山西省酸棗遺傳多樣性及遺傳結構分析
2020-12-08 08:03:26胡曉艷杜淑輝王兆山韓有志
林業科學研究 2020年5期
胡曉艷,杜淑輝,王兆山,韓有志*
(1. 山西農業大學林學院,北方功能油料樹種培育與研發山西省重點實驗室,山西 太谷 030800;2. 中國林業科學研究院林業研究所,北京 100091)
野生種質資源具有豐富的遺傳變異,能抵御各類生物或非生物脅迫的影響,其蘊含的有益基因對于栽培植物遺傳改良至關重要。然而,由于生境破壞以及全球氣候變化等對野生居群存續和自然進化的影響,其保護甚為迫切[1]。開展野生種質資源遺傳多樣性研究有助于制定其保護策略。酸棗(Ziziphus jujuba var. spinosa (Bunge) Hu ex H.F. Chow)是栽培棗的近緣野生種質,在我國分布范圍廣泛,華北、西北、東北和華東等地區均有分布,集中分布在山西、陜西、河北、山東等地[2]。酸棗適應能力強,抗旱耐寒、耐貧瘠,樹勢強健,根系深廣發達,具有很強的水土保持作用。由于人為活動的影響和資源的不合理開發利用,酸棗的分布正日益縮減[3]。開展酸棗種質資源遺傳多樣性研究,將為其有效保護和可持續利用提供科學依據,同時推動棗相關育種研究進程[4]。山西省是我國紅棗主要產區之一,也是酸棗主要分布區之一,但是目前酸棗資源面積逐年減少,開發利用嚴重不足[3]。
單拷貝核基因標記(single-copy nuclear gene markers)具有易于擴增測序、含有大量SNPs、不存在譜系分選等優點,目前在植物遺傳多樣性及譜系地理研究中得到廣泛應用[5-6]。栽培棗基因組序列的公布[7-8],使得利用生物信息學方法挖掘棗屬物種中通用的單拷貝核基因標記成為可能。本研究篩選酸棗中的單拷貝核基因標記并分析山西省酸棗居群的遺傳多樣性和遺傳結構,為后續酸棗資源的保護及可持續利用奠定基礎。
1 材料和方法

圖1 采樣點分布圖Fig. 1 Distribution of sample sites in Shanxi
1.1 樣品采集
本研究采集了山西省14 個野生酸棗居群213 個單株的葉樣。采樣地點、居群編號、樣品個數等具體信息見圖1、表1。選取居群內生長勢良好、無病蟲害的單株,且相鄰單株間距至少100 m,采集新鮮葉片置于變色硅膠中帶回實驗室用于總DNA 的提取。
1.2 標記篩選與擴增測序
使用改良的CTAB 法提取總DNA。所提DNA經1.0% 瓊脂糖凝膠電泳檢測后于-20℃儲存備用。采用以下方法篩選酸棗中的單拷貝核基因標記:(1)利用Duarte 等[9]提供的單拷貝核基因標簽在GENE(http://www.ncbi.nlm.nih.gov/gene)中檢索,獲得相應的核苷酸序列;(2)將核苷酸序列在棗基因組中進行BLAST 檢索(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch&PR OG_DEF=blastn&BLAST_PROG_DEF=megaBlast&BLAST_SPEC=OGP__326968__182558),以確定棗基因組中與其同源的序列,將此同源序列確定為參考序列;(3)利用參考序列進行核苷酸BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi) 以確定所選基因在棗基因組中為單拷貝;(4)根據選定的參考序列,利用Primer Premier 5.0 軟件設計引物。通過檢索分析初步篩選了50 個單拷貝核基因標記并設計了相應引物。選取3 個居群總共15 個樣品(DTYG、JZTG、YCRC 各5 個樣品)對篩選出的單拷貝核基因標記的變異情況進行檢測。PCR 擴增采用25 μL 體系:50 ng DNA,dNTP 0.24 mmol·L-1,Mg2+2.0 mmol·L-1, Taq 酶1.5 U, 引 物0.24 μmol·L-1(所有試劑均為北京擎科生物科技有限公司生產)。PCR 擴增程序為:94℃預變性5 min;94℃變性30 s,退火90 s(退火溫度隨引物而定),72℃延伸90 s,30 個循環;72℃延伸5 min;10℃保存。擴增產物用1.5% 瓊脂糖凝膠電泳檢測后采用DNA 純化試劑盒( Amersham Pharmacia Biotech,Piscataway, USA) 純化后, 利用ABI3730 DNA analyzer (Applied Biosystems, Foster City, Califonia,USA) 直接測序。擴增和測序采用相同引物。在標記篩選時未選用含有poly-結構的參考序列,因此,不存在需要使用克隆測序的情況。當所選標記在15 個樣品中擴增或測序結果差,或不包含變異信息時,棄用該標記。最終確定6 個單拷貝核基因標記對所有樣品進行擴增測序(表2),PCR 擴增和測序步驟與標記篩選時一致。經過擴增測序得到了所有酸棗樣品在6 個單拷貝核基因標記的序列數據。

表1 酸棗居群采樣信息 Table 1 Sampled populations of Z. jujuba var. spinosa in this study
1.3 數據統計與分析
測序結果處理時遵循簡并堿基的原則。所有序列數據采用軟件ClustalX[10]比對,并在BioEdit[11]中進行校正后輸出用于后續分析的數據文件。利用DnaSp5.10[12]對每個單拷貝核基因標記計算如下參數:多態核苷酸位點數目(S)、單倍型多樣性(Hd)[13]、π(一對序列間每堿基平均差異值)[14]、基于多態核苷酸位點數目S的Watterson’s 參數θw[15]、Tajima’D中性檢驗(顯著性檢驗由999 次聯合模擬計算得到)[16]、重組事件(Rm) 和居群間基因流Nm。利用DnaSp5.10 計算居群水平的遺傳多樣性參數S、Hd、π和θw。同時利用DnaSp5.10中的失配分布分析(Mismatch distribution analysis)[17]檢驗居群歷史動態,當居群大小保持相對穩定時,失配分布曲線會呈多峰分布;當居群經歷擴張或持續增長時,失配分布曲線一般會呈現單峰。利用Arlequin 3.5.1.3[18]中的分子方差分析(AMOVA) 計算在每個單拷貝核基因標記物種內居群間以及居群內差異對總變異的貢獻率(顯著性檢驗利用999次重復模擬得到)和居群間遺傳分化指數Fst[19](顯著性檢驗利用999 次重復模擬得到),同時計算兩兩居群間遺傳距離。使用GenAlEx 6.5[19]中的Mantel test檢驗遺傳距離和地理距離之間的相關性。采用STRUCTURE 2.3.4[20]進行群體水平的遺傳結構分析,參數設置如下:選用混合模型(admixture model) 和群體間相關聯的等位基因頻率模型( correlated alleles frequencies), length of burnin 長度設置為500 000,馬爾科夫鏈蒙特卡洛迭代次數(MCMC replicates after burn-in)設置為2 000 000,分類數目(K)值范圍為1~10,每個K值重復計算10 次。利用最大值ΔK對應的K值確定為最佳K值[21]。
2 結果
2.1 山西酸棗遺傳多樣性
6 個單拷貝核基因標記的長度在199~846 bp 之間,分離位點數目和單倍型多樣性的變化區間為7~64 和0.663~0.979;核苷酸多樣性參數π和θw的平均值分別為0.007 20 和0.009 25;Rm的平均值為8,表明酸棗基因組內重組事件發生頻率較高(表3)。在居群水平上,單倍型多樣性平均值為0.986,π和θw的平均值分別為0.006 50 和0.005 43,多樣性較高的居群如YQYX(π=0.007 89,θw=0.007 19),較低的如XZDX(π=0.005 02,θw=0.003 48)(表4)。

表3 6 個單拷貝核基因標記多樣性Table 3 Nucleotide variation of single-copy nuclear gene markers

表4 14 個山西省酸棗居群遺傳多樣性水平Table 4 Nucleotide variation of 14 Z. jujuba var. spinosa populations in Shanxi
2.2 山西酸棗遺傳結構
AMOVA 分析結果(表5)表明:居群內變異占總變異的85.48%,居群分化指數Fst平均值為0.145,表明山西省酸棗居群間基因交流平均水平較低,這與Nm分析結果基本一致(Nm平均值為1.33)。居群間遺傳距離結果(表6)表明:XZDX和DTYG 間遺傳距離最大(0.360),YCRC 和CZQX 間最低(0.025)。Mantel test 檢驗表明:居群間地理距離與遺傳距離沒有相關性(r=0.177 5,P=0.216)。STRUCTU 分析顯示:當K=2 時,山西省酸棗居群有最佳聚類結果,并沒有形成明顯的遺傳結構,部分居群間個體遺傳組成差異較大(如DTYG 和JZTG)(圖2)。

表5 山西酸棗居群遺傳結構分析Table 5 AMOVA and Fst analysis of Z. jujuba var. spinosa populations in Shanxi

表6 山西酸棗居群間遺傳距離Table 6 Genetic differentiation between Z. jujuba var. spinosa populations in Shanxi
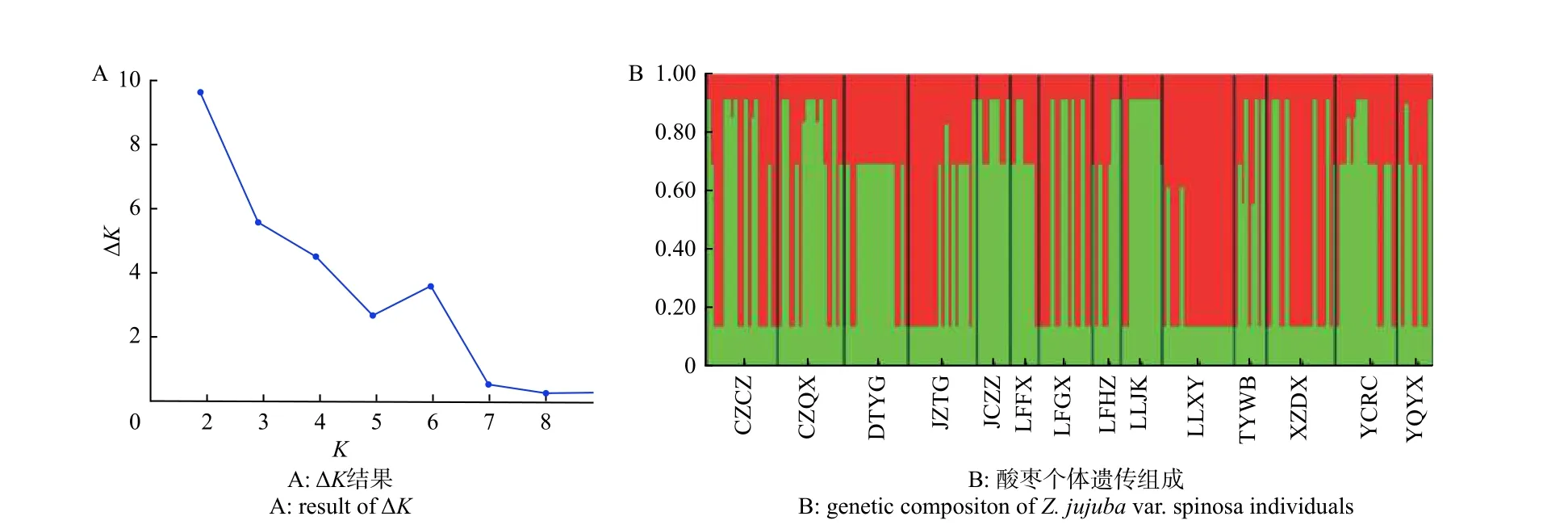
圖2 STRUCTURE 運行結果圖Fig. 2 Result of STRUCTURE
2.3 山西酸棗居群動態
Tajima’D檢驗結果均不顯著且大部分為負值,表明所選用的單拷貝核基因標記符合中性進化假設,且山西酸棗居群在進化歷史上可能經歷過居群擴張(表3)。失配分析結果呈現明顯的單峰,表明山西省酸棗自然野生居群歷史上經歷過擴張,這與Tajima’D 檢測結果一致(圖3)。

圖3 失配分布分析結果Fig. 3 Result of mismatch distribution analysis
3 討論
3.1 酸棗單拷貝核基因標記的篩選
基因組復制事件和高水平的序列變異是導致單拷貝核基因標記篩選困難的主要原因[22]。例如在566 個中國白櫟組(section Quercus)單拷貝核基因標記中,僅有19 個標記能夠在本組不同物種間通用[5]。雖然棗近期沒有發生過全基因組復制事件,但是棗染色體融合與片段復制事件發生頻率很高[7],這在一定程度上導致單拷貝核基因標記篩選的困難。標記篩選過程中,在棗基因組中成功檢索到50 個單拷貝核基因序列并設計了特異引物,但僅有6 個單拷貝核基因能夠在所有樣品中成功擴增并測序。據估計,在植物基因組中大約有10%的基因為單拷貝[9]。在棗基因組注釋到的32 808 個功能基因中[7],單拷貝基因的個數在3 000 個左右,因此,未來能夠挖掘更多的單拷貝基因標記應用于酸棗和棗居群遺傳學和譜系地理學等相關研究中。
3.2 山西酸棗的遺傳多樣性
在位點水平上,分布于山西的酸棗表現出較高的遺傳多樣性水平(π=0.007 20,θw=0.009 25),比在我國分布的其他物種如中國山楊(Populus davidiana Dode)(π=0.004 40,θw=0.007 50)[23]、甘蒙檉柳(Tamarix austromongolica Nakai)(π=0.002 59)[24]、沙芥[Pugionium cornntum (L.) Gaertn](π=0.005 32)[25]的遺傳多樣性水平高。使用其他類型分子標記技術的研究(如SSR、RAPD 等)也表明自然分布的酸棗具有較高的遺傳多樣性水平,積累了豐富的遺傳變異[26-27]。物種遺傳多樣性水平受取樣代表性、自然選擇、繁育系統及居群歷史等因素的影響[23]。本研究的取樣范圍覆蓋山西省,因此,可以排除取樣代表性對本研究結果的影響。Tajima’D 中性檢驗結果表明,6 個單拷貝核基因標記均符合中性進化假設,因此,可以排除自然選擇的影響。酸棗為異交植物,異交可以增加物種的有效群體大小和有效重組率,從而提高遺傳多樣性水平[28],并且自然狀態下酸棗居群內很少發生近交繁殖[27]。第四紀冰期循環對北半球物種的多樣性與分布產生了重大的影響[29-30],但在我國黃土高原地區冰期循環對物種的影響非常小,冰蓋也僅僅在黃土高原南部部分地區形成,持續時間非常短[31]。因此,分布于山西省的酸棗受第四紀冰期循環的影響非常小,可以保留大量的遺傳變異。同時,本研究中失配分布分析表明,山西省酸棗居群經歷過居群擴張,也支持了上述推論。
3.3 山西酸棗的遺傳結構
本研究發現,分布于山西省的酸棗居群間遺傳距離差異較大(0.025~0.360)(表6),表明酸棗居群之間的基因交流存在較大的變化,由于Mantel test 表明地理距離與遺傳距離之間沒有相關性,因此,這可能與居群之間是否存在明顯的地理隔離(如山脈、河流)或人類活動等因素相關。地理隔離可以阻礙居群之間的基因交流,而人類活動有時可以打破這種阻礙[32],因此,山西省酸棗居群間遺傳距離差異是上述因素共同作用的結果。整體上居群間表現出較高水平的遺傳分化(Fst=0.145)且基因流較小(Nm=1.33),但居群內的變異是總體變異的主要來源(表3、5)。張春梅等[27]研究了黃河流域3 個酸棗居群的遺傳分化水平,發現3 個居群間分化水平非常低(Fst=0.017~0.039)且基因流較高,推測黃河水流攜帶種子傳播是導致黃河流域3 個酸棗居群出現低分化水平的主要原因。Zhang等[33]進一步對全國范圍內酸棗居群的遺傳分化水平進行研究,發現酸棗居群間遺傳分化指數Fst為0.12。Zhang 等[33]指出,河流在酸棗居群間基因交流中(種子傳播)發揮著重要的作用;但本研究中,采樣居群間無河流連接,因此,居群間種子傳播的可能性非常小,同時酸棗花粉活力弱,擴散能力不強[34]。上述因素是導致山西省酸棗居群間表現出較高水平遺傳分化的主要原因。隨著人類活動的加劇,同時由于酸棗在棗培育過程中的使用及其種仁蘊含的藥用價值,酸棗資源不合理開發利用的情況日益嚴重,使酸棗這一寶貴的野生果樹資源得不到有效的保護與開發利用。本研究發現,分布于山西中部及南部的酸棗居群普遍表現出較高的遺傳多樣性水平,因此,在未來一方面應該加強保護,防止多樣性丟失;另一方面應加強人工馴化栽培、就地撫育改造等工作,利用好這一寶貴的種質資源[3]。
4 結論
本研究利用生物信息學方法篩選了酸棗中的6 個單拷貝核基因標記并將其應用于山西酸棗居群遺傳學相關研究,這為未來我國酸棗種質資源遺傳學及譜系地理學等研究提供了重要的技術支持。通過對山西省酸棗居群遺傳多樣性、遺傳結構及居群歷史動態的初步分析,發現山西省酸棗表現出較高的遺傳多樣性水平,繁育系統及居群歷史動態是影響遺傳多樣性的主要原因。山西省酸棗居群之間表現出較高水平的遺傳分化,未來應對分布于山西中部及南部地區的酸棗居群加強保護與開發利用。本研究的開展為深入進行山西省及我國酸棗種質資源保護、可持續開發利用等奠定了基礎。
