基于益生菌調節的太平雞回腸SNP位點分析
2020-12-11 09:18:14雒林通馬芳唐德富
安徽農業科學 2020年21期
雒林通 馬芳 唐德富
摘要 為挖掘益生菌調節下太平雞回腸SNP位點,將120只太平雞隨機分為對照組與益生菌組,采用高通量測序技術對太平雞回腸轉錄組的SNP位點進行分析。結果顯示:SNP類型中,轉換類型明顯高于顛換類型,對照組中轉換類型占22.55%~27.45%,顛換類型占11.43%~13.87%;益生菌組與對照組結果相似,轉換類型(22.53%~27.46%)明顯高于顛換類型(11.37%~13.94%)。SNP位置分析表明,對照組和益生菌組總SNP位點分別為4 684 072和4 647 171個,其中位于內含子和基因間區的SNP位點最多。另外,在SNP功能分析中,同義突變SNP占比最高,在對照組和益生菌組中分別占7.01%和7.10%;非同義突變SNP在對照組和益生菌組中占比分別為2.97%和2.98%。該研究結果可為今后太平雞的分子標記開發、遺傳連鎖圖譜構建以及輔助育種等提供基礎數據。
關鍵詞 太平雞;益生菌;回腸;轉錄組測序;SNP
中圖分類號 S831 ?文獻標識碼 A
文章編號 0517-6611(2020)21-0086-05
Abstract In order to excavate SNP sites in ileum of Taiping Chicken regulated by probiotics,120 Taiping chickens were randomly divided into control group and probiotic group.The SNP sites in the transcriptome of Taiping chickens ileum were analyzed by high throughput sequencing technique.The results showed that the transition proportion was significantly higher than that of the transversion.The transition of control group accounted for 22.55%-27.45%,and the transversion accounted for 11.43%-13.87%.The results of probiotic group were similar to those of control group,and the transition proportion (22.53%-27.46%) was significantly higher than that of the transversion (11.37%-13.94%).The analysis of SNP locations showed that the total number of SNP sites in the control group and the probiotic group was 4 684 072 and 4 647 171 respectively,among which the most SNP sites were located in introns and intergenic.In the analysis of SNP function,the proportion of synonymous mutations of SNP was the highest,which was 7.01% in the control group and 7.10% in the probiotic group.Nonsynonymous mutations of SNP accounted for 2.97% in the control group and 2.98% in the probiotic group.This study could provide basic data for the development of molecular markers,the construction of genetic linkage map and the assistant breeding of Taiping Chicken in the future.
Key words Taiping Chicken;Probiotics;Ileum;Transcriptome sequencing;SNP
基金項目 甘肅省民生科技計劃項目(1503FCME005);天水市科技支撐計劃項目(2020-SHFZKJK-7879)。
作者簡介 雒林通(1973—),男,甘肅天水人,高級實驗師,碩士,從事動物遺傳育種與繁殖方面的研究。*通信作者,副教授,博士,從事動物遺傳育種方面的研究。
收稿日期 2020-06-02;修回日期 2020-06-23
太平雞具有較強適應性和豐富的基因庫,是我國優良家禽品種和甘肅三大優良地方雞種之一,主要分布于甘肅康縣太平鄉(現為陽壩鎮)因而得名[1]。太平雞以黑羽紅冠為主要特征,體格高大,體型緊湊,生產性能以肉用為主,肉蛋兼用,具有肉嫩骨細、皮脆味鮮、肉質特佳等特點,在市場上具有較高的價值[2]。近年來,由于應對雞群混養而導致太平雞雞種退化[3]、免疫性能低下等問題,部分飼料生產企業和養殖戶為了追求養雞業效益的最大化以及疾病防治中用藥的誤區,往往存在大劑量使用抗生素[3],使得藥物殘留、耐藥菌株大量出現、損傷消化免疫系統等問題頻發[4],對人與自然的可持續發展造成嚴重威脅,成為食品安全和綠色可循環畜牧業發展中最突出、最嚴峻的問題[5],所以積極尋找綠色飼料添加劑,對太平雞進行遺傳改良、選育新品種已成為提高養殖效益和健康化養殖水平的當務之急。
益生菌(probiotic)指的是在人和動物腸道內,通過調節動物腸道微生物區系平衡對宿主健康有益的活性微生物菌種[6]。研究表明,益生菌可以維持動物機體腸道菌群平衡,抵抗病原菌入侵,分泌抗菌物質及營養活性物質和改善腸道黏膜免疫功能[7-8]。益生菌進入機體腸道后,刺激宿主腸道分泌多種消化酶,通過充分降解宿主體內未被消化吸收的抗營養因子而提高飼料利用率,對動物生長發育和生產性能的提高具有促進作用。益生菌在宿主腸道內借助體內發酵作用產生的有機酸(乙酸、乳酸和丙酸)來有效降低腸道pH,這不利于有害菌的生長,且益生菌分泌并誘導產生抑菌蛋白對致病菌生長具有抑制作用[9]。此外,益生菌通過刺激宿主腸道分泌免疫抗菌物質,調節機體的免疫系統[10]。因此,益生菌制劑作為一種新型的微生物飼料添加劑,在其調節下能更好地促進養殖業的健康和穩定發展。
作為第三代分子標記的單核苷酸多態性(single nucleotide polymorphisms,SNPs)是由單個核苷酸的變異而導致基因組層面DNA序列的多態性,通常表現為單個堿基的轉換或顛換、缺失或插入,但實際上發生的只有轉換和顛換2種[11]。由于覆蓋密度大,具有遺傳穩定性和代表性,便于高通量自動化檢測等優點,已經廣泛應用在構建高密度遺傳連鎖圖譜構建、關聯分析、分子輔助育種、群體遺傳系統、品種鑒定等方面,并表現出良好的應用前景[12-14]。筆者以太平雞為研究對象,通過在飲水中添加益生菌制劑,通過高通量測序分析其對太平雞回腸多態性位點的干預作用,利用分子標記進行輔助選擇從而獲得更大的遺傳進展,以期為太平雞的遺傳與育種提供科學依據。
1 材料與方法
1.1 試驗材料
試驗用益生菌制劑由安徽諾偉康飼料有限公司生產;試驗動物為60日齡太平雞,由隴南市康縣梅園太平山雞養殖專業合作社提供。
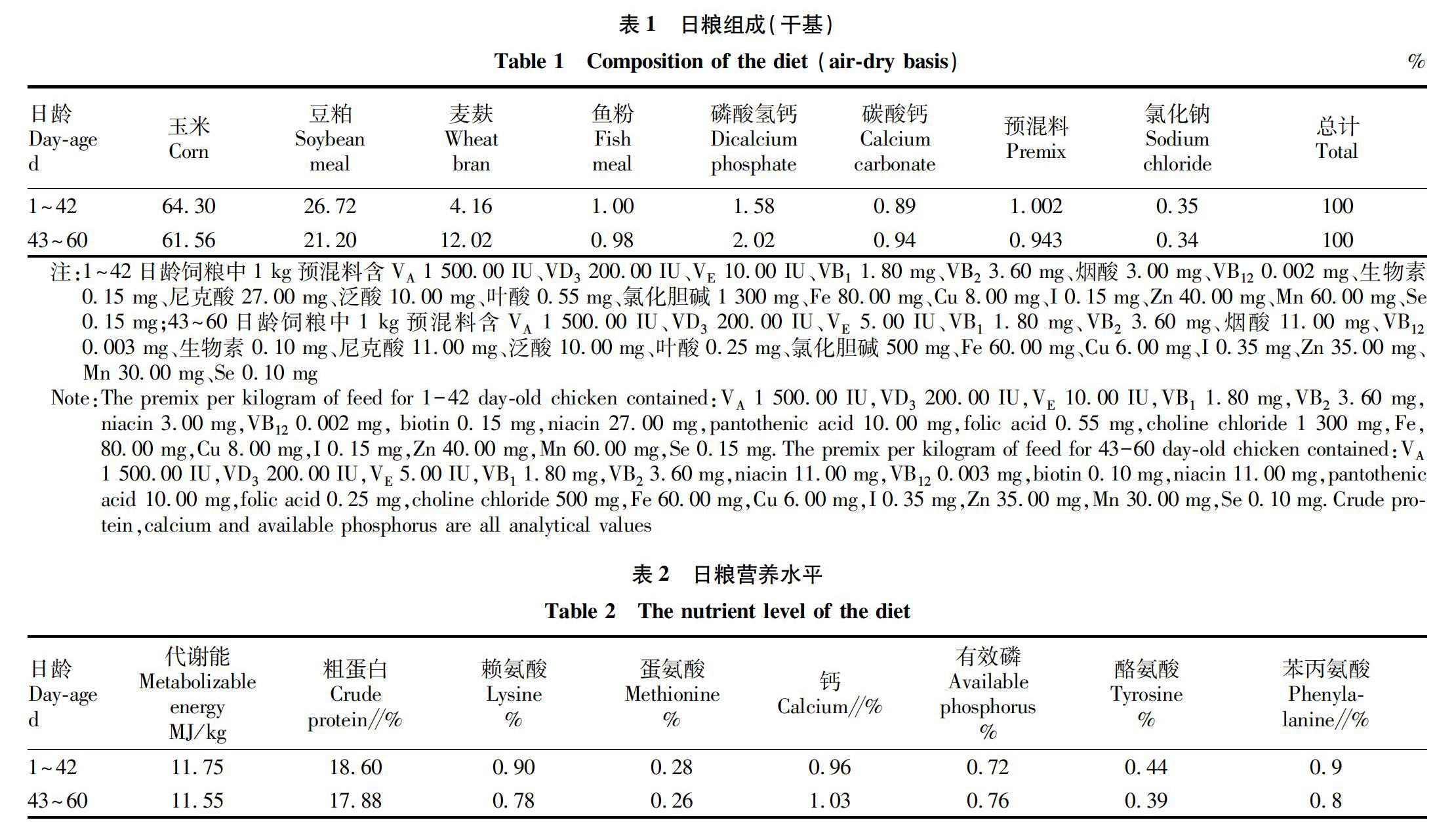
1.2 試驗設計與飼料 選擇0日齡太平雞120只,隨機分為2組,每組3個重復,每個重復20只。對照組(CT)飼喂基礎日糧,試驗組(MP)在飼喂基礎日糧的基礎上每升飲水中加入2 mL益生菌。飼料采購嚴格按照GB13078—2017《飼料衛生標準》和NY 5032—2006《無公害食品畜禽飼料和飼料添加劑使用準則》執行。參照《雞飼養標準》(NY/T 33—2004)配制玉米-豆粕型基礎飼糧,其組成及營養水平見表1~2。
1.3 飼養管理及樣品采集
第1~3天,溫度保持在33~35 ℃,此后每7 d降低2 ℃,直到最終室溫在18~21 ℃。光照時間1~3日齡為23~24 h,4~15日齡為23~16 h,16~60日齡為16~12 h;1~3日齡光照強度為10~30 lx,4~15日齡為5 lx,16~60日齡逐漸接近自然光照。室內相對濕度55%~70%,自由采食和飲水。
試驗用太平雞飼養至60日齡,各組每個重復取1只雞,即每組共3只雞。對照組(CT)樣品編號分別為CT1、CT2、CT3,試驗組(MP)樣品編號分別為MP1、MP2、MP3。稱重后心臟注射空氣致死,用新潔爾滅進行雞體消毒后,拔掉腹部羽毛,開腹后分離回腸,液氮中速凍,然后轉入-80 ℃低溫冰箱內保存。
1.4 RNA提取
用TRIzol試劑盒(Invitrogen,Carlsbad,CA,USA)分別提取MP1、MP2、MP3和CT1、CT2、CT3 6組回腸樣品的總RNA。RNA的降解和污染程度用1%瓊脂糖凝膠電泳檢測,RNA純度使用Nanodrop微量分光光度計檢測,RNA濃度和完整性用Qubit 2.0 Flurometer(Life Technologies,CA,USA)和Bioanalyzer 2100系統(AgilentTechnologies,CA,USA)進行檢測。
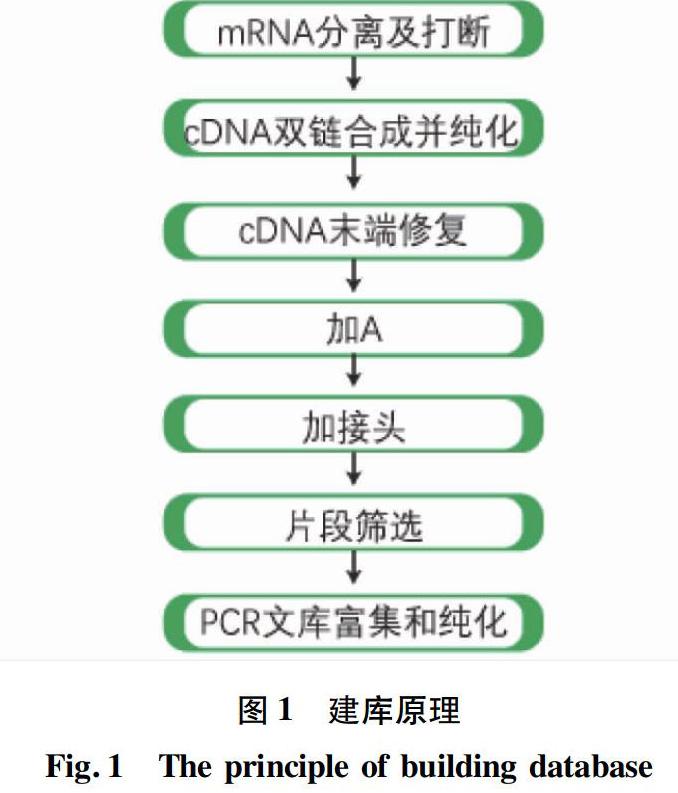
1.5 轉錄組CDNA文庫構建及質檢 試驗流程按照樣品制備試劑盒操作說明的標準步驟執行。每個樣品取3 μL的RNA用作RNA樣品準備的輸入材料構建6個測序文庫。流程如下:通過帶有Oligo(dT)的磁珠富集具有polyA尾巴的真核mRNA后,用超聲波將mRNA打斷。以片段化的mRNA為模版,隨機寡核苷酸為引物,在M-MuLV逆轉錄酶體系中合成cDNA第一條鏈,隨后用RNaseH降解RNA鏈,并在DNA polymerase Ⅰ 體系下,以dNTPs為原料合成cDNA第二條鏈。純化后的雙鏈cDNA經過末端修復、加A尾并連接測序接頭,用AMPure XP beads篩選200 bp左右的cDNA,進行PCR擴增并再次使用AMPure XP beads純化PCR產物,最終獲得文庫。建庫原理如圖1所示。
1.6 轉錄組測序
文庫質檢合格后用Illumina HiseqTM2500平臺進行pairedend測序,測序完成后將得到的序列進行去接頭、去低質量閱讀框和去污染處理,得到干凈序列。對得到的序列進行GC含量和Q30分析,以保證測序質量。
1.7 SNP挖掘及統計分析
為了保證數據質量,首先通過去除含adapter的reads,含N比例大于10%的reads,含N比例大于10%的reads,全部都是A堿基reads和低質量reads (質量值Q≤20的堿基數占整條read的50%以上),對原始數據進行數據過濾。然后利用GATK2軟件包進行SNP檢測。
2 結果與分析
2.1 RNA提取和質量檢測
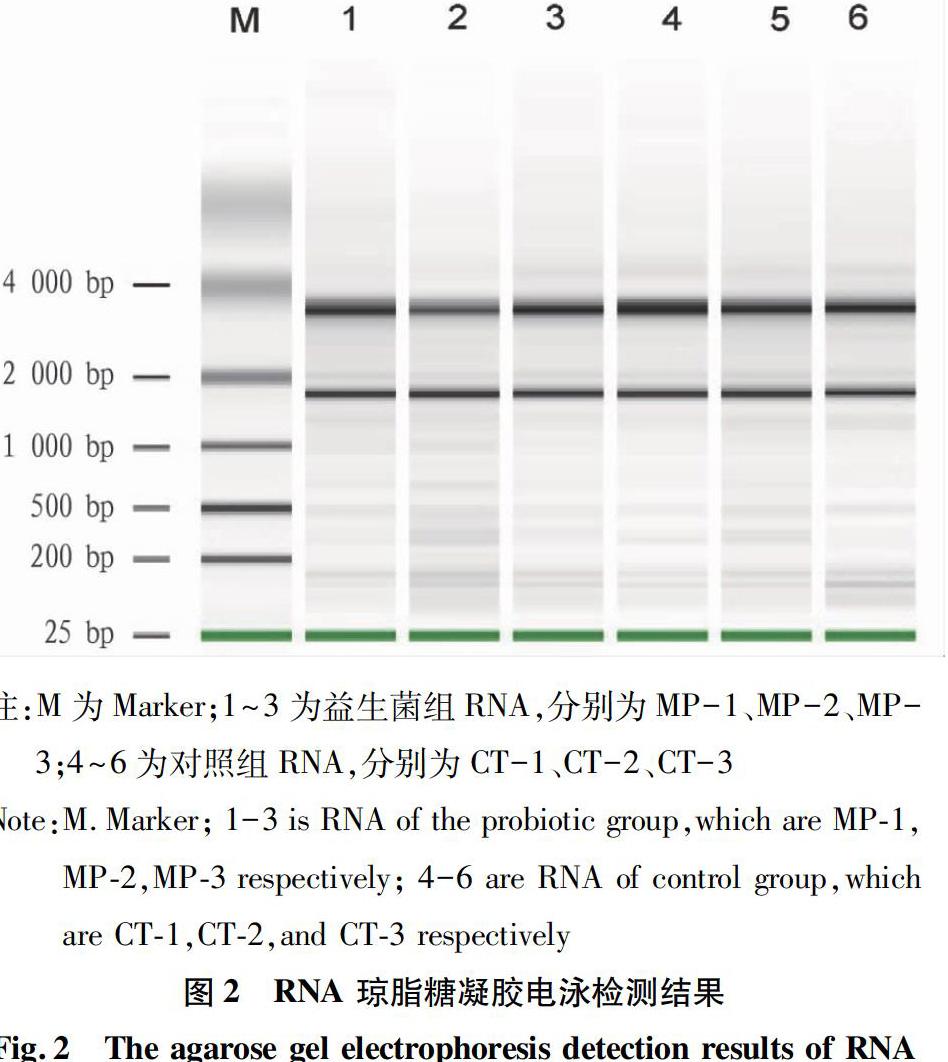
該研究提取的RNA如圖2所示,RNA樣品28S、18S、5S條帶清晰無降解;紫外分光光度法和Nanodrop結果表明OD260 nm/OD280 nm為1.8~2.0,RNA的完整性和純度都較好,達到Illumina的測序要求(表3),可用于后續的基于 HiseqTM2500平臺的轉錄組測序。
2.2 轉錄組數據分析
樣品測序產出數據質量評估情況見表4。對得到的測序數據進行錯誤率檢查,結果表明在測序長度范圍內,6個文庫中堿基的錯誤率均低于0.1%;對GC含量分布進行檢測,GC含量比例也在正常范圍內;用Fast QC分析軟件對Reads文件進行測序質量評估,每堿基質量的Q20值和Q30值最低為92.47%,表明此次測序質量較高,滿足后續分析的要求。測序所得的原始序列經過質量檢測后,過濾剔除帶接頭的、低質量的reads,6個文庫分別得到了7.3~8.7 G的clean reads,用于后續轉錄組分析。
2.3 SNP位點分析
2.3.1 SNP顛換、轉換信息。
對太平雞SNP類型的分析(表5)發現,CT組中轉換類型占22.55%~27.45%,顛換類型占11.43%~13.87%。轉換類型明顯高于顛換類型,MP組與CT組結果相似,轉換類型(22.53%~27.46%)明顯高于顛換類型(11.37%~13.94%)。轉換類型中,2組中G→A發生頻率最高,分別為27.46%和27.45%。顛換類型中,G→C發生頻率最高,分別為13.94%和13.84%%。可能是因為CG中的C(胞嘧啶)常為甲基化的,自發地脫氨后即成為T (胸腺嘧啶)。
2.3.2 SNP位置分析。對太平雞回腸SNP轉錄組序列位置變異類型進行分析,CT組和MP組總的SNP位點分別為4 684 072和4 647 171個,其中位于內含子和基因間區的SNP位點最多,位于內含子的SNP位點在CT組占59.69%,MP組中占59.94%。位于基因間區的SNP位點在CT組占20.56%,MP組中占20.12%。另外,在基因組中不同的2個位置間,存在著共有的SNP位點,CT組中,3′-UTR與5′-UTR中共有的SNP位點為603個,上游區與下游區共有的有3940個,外顯子與剪接位點中的為66個;MP組中,3′-UTR與5′-UTR中共有的SNP位點為606個,上游區與下游區共有3 879個,外顯子與剪接位點中的SNP位點為70個,如圖3所示。
2.3.3 SNP功能類型分析。
對太平雞回腸SNP的功能類型進行分析,發現主要存在一些小規模的突變,其中同義突變SNP占比最高,CT組為7.01%,MP組占比為7.10%;非同義突變SNP在CT組中占比為2.97%,MP組占比為2.98%。其他突變既移碼缺失、終止密碼突變、非移碼缺失、移碼插入、非移碼插入的SNP位點相繼減少,另外CT組和MP組中分別存在未知功能的58個SNP位點(圖4)。
3 討論
新一代的測序技術出現后,通過轉錄組測序篩選獲得SNP已成為開發應用于遺傳多樣性分析、分子育種等分子標記的重要技術手段,此技術相較于基因組篩序獲得分子標記,不僅能大大降低成本,且能快速精且能快速精準地獲得具有較高通用性的有用序列,使得開發利用分子標記更加有效便捷[15-16]。
該研究對太平雞轉錄組進行了初步研究,采用高通量測序技術對太平雞的回腸組織進行轉錄組測序。對太平雞SNP位點進行分析,MP共獲得SNP位點數4 647 171個,CT中含SNP 4 684 072個,SNP類型中轉換類型明顯高于顛換類型,MP和CT組中轉換類型分別達到27.46%和27.45%,理論上發生轉換的概率與發生顛換概率的比值應該等于0.5 (1∶2),但有些生物的比值常常>0.5,這種差異被稱為“轉換偏差”[17]。該研究發現轉換概率約為顛換的1.98倍,大于理論值,說明存在轉換偏差,這可能與堿基組成及進化過程的選擇機制有關,表明堿基轉換突變并不是隨機產生[17]。另外,該研究發現G/A發生頻率在轉換類型中最高,這可能與在CG序列上出現的高頻率,胞嘧啶極易甲基化有關。
對太平雞回腸轉錄組序列SNP位置進行分析,CT組和MP組總的SNP位點分別為4 684 072和4 647 171個,其中位于內含子和基因間區的SNP位點最多,位于內含子的SNP位點在CT組占59.69%,MP組中占59.94%。位于基因間區的SNP位點在CT組占20.56%,MP組中占20.12%。SNP可在DNA、RNA和蛋白質不同水平影響基因的功能,尤其是位于編碼區域內的編碼SNP (cSNP)與基因的表達相關,直接影響著功能基因的作用途徑[18]。通過cSNP分布情況以及功能分析可以得到許多與物種生長發育性狀相關的重要資料[19]。位于基因調控區的SNP(pSNP)則會影響基因表達量[20]。因此,研究這兩類SNP在太平雞的功能和疾病發生發展方面具有更重要的意義。
對太平雞突變類型的功能SNP進行分析,同義突變SNP占比最高,CT組中為7.01%,MP組中占7.10%;非同義突變SNP在CT組中占比為2.97%,MP組中占2.98%。其他突變既移碼缺失、終止密碼突變、非移碼缺失、移碼插入、非移碼插入的SNP位點相繼減少。編碼區內的同義SNP造成的編碼序列的變化不會引起氨基酸序列變化;而非同義SNP則會影響蛋白質序列,導致生物性狀改變[21]。因此,開發太平雞編碼區非同義SNP具有重要的生物學意義。
從太平雞回腸轉錄組獲得的SNP位點標記可以進一步用來完成后續群體的進化分析和特異性SNP標記的開發,對太平雞進行遺傳改良,資源鑒定分析、高密度遺傳連鎖圖譜的構建、提高養殖效益和健康化養殖水平以及分子標記輔助育種提供依據。
參考文獻
[1]劉媚媚,徐魏芹,李金錄,等.太平雞遺傳多樣性及系統發育研究[J].基因組學與應用生物學,2016,35(7):1731-1737.
[2]巨玉鑫,郭志明,史兆國,等.太平雞品種資源現狀及發展建議[J].畜牧獸醫雜志,2019,38(3):55-56,59.
[3]萬太軍.太平雞飼養管理技術[J].農村養殖技術,2013(8):39-42.
[4]陳永亮,劉丹丹.抗生素在雞體內殘留及控制技術研究[J].家禽科學,2018(12):55-58.
[5]伏春燕,張燕,姚啟蒙,等.益生菌對肉雞生產性能和免疫機能的影響[J].山東農業科學,2020,52(2):111-116.
[6]劉維,勵飛,聶勇.益生菌的作用機理及在養豬生產中的應用研究進展[J].飼料博覽,2019(4):23-26.
[7]VANDEPLAS S,DUBOIS D R,BECKERS K,et al.Salmonella in chicken:Current and developing strategies to reduce contamination at farm level[J].Journal of food protection,2010,73(4):774-785.
[8]侯成立,季海峰,周雨霞,等.益生菌的作用機制及其在母豬生產中的應用[J].中國畜牧獸醫,2011,38(7):20-22.
[9]葛影影,何國戈,鄭經成,等.益生菌對禽消化道疾病防治作用研究進展[J].安徽農業科學,2020,48(6):23-24,29.
[10]LANDER E S.The new genomics:Global views of biology[J].Science,1996,274(5287):536-539.
[11]黃琳.牙鲆免疫相關基因及熱休克蛋白基因的轉錄表達[D].青島:中國科學院研究生院(海洋研究所),2015.
[12]郭曉峰,鄭先虎,匡友誼,等.利用GoldenGate分析的高通量SNP芯片對鯉基因進行分型[J].水產學雜志,2016,29(6):19-25.
[13]金方彭,周睿,李光華,等.基于SLAF-seq技術的魚康浪白魚SNP位點開發[J].水產科技情報,2018,45(2):85-89.
[14]CHEN S F,ZHOU Y Q,CHEN Y R,et al.fastp:An ultrafast allinone FASTQ preprocessor[J].Bioinformatics,2018,34(17):i884-i890.
[15]SYVNEN A C.Accessing genetic variation:Genotyping single nucleotide polymorphisms[J].Nature reviews genetics,2001,2(12):930-942.
[16]孫揚,郭寶英,祁鵬志,等.基于轉錄組的曼氏無針烏賊SSR與SNP位點信息分析[J].浙江海洋大學學報(自然科學版),2019,38(2):100-106.
[17] LI H J,LIU M,YE S,et al.De novo assembly,gene annotation,and molecular marker development using Illumina paired-end transcriptome sequencing in the clam Saxidomus purpuratus[J].Genes & genomics,2017,39(6):675-685.
[18]化文平,韓立敏,魏磊,等.基于盾葉薯蕷轉錄組的SNP和SSR位點分析[J].分子植物育種,2017,15(10):4003-4009.
[19]聶鴻濤,姜力文,鄭夢鴿,等.大竹蟶高通量轉錄組測序數據組裝和分析[J].大連海洋大學學報,2017,32(6):658-663.
[20]ZHAO H,LI Q Z,LI J,et al.The study of neighboring nucleotide composition and transition/transversion bias[J].Science in China series C:Life sciences,2006,49(4):395-402.
[21]田義珂,王彩虹,白牡丹,等.基于梨貝殼杉烯氧化酶基因PpKO序列的功能性SNP標記[J].園藝學報,2012,39(10):1876-1884.
