基于表型性狀的春蘭種質遺傳多樣性分析
2020-12-02 01:15:44傅巧娟李春楠張曉瑩趙福康
農學學報 2020年11期
關鍵詞:分析
傅巧娟,李春楠,張曉瑩,趙福康
(杭州市農業科學研究院,杭州 310024)
0 引言
春蘭(Cymbidium goeringii)又名草蘭、山蘭或朵朵香,為蘭科(Orchidaceae)蘭屬(Cymbidium)多年生草本地生植物,其味清幽,花色淡雅,具有很高的觀賞和經濟價值。廣泛分布在中國南方各省,日本與朝鮮半島南端也有分布。春蘭是栽培歷史最為悠久、分布最廣、資源最豐富、選育園藝栽培品種最多的蘭花之一[1],也是人們最為喜歡的種類之一。
種質資源是遺傳研究與育種應用的基礎,如何科學有效地評價種質資源的遺傳多樣性和遺傳關系也是資源鑒定、保護、品種改良等工作的重要內容[2]。運用形態學性狀檢測遺傳變異是最古老和簡便易行的方法,也是植物學分類的主要依據之一[3]。利用植物的表型性狀研究其遺傳多樣性已被廣泛應用于番茄[4]、蘿卜[5]、柑橘[6]、梨[7]、狼尾草[8]、紅花[9]、芍藥[10]、一串紅[11]等多種園藝植物中。近年來,張韶伊等[12]、陳君梅等[13]對部分春蘭種質進行了形態多樣性研究,但所涉及的種質及檢測的性狀較少。筆者以120份春蘭種質為材料,從50 個表型性狀研究分析其遺傳多樣性,以期為春蘭的資源鑒定、保護及利用提供理論依據。
1 材料與方法
1.1 試驗材料
供試的春蘭種質共120份(表1),每份10株以上,分別來自浙江、安徽、福建、江蘇、廣東等地,統一種植于杭州市農業科學研究院種質資源圃2年以上,統一管理。
1.2 試驗方法
1.2.1 數據采集每份種質在連續2年的開花期,選3株以上植株,參考農業部農業行業標準《植物新品種特異性、一致性和穩定性測試指南春蘭》(以下簡稱《測試指南》),選取50個表型性狀(包括38個質量性狀和12個數量性狀)進行數據采集。
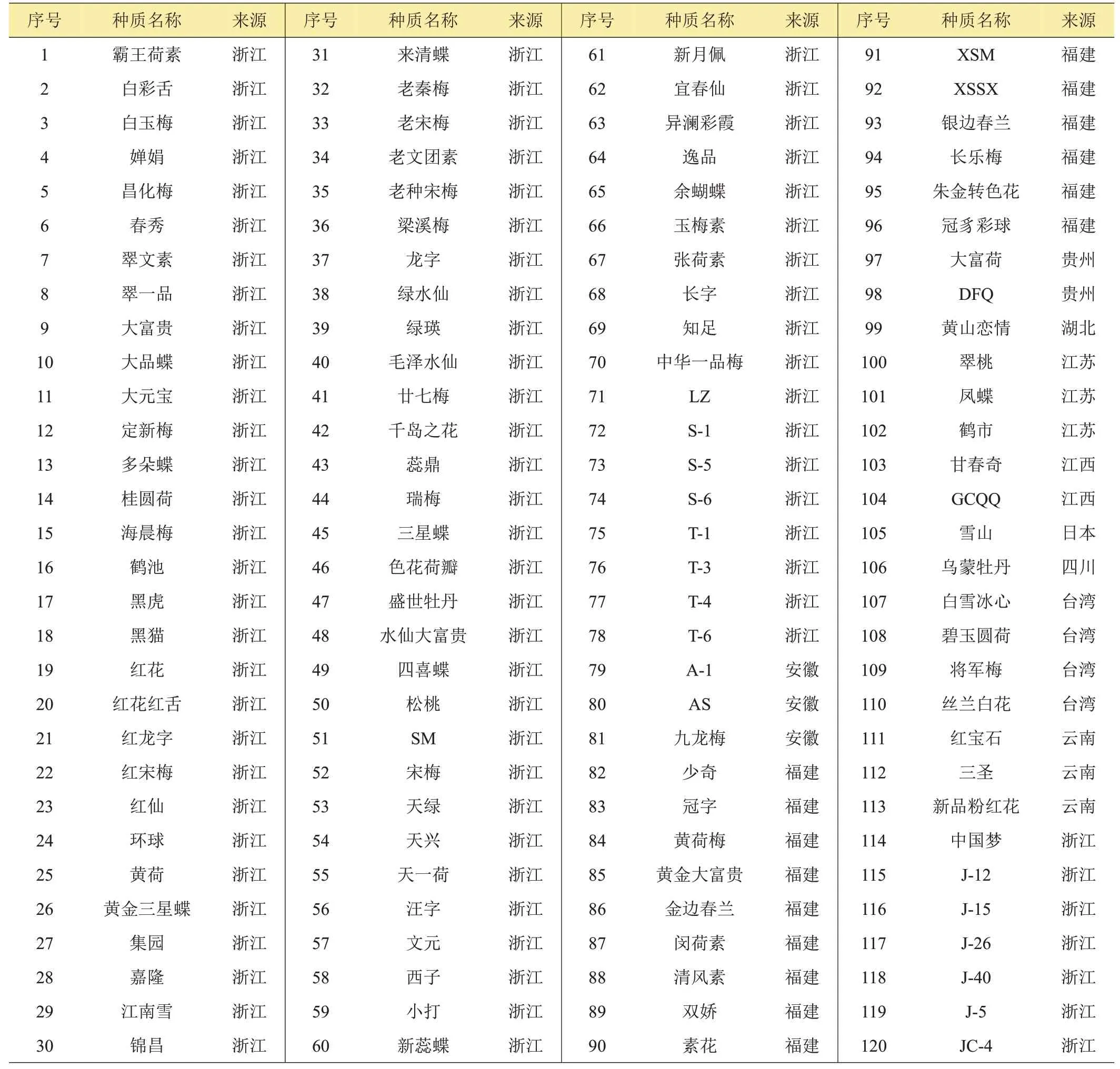
表1 春蘭種質及來源
質量性狀包括植株葉姿、大小,葉形、葉片扭曲、葉藝、葉色,開花期,花梗花青甙顯色、子房著生處苞片顏色、苞片斑點及條紋,花主色,花被片數,萼片及花瓣姿態,萼片顏色數量、主色、邊緣色、斑點、條紋、中萼外卷程度、側萼邊緣卷曲度,花瓣著生狀態、唇瓣化、顏色數量、主色、邊緣色、斑點、條紋,唇瓣主色、邊緣色、斑點、條紋及外卷程度,蕊柱數量、中部色、頂部色、斑點及藥帽顏色等,以《測試指南》為標準,通過目測獲得。
數量性狀包括葉片數、葉長、葉寬,花梗長,花朵、中萼片、側萼片、唇瓣等的長和寬,中萼片寬長比等,采用直尺測量,測量值保留1位小數。
1.2.2 數據處理 采集的數據均在Microsoft Office Excel 2007 中進行整理,分別計算各種質數量性狀的平均值(M)、最大值(Max)、最小值(Min)、極差(R)、標準差(S)和變異系數(CV),并對各性狀進行9~10級分類,1級<M-2S,10 級≥M+2S,中間每級差0.5S(M為群體性狀平均值,S為標準差),利用IBM SPSS Statistics 21軟件中非參數檢驗Kolmogorov-Smirnov(K-S)法檢驗各性狀是否遵從正態分布。對符合正態分布的性狀,依據劉孟軍[14]概率分級法,用M-1.2818S、M-0.5246S、M+0.5246S、M+1.2818S 4 個分點分為5 級,使1~5 級的出現概率分別為10%、20%、40%、20%和10%;用M-0.5246S、M+0.5246S 2 個分點分為3 級,使1~3 級的出現概率分別為30%、40%、30%。對于不符合正態分布的性狀,根據《測試指南》各性狀分級數,按照其數據的實際分布情況進行分級[15]。
質量性狀依據《測試指南》進行量化,其中顏色性狀從淺到深記為1~9。采用Shannon-Weaver多樣性指數[式(1)]分析種質多樣性[16-18]。
式中,Pi為某性狀第i級出現的頻率。
1.2.3 主成分分析及聚類分析 運用IBM SPSS Statistics 21 軟件進行主成分分析和系統聚類分析,繪制樹狀聚類圖。
2 結果與分析
2.1 質量性狀多樣性
2.1.1 有序多態性狀多樣性分析表2顯示的是有序多態性狀多樣性,可以看到,26 個質量性狀Shannon-Weaver多樣性指數(I)在0.13~1.69之間。I值最高的為蕊柱中部主色,其次是子房著生處苞片顏色1.65;相關顏色的性狀I值均較高,多樣性更豐富。I值最低的是花瓣著生狀態,其次是萼片顏色數量0.24。
2.1.2 二元性狀分布表3顯示的春蘭12個質量性狀均表現為2 種形式,即“有”或“無”。可以看到,參試的120 份材料唇瓣均無條紋;葉片無扭曲、無葉藝、萼片無斑點占87%以上,而花梗花青甙顯色、唇瓣斑點以“有”居多,占85%以上;其他5個性狀2種表現形式均占較高比例,多樣性更為豐富。
2.2 春蘭數量性狀變異分析
群體數量性狀的差異性和多樣性常用變異系數(CV)來反映,變異系數大則分化程度高、多樣性顯著,相反則群體相對穩定、一致性較高。120 份春蘭種質數量性狀的變異情況見表4。總體上看,各性狀的CV值均在13%以上,變異程度較高;其中CV值最低的為葉片數(13.5%),說明該性狀在各種質間相對穩定;其次是唇瓣長、唇瓣寬,分別為18.3%、20.4%;CV值最高的為花梗長(48.8%),多樣性豐富。
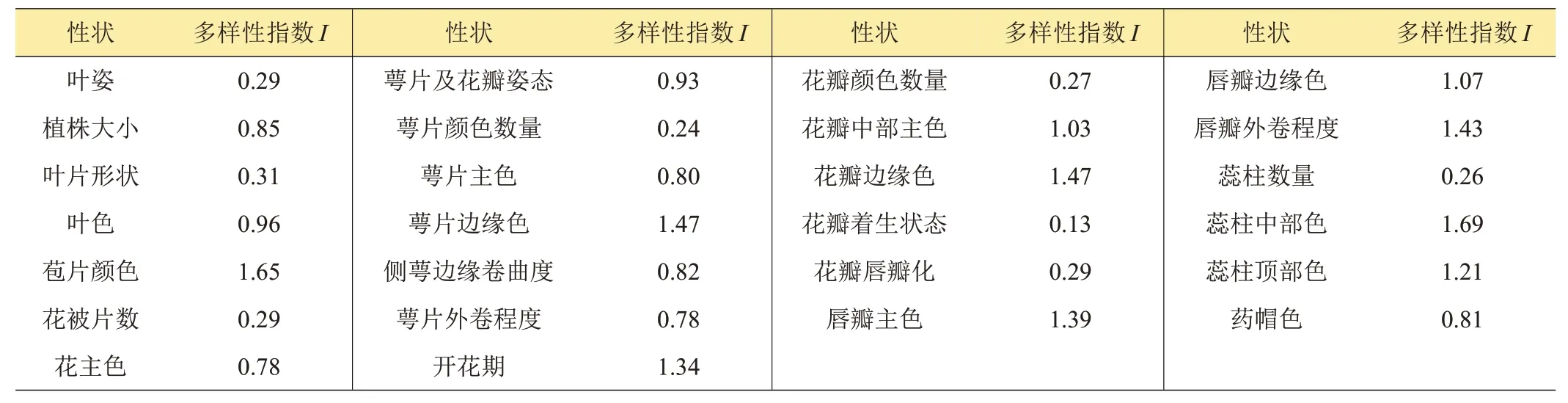
表2 春蘭質量性狀Shannon-Weaver多樣性指數比較
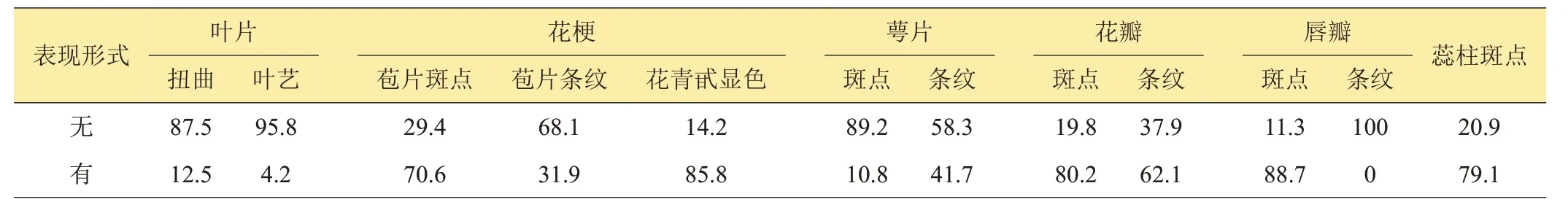
表3 二元性狀分布比例 %
2.3 數量性狀的分布規律及分級
K-S檢驗結果(表5)顯示,中萼片寬長比和側萼片寬的Sig.值大于0.05,符合正態分布,按1.2.2 進行分級;其余11個性狀Sig.值均小于或等于0.05,不符合正態分布。對11個非正態分布性狀,按照實際測量結果的10%、20%、40%、20%、10%或30%、40%、30%的比例進行5級或3級的分級,確定分點值,將數量性狀進行統一化和規范化的描述[15]。
2.4 主成分分析
從檢測的51個表型性狀指標中,選取變異較大(I值或CV值較高,二元性狀分布分散)的41 個指標(表6)進行主成分分析,按照特征值大于1的原則,提取前13 個主成分,累計貢獻率達到71.301%。第一主成分的特征值6.066,方差貢獻率最大14.795%,占主導地位;特征向量絕對值較大的性狀有中萼片長、中萼片寬、中萼片寬長比、側萼片長、側萼片寬、花朵寬、花朵長等,其中中萼片寬、中萼片寬長比、側萼片寬為負值,說明它們與其余4 個性狀之間呈負相關,決定了春蘭花朵的大小及形狀,為花朵大小因子和花型因子。第二、第三和第四主成分的特征值分別為4.853、2.667、2.309,方差貢獻率11.837%、6.505%、5.632%,主要由唇瓣長、唇瓣寬、花主色、萼片中部主色、花瓣中部主色、花瓣邊緣色、蕊柱頂部色、唇瓣外卷程度、花瓣斑點、花瓣條紋、花梗花青甙顯色等性狀決定,為花色因子、唇瓣因子。第六主成分開始主要由植株大小、花梗長、葉長、葉寬、葉片數等性狀起決定作用,稱為株型因子、葉形因子。6 個因子可視為春蘭種質評價與雜交育種中親本選擇的主要表型指標。
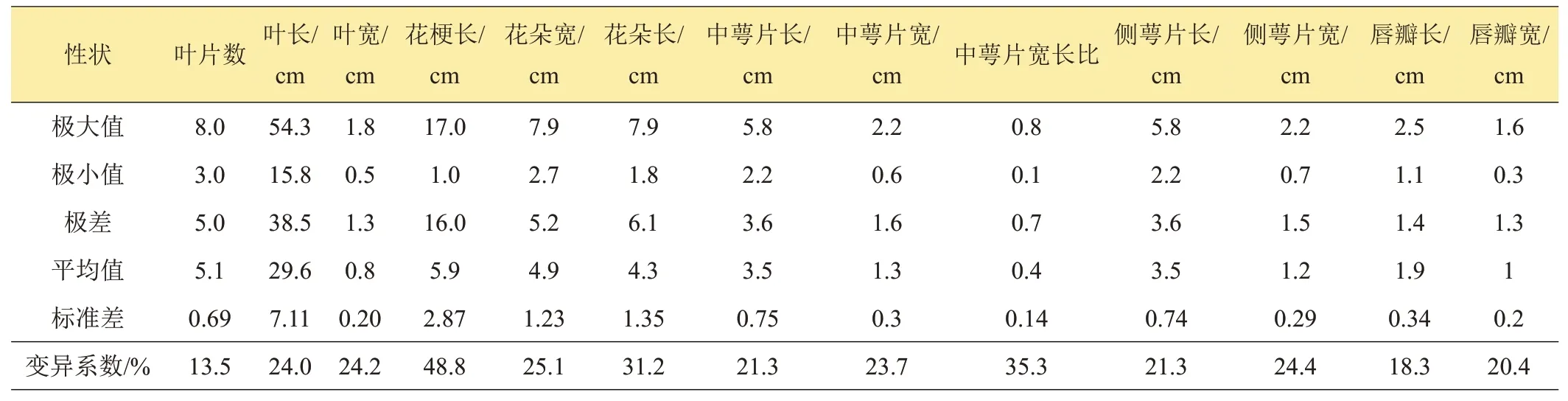
表4 春蘭數量性狀變異分析

表5 數量性狀K-S正態性檢驗及概率分級分點值
2.5 聚類分析
采用歐式距離(Euclidean distance)、Word 法對120份種質進行系統聚類,得到樹型聚類圖(圖1)。可以看到,在歐氏距離為18.1處可將群體分為2大類,第一大類(Ⅰ)包含24 份種質,主要以素心為主,唇瓣較小(表7);另外96 份種質聚為第二大類。在歐氏距離為9.2 處,第二大類又分為3 個亞類:第一亞類以奇花型為主(Ⅱ),葉片狹長,花大,但萼片較小,共11 份種質;第二亞類(Ⅲ)38 份種質,包含一些色花、蝶花等種質;其余47 份種質聚為第三亞類(Ⅳ),表現為萼片較短、寬,且基本無條紋。從聚類情況還可以看到,花朵大小、花色、唇瓣色相近的首先聚在一起,如第一大類(Ⅰ)以素心(花朵黃綠色,無斑點、條紋)為主;大元寶和大品蝶均為蝶花;銀邊春蘭、金邊春蘭、雪山和雙嬌均為覆輪復色花;冠豸彩球與DFQ、盛世牡丹與GCQQ 等均為奇花等;Ⅱ類花朵較大;Ⅳ類花朵較小、萼片較寬。與主成分分析結果一致。

表6 入選主成分的特征值、貢獻率及特征向量
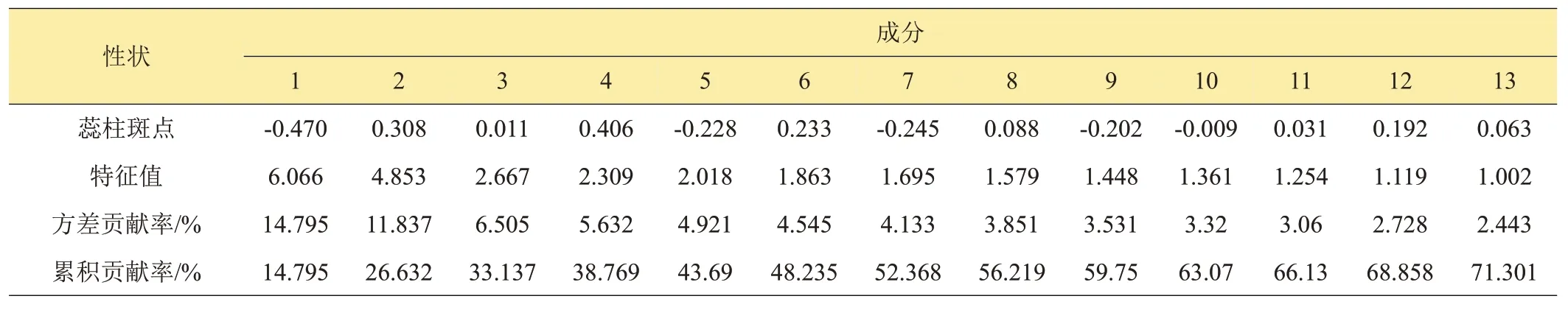
續表6
3 結論
筆者從50 個表型性狀入手,研究分析120 份春蘭種質遺傳多樣性,發現春蘭表型多樣性極為豐富,無論是質量性狀還是數量性狀,都表現出極為豐富的多樣性,與張韶伊等[12]、陳君梅等[13]的研究結果一致。其中顏色性狀Shannon-Weaver多樣性指數(I)較高,花梗長CV值最高,多樣性更豐富,與陳君梅等[13]的研究結果一致;葉片數變異相對穩定,數量性狀的分布規律多數不符合正態分布,可能與種質來源及人工選擇栽培有關。
主成分分析結果顯示,提取的前13個主成分6個因子綜合反映了春蘭的表型,但其累積貢獻率只有71.301%,說明各性狀貢獻率較為分散,累積貢獻率增長不明顯,也表明性狀變異具有多向性。聚類分析發現,花朵大小、花色、唇瓣色相近的首先聚在一起,與主成分分析結果相符;但與傳統春蘭品種按瓣形進行區分的結果不盡一致,王曉英等[1]和牛田等[19]的試驗結果也認為,聚類結果與春蘭的瓣形分類是不一致的,這可能與春蘭種質的人為選擇及多樣性有關,因此,分類標準有待進一步研究。此外,季祥彪等[20]研究表明,野生春蘭樣品的聚類具有很強的地域性;牛田等[19]研究認為,來自于相同地區的春蘭品種的親緣關系較近,與本試驗結果不完全一致,可能與春蘭在不同地區間交流引種增多有關,也可能與選擇分析的表型性狀有關,這些將在今后的研究工作中進一步完善。
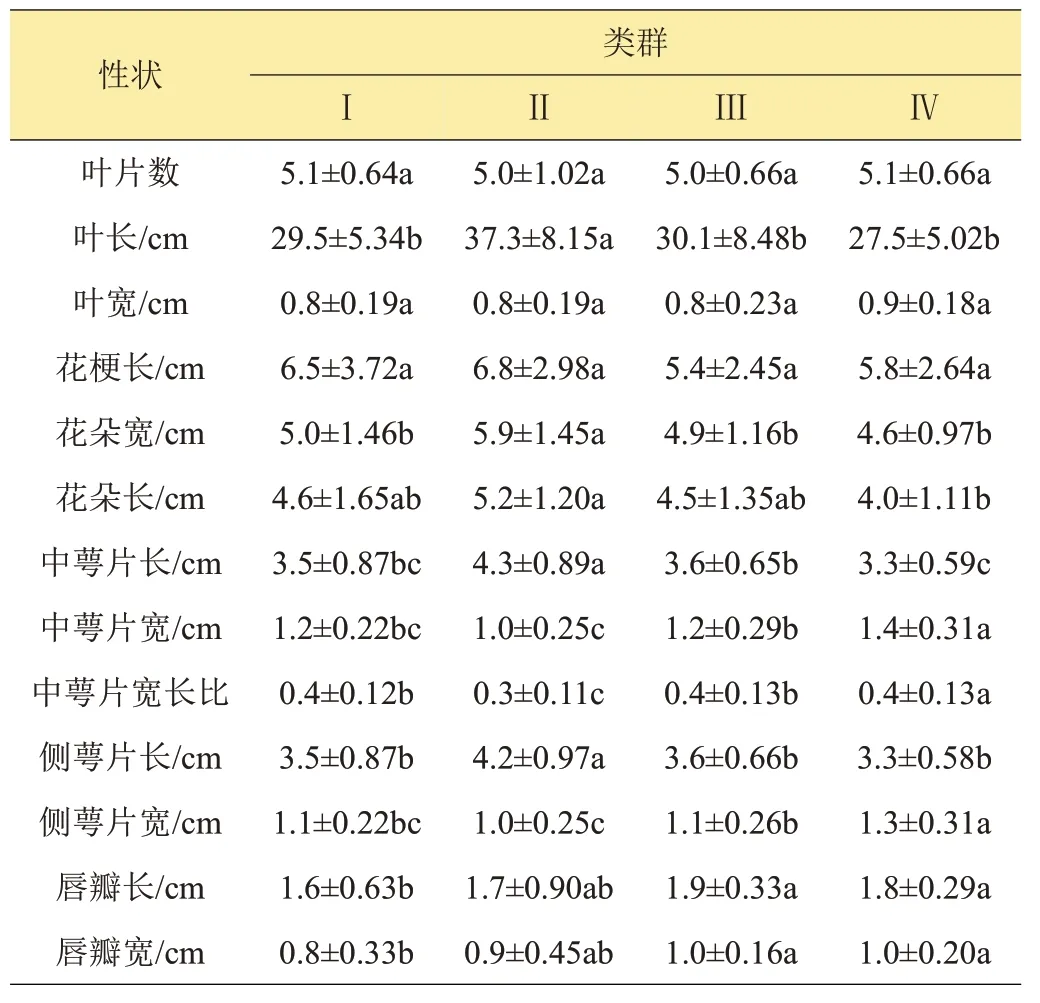
表7 4個類群13個數量性狀指標比較
4 討論
種質資源遺傳多樣性的豐富程度對品種改良、新品種選育有著直接作用。了解春蘭的遺傳多樣性水平,對解析其遺傳與進化,理解人為或生態變遷的影響力具有極大的參考價值,也是未來遺傳變異的起點和遺傳改良的前提[1]。隨著分子生物學技術的不斷發展,DNA分子標記技術已經逐漸應用到春蘭的遺傳多樣性分析及鑒定中,目前應用于春蘭的分子標記主要為SRAP[19]、RAPD[20]、AFLP[1,21]及ISSR[22-23]等。同工酶技術也應用于春蘭品種間的親緣關系分析[24]。而表型性狀是植物生長的最直觀表現,受植物本身基因型和環境因素的綜合作用,表現出穩定性和變異性共存的特點[25],長期以來,植物種質資源的分類、鑒定及育種材料的選擇通常是依據表型性狀來開展的[26-28]。當然,將表型分析與其他手段或技術結合研究種質遺傳多樣性,將更有助于資源的合理評價、鑒定、開發和利用。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
