成人與新生兒來源的間充質干細胞circRNAs 差異表達譜分析及功能預測
2021-01-04 07:03:02梁甲武袁一方
解放軍醫學院學報 2020年10期
關鍵詞:分析
梁甲武,胡 巍,屈 爽,袁一方,郭 斌
解放軍醫學院/ 解放軍總醫院第一醫學中心 口腔科,北京 100853
間充質干細胞(mesenchymal stem cells,MSCs)具有多向分化和自我更新能力,其促進組織動態平衡和損傷修復方面的作用越來越受到重視[1]。然而,近年來研究發現,隨著年齡增長,MSCs 的再生能力下降,細胞功能出現障礙[2]。增齡對于MSCs 細胞命運的具體調控機制尚不清楚。通過探索增齡調控MSCs 細胞命運的具體機制,從而實現延緩甚至逆轉MSCs 衰老,成為未來的發展方向。環狀RNA(circular RNAs,circRNAs) 是一類反向剪接形成的閉合環狀結構的非編碼小RNA,與微小RNA(microRNAs,miRNAs) 內源性競爭性結合是circRNAs 的主要生物學功能之一。CircRNAs通過競爭性結合miRNAs,解除miRNAs 對靶基因的抑制作用,進而促進靶基因表達。目前研究發現,circRNAs 在組織器官發育和疾病發展等多個領域均發揮調控作用[3-4]。但circRNAs 在干細胞生物學領域的研究相對較少,對MSCs 的細胞命運影響仍待深入探索。本研究通過運用多種生物信息學方法,對成人與胎兒來源MSCs 的circRNAs差異表達譜進行分析, 并通過基因本體(Gene Ontology) 注釋和 KEGG 通路 (KEGG pathway) 分析對circRNAs 可能發揮作用的信號通路進行預測,探究circRNAs 在增齡調控MSCs 細胞命運中的作用,為延緩甚至逆轉MSCs 衰老提供新的理論依據。
材料與方法
1 circRNA 微陣列數據及數據分析 本研究中使用的circRNA 微陣列數據GSE122178 從Gene Expression Omnibus(GEO) 數 據 庫 (www.ncbi.nlm.nih.gov/geo) 下載,獲得了來源于成人與新生兒MSCs 的 circRNAs 表達譜。GSE122178 數據集包括3 例成人骨髓來源的MSCs 樣本(GSM3457280、GSM3457281、GSM3457282)、3 例 圍 生 期 新生兒臍帶血來源的MSCs 樣本(GSM3457283、GSM3457284、GSM3457285)。采用 GEO2R 在線分析工具對上述兩組數據進行分析,并對差異表達circRNAs 進行篩選。
2 miRNAs 結合位點及下游靶基因預測 利用CircInteractome 在線工具預測circRNAs 的miRNAs結合位點, 并根據 Context+score percentile 評分對預測位點進行篩選。之后聯合應用miRDB 和Targetscan 數據庫對下游靶基因進行預測,并通過韋恩分析檢測出共表達的靶基因。
3 GO 注釋和KEGG 分析 GO 注釋將基因功能分為生物過程(BP)、細胞成分(CC) 和分子功能(MF)三個部分。應用DAVID 在線工具對共表達的靶基因進行GO 注釋和KEGG 分析。
4 統計學分析 計量數據以-x±s表示。以P<0.05 作為 mRNAs 差異表達、GO 注釋和 KEGG 分析的標準。以 |Fold change| > 2 且P< 0.05 作為circRNAs 顯著差異表達的標準。
結 果
1 成人與新生兒來源的MSCs 差異表達的circRNAs 基因芯片微陣列分析顯示,圍生期新生兒臍帶血來源的MSCs 與成人骨髓來源的MSCs 相比,存在4 140 個不同程度差異表達的circRNAs,其中66 個circRNA 表達顯著上調,107個circRNAs 表達顯著下調( 圖1)。在表達上調和下調變化中,最為顯著的分別是hsa_circ_0080170和hsa_circ_0060055。
2 hsa_circ_0080170 和 hsa_circ_0060055 下 游miRNAs 及靶基因預測 利用CircInteractome 對差異表達最顯著的hsa_circ_0080170 和hsa_circ_0060055 的miRNAs 競爭性結合位點進行預測。結果顯示,hsa_circ_0080170 具有 6 個 miRNAs 結合位點,hsa_circ_0060055 具有 28 個 miRNAs 結合位點。通過進一步參考Context+score percentile評分,最終各篩選出2 個miRNAs 進行下一步功能分析預測( 表1)。聯合運用miRDB 和Targetscan數據庫分別預測hsa-miR370、hsa-miR203a、hsamiR1225-5p 和hsa-miR197 可能調控的下游靶基因,并將2 個數據庫的預測結果進行韋恩分析。結果顯示,hsa-miR370 有 551 個靶基因,hsamiR230a 有 163 個靶基因,hsa-miR1225-5p 有 180個靶基因,hsa-miR197 有411 個靶基因( 圖2)。

表 1 hsa_circ_0080170 和 hsa_circ_0060055 的 miRNAs結合位點Tab. 1 miRNAs binding sites of hsa_circ_0080170 and hsa_circ_0060055
3 GO 注釋 和 KEGG 分 析 利 用 DAVID 對 hsamiR370 的 551 個靶基因和 hsa-miR230a 的 163 個靶基因進行GO 功能注釋,結果顯示靶基因在94個GO 項中富集(P<0.05),多涉及基因的轉錄調控和干細胞的分化及干性維持。hsa-miR1225-5p的180 個靶基因和hsa-miR197 的411 個靶基因的GO 注釋結果顯示,靶基因在58 個GO 項中富集(P<0.05),多與基因表達調控和組織再生相關。兩組GO 注釋結果中的前10 個重要富集項以生物過程、細胞組分和分子功能為分類在圖3 和圖4中顯示。基于GO 注釋的結果,對hsa-miR370 和hsa-miR203a 的714 個預測靶基因進行KEGG 通路分析,結果顯示所選靶基因與包括多能干細胞調控通路和Ras 信號通路在內的31 條信號通路相關(P<0.05)( 圖5)。此外,KEGG 通路分析發現與hsamiR1225-5p 和 hsa-miR197 的 591 個預測靶基因相關的17 條信號通路,包括肌動蛋白細胞骨架調控通路和泛素介導的蛋白水解等與干細胞命運調控相關的信號通路( 圖6)。
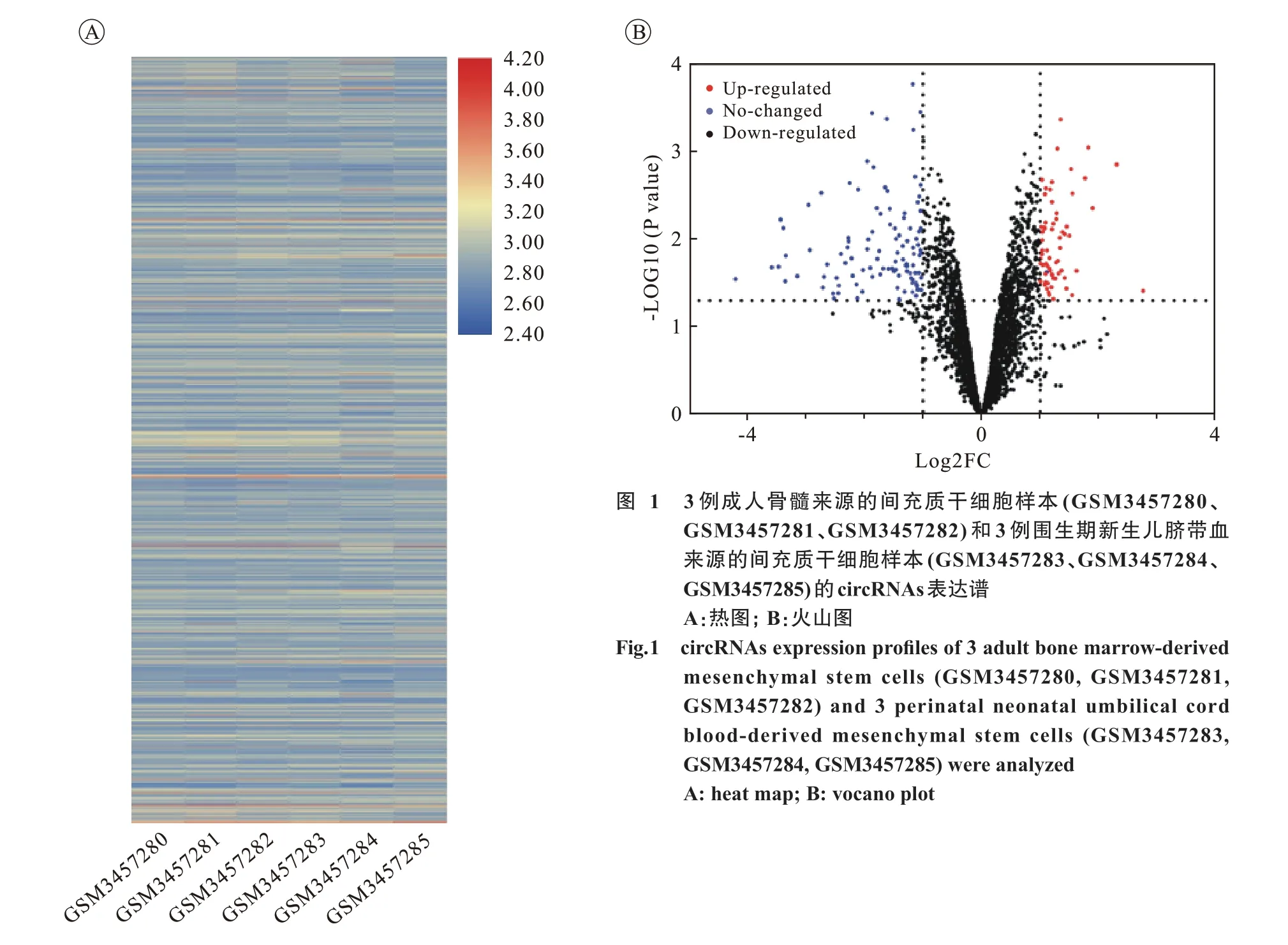
圖 1 3 例成人骨髓來源的間充質干細胞樣本(GSM3457280、GSM3457281、GSM3457282) 和3 例圍生期新生兒臍帶血來源的間充質干細胞樣本(GSM3457283、GSM3457284、GSM3457285)的circRNAs表達譜A:熱圖; B:火山圖Fig. 1 circRNAs expression profiles of 3 adult bone marrow-derived mesenchymal stem cells (GSM3457280, GSM3457281,GSM3457282) and 3 perinatal neonatal umbilical cord blood-derived mesenchymal stem cells (GSM3457283,GSM3457284, GSM3457285) were analyzed A: heat map; B: vocano plot

圖 2 hsa-miR230a、hsa-miR370、hsa-miR197和hsa-miR1225-5p各自的miRDB和Targetscan數據庫的下游靶基因預測結果及韋恩分析Fig. 2 Prediction results and Venn analysis of downstream target genes in the respective miRDB and Targetscan databases of hsa-miR230a,hsa-miR370, hsa-miR197 and hsa-miR1225-5p
討 論
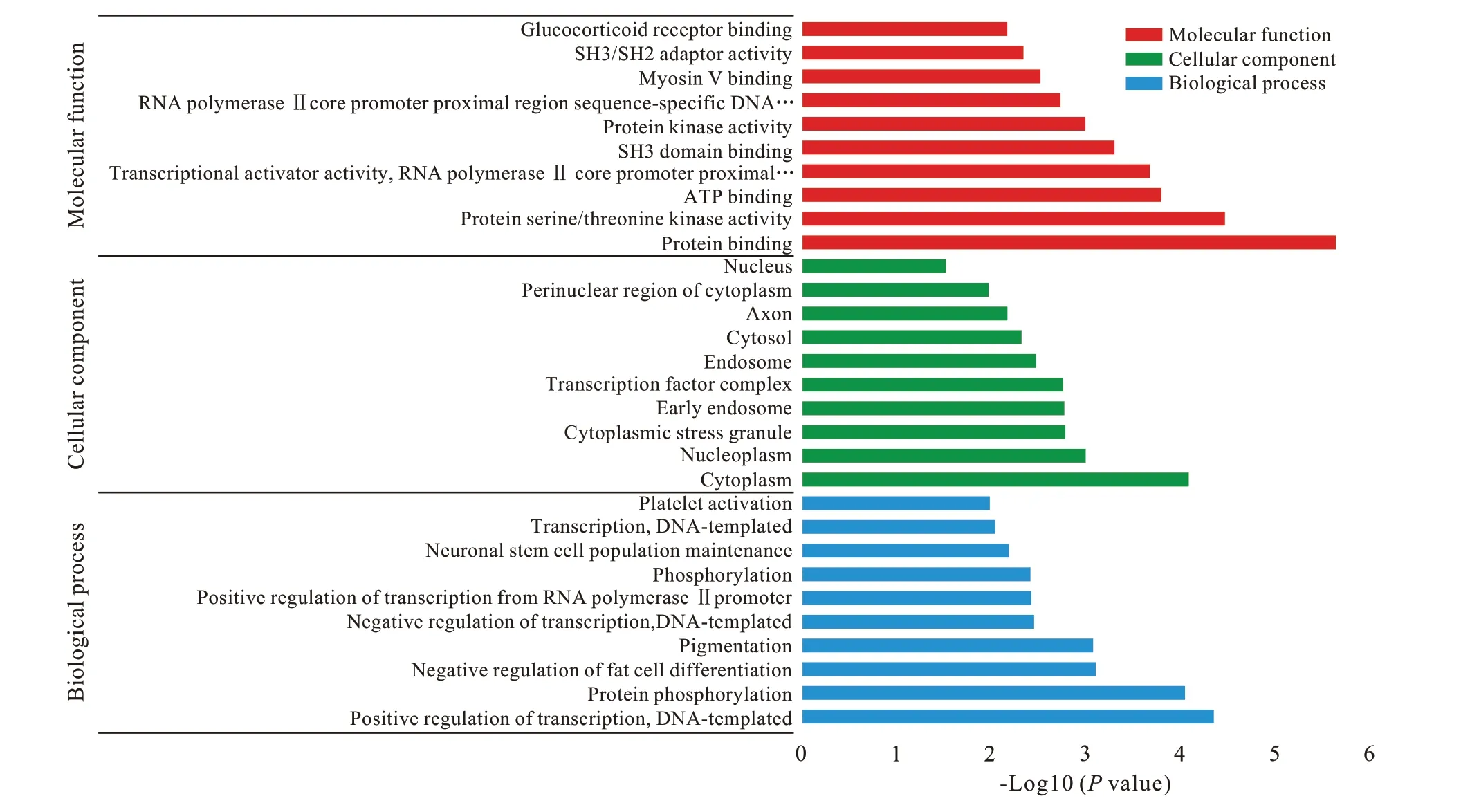
圖 3 hsa-miR370和hsa-miR203a的預測靶基因的前10項重要富集項以生物過程(BP)、細胞組分(CC)和分子功能(MF)為分類展示Fig. 3 The top 10 enriched gene ontology (GO) terms of the predicted target genes of hsa-miR370 and hsa-miR203a under the three main GO categories (BP: biological process; CC: cellular component; MF: molecular function)
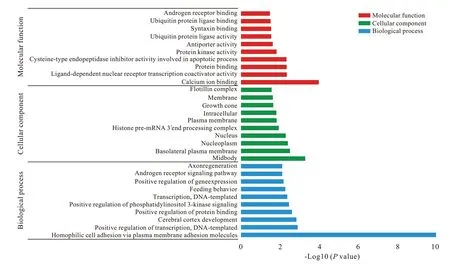
圖 4 hsa-miR1225-5p和hsa-miR197的預測靶基因的前10項重要富集項以生物過程(BP)、細胞組分(CC)和分子功能(MF)為分類展示Fig. 4 The top 10 enriched gene ontology (GO) terms of the predicted target genes of hsa-miR1225-5p and hsa-miR197 under the three main GO categories (BP: biological process; CC: cellular component; MF: molecular function)
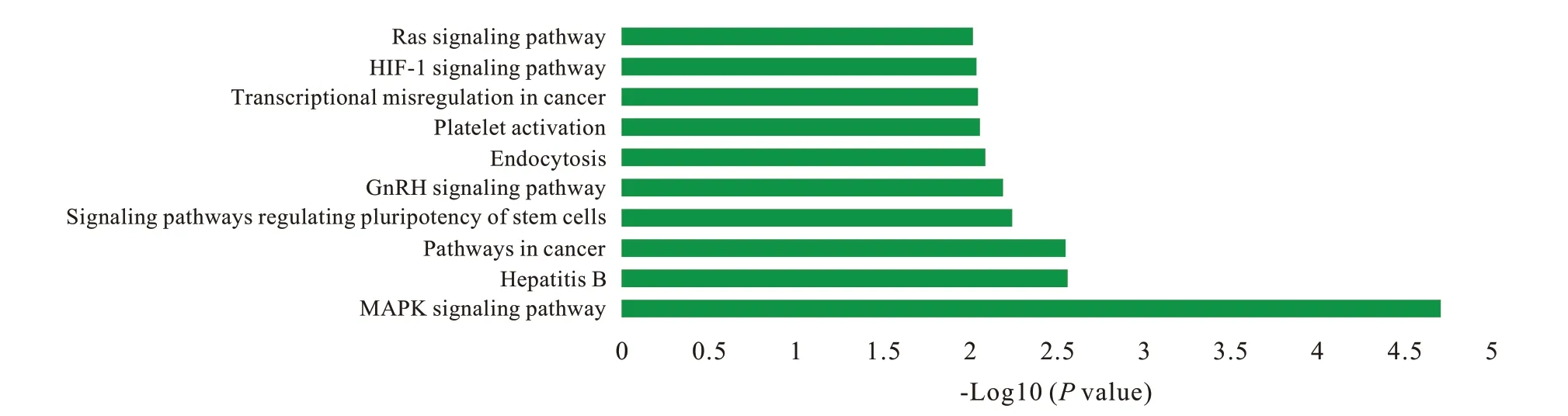
圖 5 hsa-miR370和hsa-miR203a的預測靶基因的前10條重要KEGG通路Fig. 5 The top 10 significant KEGG pathways of the predicted target genes of hsamiR370 and hsa-miR203a
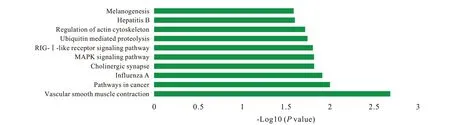
圖 6 hsa-miR1225-5p和hsa-miR197的預測靶基因的前10條重要KEGG通路Fig. 6 The top 10 significant KEGG pathways of the predicted target genes of hsamiR1225-5p and hsa-miR197
circRNAs 不具有5' 末端帽子和3' 末端poly(A)尾結構,以外顯子環化或內含子環化的方式成環。最初circRNAs 發現于植物類病毒中,并被認為是低豐度和技術限制造成的剪接錯誤的副產品[5]。直到近幾年,伴隨著高通量測序技術和生物信息學的快速發展,circRNAs 的結構和功能才逐步被發現。目前研究發現,circRNAs 由于特殊的環形結構,具有高度保守性、不易被核酸外切酶降解等特點[6-7]。同時,隨著對circRNAs 功能的深入研究,circRNAs 的海綿機制( 競爭性結合miRNAs)、與RNA 結合蛋白相互作用、參與蛋白質翻譯、調控基因轉錄等功能已陸續被發現。此外,研究人員已發現circRNAs 在口腔癌[8]、心血管疾病[9]、骨關節炎[10]等疾病發生發展過程中發揮作用,而且還會參與調控骨形成能力[11]、神經系統發育和衰老[3]等生理過程。近期,關于circRNAs 對間充質細胞影響的研究也逐漸增多。Wang 等[12]發現,CDR1as 可通過海綿吸附miR-7,激活ERK 信號通路,促進LPS 誘導炎癥條件下的牙周膜干細胞增殖。hsa_circ_0074834 可通過競爭性結合miRNA-942-5p 調控ZEB1 和VEGF 的表達,以及促進骨髓間充質干細胞的成骨分化,進而促進骨缺損修復,有望成為骨不連的關鍵治療靶點[13]。
近年來,已有多項研究表明,增齡至少會影響MSCs 的部分功能。Du 等[14]發現,源自18 ~ 20歲、30 ~ 35 歲、45 ~ 50 歲三個不同年齡段捐贈者的牙周膜干細胞之間,細胞集落形成率、細胞周期和凋亡情況以及堿性磷酸酶活性存在顯著差異。隨著年齡的增長,牙周膜干細胞的集落形成能力下降,G2/S 期細胞減少,細胞凋亡增加,成骨分化能力下降。此外,陳裕浩等[15]發現,與源于幼年雄性獼猴的骨髓間充質干細胞相比,源于老年雄性獼猴的骨髓間充質干細胞形態呈寬大、扁平形,細胞增殖緩慢,成骨、成軟骨及成脂分化能力下降, 伴隨著 IL-1β、IL-4、IL-6 和 TNF-α等細胞因子表達降低。然而,目前關于circRNAs在增齡對于MSCs 細胞命運的調控機制中發揮的作用少有研究。
本研究在GEO 數據庫檢索出建立于2019 年的關于circRNAs 在成人與新生兒來源MSCs 的circRNAs 差異表達數據集GSE122178,并對其中的3 例成人骨髓來源的MSCs 樣本和3 例圍生期新生兒臍帶血來源的MSCs 樣本進行GEO2R 在線分析,獲得顯著上調的hsa_circ_0080170 和顯著下調的 hsa_circ_0060055。 同時, 對 hsa_circ_0080170和hsa_circ_0060055 的下游miRNAs 及靶基因進行預測,初步形成circRNA-miRNA-mRNA 關系網絡。并且KEGG 分析發現,hsa-miR370 和hsa-miR203a作為hsa_circ_0080170 的下游miRNAs 結合位點,顯著富集到31 條信號通路;hsa-miR1225-5p 和hsa-miR197 作為hsa_circ_0060055 的下游miRNAs結合位點,顯著富集到17 條信號通路。其中多條信號通路與干細胞分化及干性維持相關。因此,根據生物信息學分析結果,預測hsa_circ_0080170和hsa_circ_0060055 可能在增齡調控MSCs 細胞命運中發揮作用,具體作用機制待進一步實驗驗證。本研究通過探究circRNAs 在增齡調控MSCs 細胞命運中的作用,為進一步探索延緩間充質細胞衰老的調控機制提供了新的理論依據。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
