胸徑14.7 cm。2016年,CK處理林木胸徑分布: 11~12 cm(4.3%)、12~13 cm(8.5%)、13~14 cm(15.0%)、14~15 cm(25.5%)、 15~16 cm(15.0%)、16~17 cm(10.6%)、17~18 cm(8.5%)、18~19 cm(12.8%); UR處理林木胸徑分布: 11~12 cm(16.7%)、12~13 cm(27.1%)、13~14 cm(14.6%)、14~15 cm(0)、15~16 cm(2.1%)、16~17 cm(22.9%)、17~18 cm(12.5%)、18~19 cm(4.2%)。CK處理中胸徑為11~12 cm的林木少; UR處理沒有胸徑為14~15 cm的林木,且15~16 cm及18~19 cm兩個范圍內的林木少。為確保不同處理間具可比性,即相同徑級CK和UR處理都有多個樣樹重復,所以重點關注12~14 cm和16~18 cm兩個徑級。其中,12~14 cm和16~18 cm兩徑級杉木占CK處理總林木數的42.6%,占UR處理總林木數的77.1%。
2.2 樣品采集與分析
基于樣地內林木徑級分布調查,每個樣地選8棵涵蓋整個徑級分布范圍的樣樹,12個樣地共計96棵樣樹。于2016年8月、2018年8月用胸徑尺測定每棵樣樹的胸徑,分別記錄為D2016、D2018。每棵樣樹胸徑處均有固定樹牌以確保數據的連續性和可比性。相對生長速率(RGR)的計算方法: RGR=0.5×(D2018-D2016)/D2016。
于2016年8月(旱季)使用Li-6400便攜式光合儀(Li-Cor,Lincoln,Nebraska,USA)測定葉片光合參數。在每棵樣樹的樹冠中下向陽部位選取1個樹枝,選擇健康且成熟的葉子進行光合參數的測定,以獲取每顆樣樹葉片最大瞬時光合速率(Amax)、蒸騰速率(Tr),并計算WUE(WUE=Amax/Tr)。然后,將樹枝、葉片分別按照新(1年)與老(2年、3年)分開,測定各葉齡對應的葉片面積。將枝條和葉片樣品烘干,估算葉片比葉面積(SLA)。用生長錐在每棵樣樹胸高處的樹干陰陽兩面取約4 cm深的樣品,去樹皮后混合成1個樹干木質部樣品,冷藏殺青后65 ℃烘干。將所有烘干后的植物樣品用球磨儀粉碎。
采用酶解法測定各樣品中的NSC(包括可溶性糖(SS)和淀粉)(Kozlowski, 1992)。為確定林木碳水化合物是優先用于存儲還是生長,計算每棵樹的NSCtrunk/RGR和NSCleaf/RGR(trunk: 樹干,leaf: 樹葉)。其中,為確定光合固定的碳在存儲與生長間分配的相對優先性,對NSC和RGR的數值均相對各自最大值進行了歸一化(Saffelletal., 2014)。NSCtrunk/RGR和NSCleaf/RGR值越大表示光合固定的碳越優先用于存儲,而非生長(Saffelletal., 2014)。
葉片全氮采用元素分析儀(德國elementar公司)測定,全磷采用電感耦合等離子體光譜儀(ICP-OES 5300DV,PerkinElmer)測定。用SLA將葉片N質量百分比含量換算成單位葉面積N含量(Narea),然后計算PNUE(PNUE=Amax/Narea)。
2.3 數據處理
利用SPSS18.0,采用雙因素方差分析法分析徑級和林下植被處理對RGR、Amax、WUE、PNUE、NSC及其組分的影響,采用t檢驗分析同一徑級不同處理間參數的差異性,顯著水平為P<0.05。數據在分析前均進行方差同質性檢驗,以滿足正態分布。采用多元逐步回歸法分析不同因素(化學計量、資源利用效率、碳分配)對于相對生長速率的影響。利用SigmaPlot12.5軟件進行繪圖。
3 結果與分析
3.1 林下植被清除對中齡杉木生長的影響
林分水平上,杉木RGR在UR處理為2.54%,CK處理為3.24%,即清除林下植被顯著降低了中齡杉木相對生長速率(P=0.035)。清除林下植被對中齡杉木生長的影響因徑級不同而異(圖1A)。為確保每個徑級內均有樣樹重復,分析中主要關注林下植被管理對12~14 cm和16~18 cm兩個徑級的中齡杉木生理生態屬性的影響。清除林下植被對12~14 cm徑級的中齡杉木相對生長速率無顯著影響,但會減緩16~18 cm徑級的中齡杉木生長(圖1B)。
3.2 中齡杉木光合參數和葉片養分的徑級差異
清除林下植被對12~14 cm和16~18 cm徑級的中齡杉木Amax、WUE、PNUE均無顯著影響(表1),這表明清除林下植被沒有改變中齡杉木的葉片光合固碳能力、葉片氮素光合效率和水分利用效率。對兩徑級的中齡杉木葉片全N、全P含量和N/P也無顯著影響(表2),表明清除林下植被不會影響中齡杉木的養分限制情況。但兩徑級的中齡杉木葉片全N、N/P差異顯著。16~18 cm、12~14 cm徑級的中齡杉木葉片N/P分別為14.00和12.89。
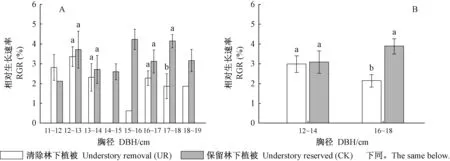
圖1 林下植被清除和徑級對中齡杉木相對生長速率的影響Fig.1 Effects of understory removal and DBH classes on relative growth rate of middle-aged Chinese fir不同小寫字母表示在某一徑級不同處理間差異顯著(P<0.05)。下同。Different lowercase letters indicated significant differences among treatments in a given DBH class (P<0.05).The same below.
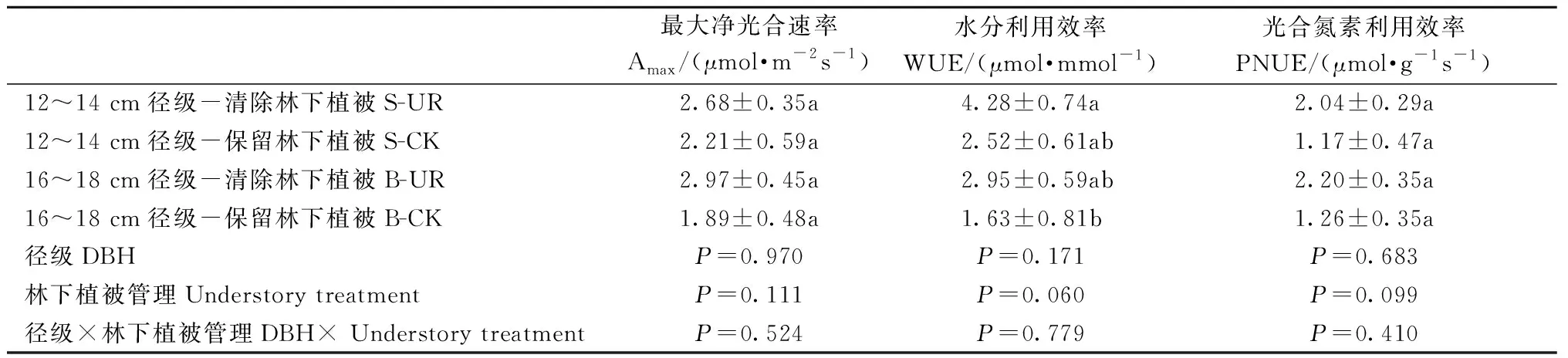
表1 徑級和林下植被清除對中齡杉木光合參數的影響①Tab.1 Effects of DBH class and understory removal on photosynthetic parameters of middle-aged Chinese fir
3.3 中齡杉木非結構性碳水化合物分配的徑級差異
清除林下植被對12~14 cm和16~18 cm兩徑級NSCleaf、NSCbranch和NSCtrunk均無顯著影響(表3),但對16~18 cm徑級中齡杉木NSC在不同年齡葉片中的分配影響顯著(圖2,3)。對于16~18 cm徑級的中齡杉木而言,清除林下植被顯著降低NSCnl(圖2A)、NSCnl/NSCol(圖3A),SSnl(圖2B)、SSnl/SSol(圖3B)(nl: 新葉,ol: 老葉),而對新葉淀粉無顯著影響(P=0.667)。以上結果表明, 清除林下植被主要是通過降低16~18 cm徑級的中齡杉木新葉可溶性糖的含量及分配比例,進而降低NSC向新葉的分配。對于12~14 cm徑級中齡杉木,清除林下植被顯著增加了SSnl/SSol(圖3B),而對NSCnl(圖2A)、SSnl(圖2B)、NSCnl/NSCol無顯著影響(圖3A)。以上結果表明: 盡管清除林下植被有促進該徑級中齡杉木可溶性糖向新葉分配的趨勢,但并未造成新葉SS和NSC含量發生顯著變化。
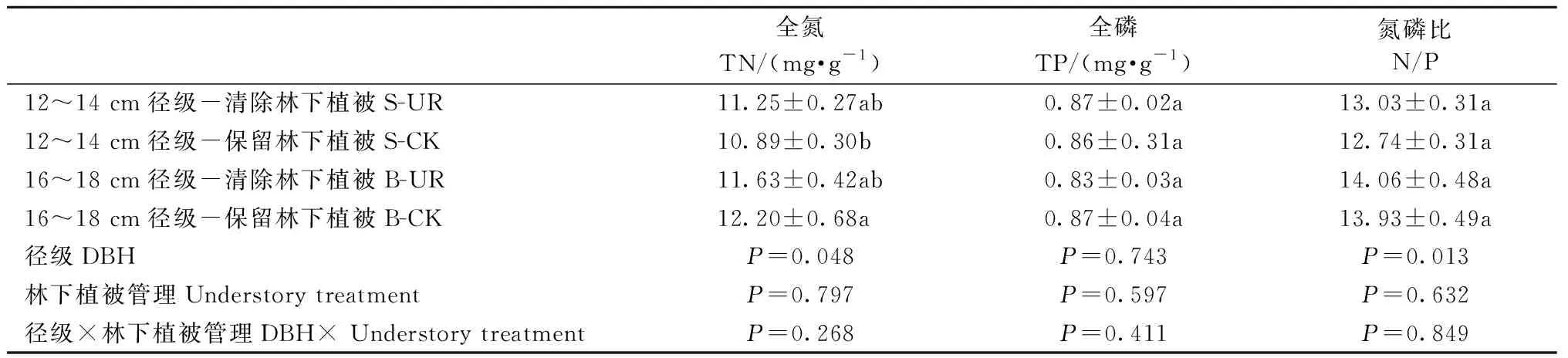
表2 徑級和林下植被清除對中齡杉木葉片養分含量的影響Tab.2 Effects of DBH class and understory removal on leaf nutrient of middle-aged Chinese fir

表3 徑級和林下植被清除對中齡杉木葉、枝、干非結構性碳水化合物的影響Tab.3 Effects of DBH class and understory removal on NSCs of leaf, branch,trunk of middle-aged Chinese fir
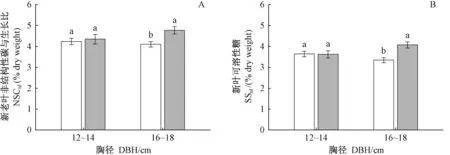
圖2 林下植被清除和徑級對中齡杉木新葉NSC和可溶性糖的影響Fig.2 Effects of understory removal and DBH classes on NSC and SS in new foliage of middle-aged Chinese fir下角的nl、ol分別為新葉、老葉。The inferior figures nl and ol means newleaf and old leaf respectively.
對于16~18 cm徑級的中齡杉木,清除林下植被對NSCtrunk和NSCleaf無顯著影響(表3),但減小了其相對生長速率(圖1B),導致NSCtrunk/RGR和NSCleaf/RGR比值增大(圖4A,4B)。以上結果表明, 清除林下植被后,16~18 cm徑級的中齡杉木光合固定的碳優先用于存儲,而非生長。清除林下植被對12~14 cm徑級的中齡杉木NSCtrunk/RGR和NSCleaf/RGR無顯著影響(圖4A,4B),說明清除林下植被沒有改變12~14 cm徑級的中齡杉木光合固定的碳在存儲與生長間的分配模式。
3.4 影響中齡杉木生長速率的主控因子
多元逐步回歸結果表明: 光合固定的碳在存儲與生長間的分配模式是影響中齡杉木生長的主要因素(表4)。NSCleaf/RGR是影響12~14 cm徑級的中齡杉木生長速率的主要驅動因子,NSCleaf/RGR越大越不利于杉木生長。對于16~18 cm徑級的中齡杉木,NSCleaf/RGR與SSnl共同調控杉木生長速率,NSCleaf/RGR與RGR呈負相關,SSnl與RGR呈正相關,且NSCleaf/RGR對生長速率的解釋度高于SSnl。
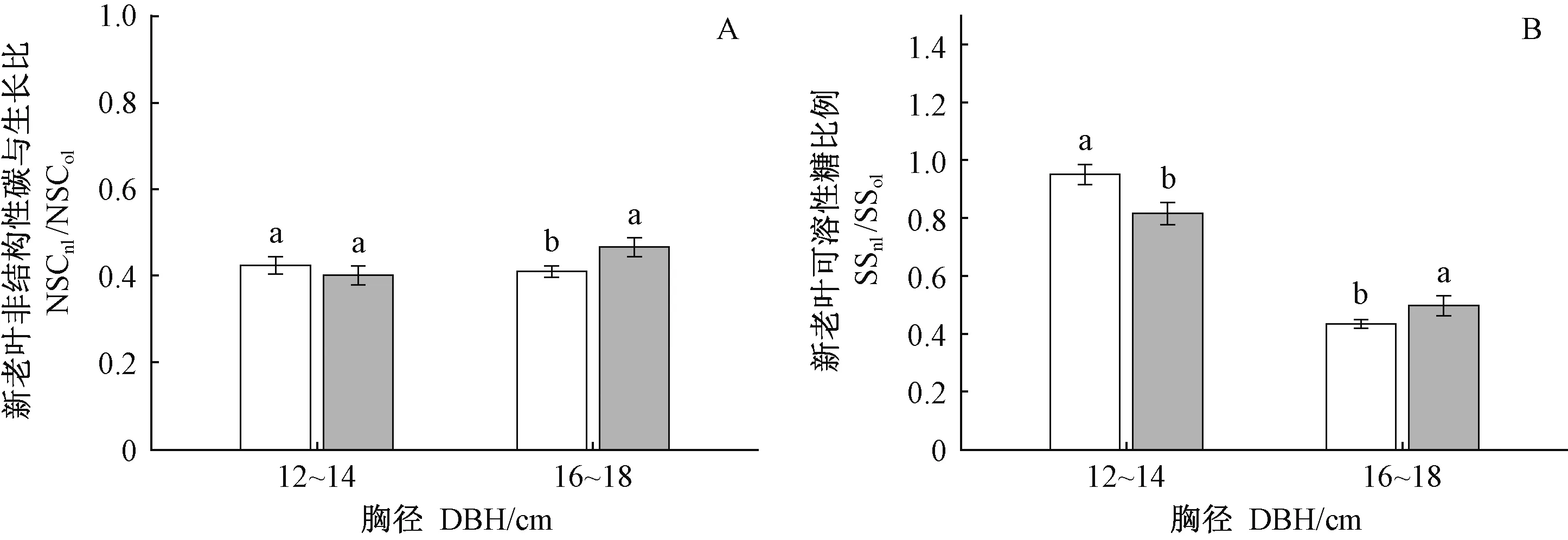
圖3 林下植被清除和徑級對中齡杉木葉片NSC和可溶性糖在新、老葉間分配的影響Fig.3 Effects of understory removal and DBH classes on allocations of NSC and soluble sugar between the leaves of middle-aged Chinese fir
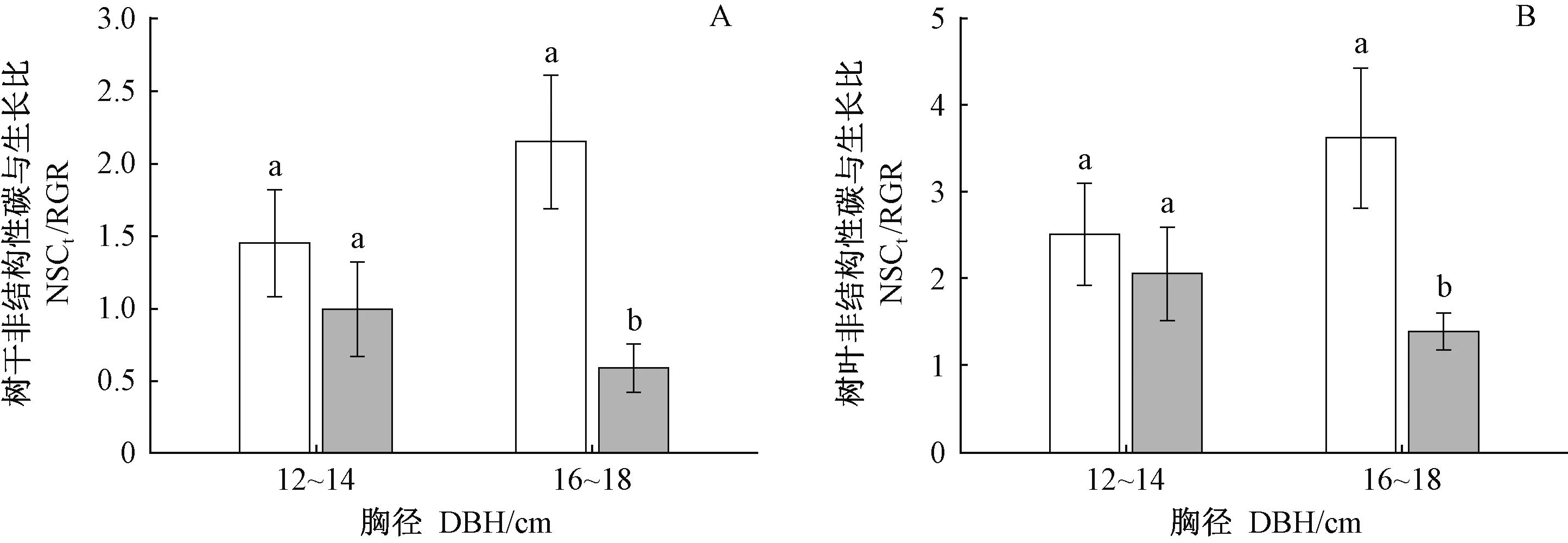
圖4 林下植被清除和徑級對中齡杉木碳存儲與生長關系的影響Fig.4 Effects of understory removal and DBH classes on the relationships between carbon storage and growth of middle-aged Chinese fir
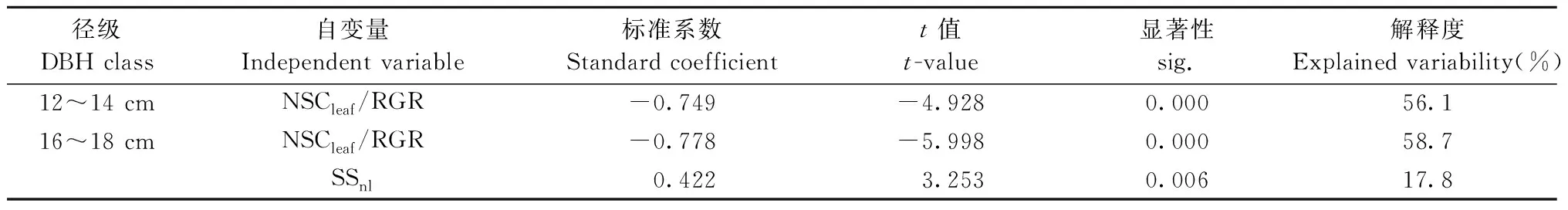
表4 中齡杉木相對生長速率多元逐步回歸結果Tab.4 Results of stepwise regression analysis for relative growth rate of middle-aged Chinese fir
4 討論
4.1 林下植被清除對林分水平喬木生長的影響
當林下植被與喬木競爭激烈土壤水分、養分時,清除林下植被一般可促進保留木生長(Takahashietal., 2003; Takahashietal., 2006; Lebeletal., 2008; Zhangetal., 2013; Stokesetal., 2014)。但本研究結果與此相反,Wan等(2014)研究4年生桉樹(Eucalyptusrobusta)林中清除林下植被減慢了喬木生長(Wanetal., 2014)與本研究相似。這說明清除林下植被對喬木生長的影響存在不確定性,可能是因林下植被與喬木間的競爭會隨喬木生長而相對減弱(Busseetal., 1996; Zhangetal., 2013),或使二者關系主要表現為協同關系。
首先,林下植被覆蓋能有效減少土壤侵蝕(袁正科等, 2000),林下植被也可將滲入土壤深層的養分運移到表層土壤以供其他植物利用(Jobbágyetal., 2004)。其次,林下植被可緩解土壤酸化、優化土壤微生物群落結構、提高真菌比例(Fuetal., 2015)。此外,土壤碳分布與林下植被細根密切相關,林下植被細根多的土壤斑塊的碳含量高(Liaoetal., 2014),且土壤表層中林下植被吸收根的量高于喬木(Jiangetal., 2018),林下植被吸收根分泌物能提高土壤微生物活性、加快凋落物分解(鄭琳琳等, 2017),因此林下植被細根對促進養分釋放有重要意義。清除林下植被會減少根系分泌物,導致β-1,4-葡糖苷酶活性及其最大反應速率降低,碳獲取酶活性下降(Yangetal., 2018; Yangetal., 2019)。因此,清除杉木人工林林下植被可顯著降低土壤全N、全P及可溶性C含量、減小土壤真菌含量、減緩土壤養分循環速率(Wuetal., 2011; Liuetal., 2012; Zhaoetal., 2012)、使植物光合固定的碳優先用于地下根系生長而非地上生長(Reichetal., 2014)。
清除林下植被會使太陽輻射能、蒸散發量、地表空氣流動等環境因子進行系統內重新分配(吳彥等, 2004; 臧潤國, 2016),土壤溫度及水分分布特征發生變化(Hoggetal., 1991; Takahashietal., 2003; Matsushimaetal., 2007; Wuetal., 2011),間接影響表層土壤有機質的分解及氮礦化過程。研究發現, 清除林下植被可使10 cm處土壤溫度升高(Hoggetal., 1991; Matsushimaetal., 2007; Wuetal., 2011)、提高土壤微生物活性和凋落物分解速率、加快N的礦化與硝化、提高N有效性、促進喬木生長(杜忠等, 2016)。而同在本試驗樣地開展的研究發現,林下植被清除后土壤溫度、水分均無顯著變化(楊洋等, 2016)。因此,土壤溫度、水分不是本研究中喬木生長減緩的原因。
4.2 林下植被清除對林木個體化學計量、資源獲取及利用效率的影響
不同林下植被物種的根際效應不同(Daietal., 2018; 莫雪麗等, 2018),且不同喬木個體周圍的林下植被物種可能不同。因此,清除林下植被對不同喬木個體周圍土壤養分釋放的影響可能會有差異。但本研究中的林木樣樹均是隨機取樣,可排除這個潛在影響因素。
氮和磷在植物的生理功能中起著至關重要的作用,是陸地生態系統中限制植物生長的最重要的元素(Chapin, 1980; Reichetal., 1997)。葉片N/P可表征植物養分受限情況。通常,葉片N/P>16表示植物受P限制; 葉片N/P<14表示植物受N限制; 葉片N/P處于14和16之間表示N、P均不受限制或者均受限制(Koerselmanetal., 1996)。本研究中12~14 cm徑級的中齡杉木受N限制,而16~18 cm徑級的中齡杉木N、P化學計量平衡較好。但林下植被清除并未影響兩徑級的中齡杉木的養分狀況(表2)。這可能是因為清除林下植被一方面減少了單位土體養分釋放的總量,另一方面排除了林下植被對土壤養分消耗這一分量(即清除林下植被后土體養分僅供喬木所用)。
在一個植物群落中,長勢好的植物個體通常會有較高的資源獲取能力或資源利用效率,如碳同化速率較大、水分利用效率或氮利用效率高等(Funketal., 2007; 劉鵬等, 2019)。盡管清除林下植被可增大喬木冠層葉片的最大凈光合速率(Kitaoetal., 2003; Kobayashietal., 2006; Sirdah, 2017),減小林分水分利用效率(張浩, 2017)。但本研究中林下植被清除未改變林木的資源吸收和利用效率(表1)。
4.3 林下植被清除對林木個體碳分配的影響
碳水化合物主要以可溶性糖的形式在韌皮部運輸,參與新陳代謝,為樹木生長提供重要能源(Chapin, 1990)。樹木遇到干旱等逆境脅迫時,用可溶性糖調節滲透壓,以維持樹木細胞的正常活性(Salaetal., 2012)。新葉的CO2光合能力、凈同化量與凈光合強度高于老葉,對林木的碳固定及生長意義重大(張小全等, 2000; 謝國陽, 2002; 許忠坤等, 2008)。清除林下植被后,16~18 cm徑級樹木新葉的NSC和可溶性糖含量均下降,可導致干旱季節杉木葉片滲透壓調節能力下降,進而影響杉木生長。其次,林下植被清除后,16~18 cm徑級的中齡杉木相對提高了樹干、葉片NSC存儲量,進而相對地減少了用于生長的結構性碳的分配(圖4)。即當光合吸收的碳不變時,16~18 cm徑級中齡杉木光合同化的碳優先用于存儲,而非生長。綜上可知,對于16~18 cm徑級的中齡杉木而言,清除林下植被后,葉片(尤其是新葉)NSC及可溶性糖的降低及光合固定的碳優先用于存儲而非生長共同導致了其生長緩慢(表4)。
對于12~14 cm徑級的中齡杉木,葉片SSnl/SSol顯著增加(圖3)。這可能是由于該徑級的中齡杉木受N限制顯著,而植物面對資源不足時將固定的碳優先向獲取資源能力強的器官或組織中分配(Chapinetal., 1987)。然而,限制12~14 cm徑級的中齡杉木生長的主要因子是NSCleaf/RGR(碳存儲與生長間的關系),而非葉片NSC(或可溶性糖)在新、老葉之間的分配比例(表4),且林下植被清除對該徑級中齡杉木NSCleaf/RGR無顯著影響。因此,林下植被清除沒有顯著改變12~14 cm徑級的中齡杉木的生長速率。
4.4 本研究的局限與不足
本研究是用2016年杉木生理生態指標來解釋其后續2年的生長,這存在一定的不確定性,因為隨著林下植被管理措施的年限延長,林木生長的響應規律可能會發生變化。因此,林木生長對林下植被管理措施的長期響應規律還需未來更多研究進行驗證。
此外,林業生產中通常是割倒林下植被后將殘余物歸還到林分中。林下植被割后歸還比林下植被正常生長更有利于增加土壤酶活性,因為產生的林下植被地上、地下植物殘體比活根和正常凋落物輸入提供的碳源更多(楊洋等, 2016)。而本研究是持續地將林下植被割倒并將產生的殘余物移出到林分外,這可能導致水土流失、不利于土壤水分和微生物群落結構等土壤微環境的維持(Zhaoetal., 2013)。因此,將林下植被殘余物移出到林分外也可能導致林下植被去除未促進林木生長。未來研究需增加割倒林下植被但不移出的處理,并設計不同的林下植被刈割強度梯度,進一步探討林下植被管理對林木生長的影響機制。
5 結論
基于本研究的觀測數據,表明清除并移除林下植被會減緩中齡杉木的生長速率。但不同徑級的中齡杉木生長速率對林下植被管理措施的響應不同,即林下植被清除會降低16~18 cm徑級的中齡杉木的相對生長速率,而對12~14 cm徑級的中齡杉木相對生長速率無顯著影響。林下植被管理對中齡杉木生長影響的徑級依賴性主要受碳分配(不同形態間,相同形態不同器官間)機制調控,而非化學計量、資源獲取及利用效率等機制影響。清除林下植被導致16~18 cm徑級的中齡杉木新葉NSC(尤其是可溶性糖)含量降低,且使林木光合產物優先用于存儲而非生長,進而減緩了該徑級的林木生長速率。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
