HJURP基因促進腎癌細胞增殖遷移的機制研究
2020-11-30 16:48:24楊柳熊吟余永紅胡顯鋒石園園江文凜李艷蘭杜艷華
天津醫藥 2020年11期
楊柳,熊吟,余永紅,胡顯鋒,石園園,江文凜,李艷蘭,杜艷華△
腎細胞癌(renal cell carcinoma,RCC)是起源于腎實質泌尿小管上皮系統的惡性腫瘤,臨床80%~90%的腎臟惡性腫瘤都是腎細胞癌[1]。其發病占所有惡性腫瘤的2%~3%,在我國泌尿系腫瘤中僅次于膀胱腫瘤[2-3]。腎細胞癌起病隱匿,早期臨床表現不明顯。盡管診斷技術的改善提高了偶發性腎細胞癌的診斷率,依然有25%~30%的患者在確診時已出現遠處轉移;而晚期轉移患者治療效果不夠理想,生活質量較差[4]。深入探究腎癌發生進展的機制,尋找臨床更有效的生物標志物和治療靶標對腎癌的早期診斷和治療顯得尤為重要。
HJURP基因位于2號染色體,其與真菌蛋白Scm3結構和功能相似且高度保守,作為分子伴侶與著絲粒蛋白A(CENPA)結合,介導CENPA與染色體著絲粒的結合,從而參與調節染色體的穩定性[5-7]。目前在腫瘤研究中已經發現,HJURP在肝癌及肺癌中高表達,并且可以作為腫瘤的預后標志物[8-9],在神經膠質瘤中敲低HJURP基因可影響細胞增殖能力且增強腫瘤放射敏感性[10]。但HJURP在人腎癌中的作用尚未見報道。本文通過檢測HJURP基因在惡性腎癌組織和細胞系中的表達,并敲減腎癌細胞CAKI-1中HJURP基因,分析HJURP基因敲減后細胞增殖、周期及遷移的變化,旨在研究HJURP對腎癌細胞生物學特性的影響。
1 材料與方法
1.1 基因芯片數據提取從TCGA數據庫下載腎癌測序基因數據(TCGA-KIRP),利用R軟件讀取原始文件后,使用Affy包中的RMA算法標準化數據后得到基因的表達矩陣,將預處理后得到的基因表達矩陣文件讀入R軟件,利用R軟件中limma包對腎癌組織樣本和癌旁組織樣本基因進行差異表達分析,并應用貝葉斯檢驗方法進行多重檢驗校正。差異基因判定標準:表達值倍數變化(fold change,FC)≥2或≤-2且P<0.01。同時提取TCGA腫瘤數據庫腎癌患者生存預后信息,根據HJURP表達的中位數將患者分為HJURP低表達組及HJURP高表達組。
1.2 實驗材料組織標本選自武漢市第四醫院2016年1月—2019年10月收治的20例腎癌患者腎癌組織與對應癌旁組織標本,其中男12例,女8例,年齡38~80歲,平均(52.30±7.24)歲。腎癌細胞株CAKI-1、786O、A498及正常化腎小管細胞HK2購自中國科學院典型培養物保藏委員會細胞庫,RPMI 1640培養基購自Hyclone公司,胎牛血清購自中國四季青公司。HJURP-靶特異性siRNA(si-HJURP)由吉凱基因合成。為避免脫靶效應,實驗共設計3條si-HJURP序列:siRNA1,5′-ATCGAGCATCATCAGTATG-3′;siRNA2,5′-TAGCGCTGTATGAGACCAGTCAA-3′;siRNA3,5′-TACAGCACATGTCTGTGACAGAC-3′。空載體對照序列5′-CCTAAGGTTAAGTCGCCCTCG-3′。HJURP一抗購自英國Abcam公司,鈣黏附蛋白E(E-cadherin)、鈣黏附蛋白N(N-cadherin)、蛋白激酶B(AKT)、磷酸化蛋白激酶B(p-AKT)、糖原合成酶激酶3β(GSK-3β)及磷酸化糖原合成酶激酶3β(p-GSK3β)一抗均購自美國CST公司。GAPDH內參抗體購自美國Santa cruz公司。TRIzol試劑盒購自美國Invitrogen公司;RNA反轉錄試劑盒購自東洋紡(上海)科技有限公司,SYBR mix購自美國Bio-Rad公司。LipofectamineTM2000脂質體轉染試劑盒購自美國Invitrogen公司,MTT試劑盒購自北京百奧萊博科技有限公司,流式周期試劑盒和Transwell小室購自美國BD公司。
1.3 細胞培養及實時熒光定量PCR(qPCR)檢測HJURP表達CAKI-1、786O、A498及正常化腎小管細胞HK2培養于含10%胎牛血清的RPMI 1640培養基,于37℃、5%CO2培養箱中常規培養。取對數生長期的4種細胞,TRIzol法提取總RNA并反轉錄成cDNA。以cDNA為模板,利用Primer Bank數據庫(https://pga.mgh.harvard.edu/primerbank/)查詢并設計相應引物,HJURP引物上游5′-CCACGCTGACCTACGAGAC-3′,下游5′-CTCACCGCTTTTTGAATCGGC-3′,內參GAPDH引物上游5′-TGCACCACCAACTGCTTAG-3′,下游5′-GATGCAGGGATGATGTTC-3′。反應體系:cDNA模板1μL,SYBR mix 10μL,上、下游引物(20μmol/L)各1μL,無RNA酶水7 μL。反應條件:95℃5 min;95℃30 s,60℃30 s,40個循環;72℃30 s,72℃10 min。目的mRNA的相對表達量用內參GAPDH進行均一化,采用2-ΔΔCt法進行數據處理。
1.4 腎癌細胞株轉染實驗及siRNA敲減效率驗證取對數生長期的CAKI-1細胞接種于6孔板,待細胞密度融合度約60%時將細胞分為HJURP基因siRNA轉染組(si-HJURP組)及對照序列轉染組(NC組),分別進行小干擾RNA及對照序列空載體siRNA轉染。轉染后48 h采用qPCR檢測HJURP表達水平,檢測方法同1.3。根據qPCR結果篩選出敲減效果最佳的siRNA用于后續實驗。
1.5 細胞克隆形成實驗取si-HJURP組及NC組CAKI-1細胞,制備單細胞懸液,以800個/孔接種于6孔板,細胞培養箱溫育靜置培養2周。待細胞克隆形成,終止培養,使用4%多聚甲醛固定克隆細胞,0.1%結晶紫對細胞群落進行染色,顯微鏡下觀察并統計細胞克隆形成數量。
1.6 MTT法檢測CAKI-1細胞增殖活性取si-HJURP組及NC組轉染的CAKI-1細胞,以3 000個/孔接種于96孔板培養,培養24 h后,每隔1 d在每個孔中加入20μL MTT在37℃孵育4 h棄培養液,各孔再加入DMSO 150μL,室溫振蕩10 min。通過酶標儀測量490 nm處的光密度(OD)值,實驗重復3次。
1.7 流式細胞周期檢測將si-HJURP組及NC組細胞消化離心后加入含有5%的PI和2%的RNaseA的PBS重懸并調整細胞濃度為1×106/mL,并用錫箔紙包裹離心管避光37℃孵育30 min,然后通過流式細胞儀檢測各實驗組細胞周期分布情況,實驗重復3次。
1.8 Transwell實驗檢測CAKI-1細胞侵襲能力消化轉染后的對數生長期細胞,制備單細胞懸液,離心后用不含血清的培養液重懸細胞計數,然后在24孔板中加入600μL含10%胎牛血清的RPMI 1640培養液,取Transwell小室置于24孔板,在小室中加入200μL密度為3×105/mL的細胞懸液,培養24 h后取出Transwell小室,4%多聚甲醛固定細胞,0.1%結晶紫進行染色。顯微鏡下采集圖像,計數遷移細胞,實驗重復3次。
1.9 Western blot檢測上皮間質轉化(EMT)及AKT通路蛋白表達水平變化取2組轉染后的細胞,RIPA裂解液裂解細胞提取蛋白,BCA法檢測蛋白濃度后每孔40μg蛋白上樣,行十二烷基硫酸鈉聚丙烯凝膠電泳(SDS-PAGE),電泳結束后濕轉法(恒流17 mA轉膜過夜)將蛋白轉移于PVDF膜上,次日切取含目的條帶的PVDF膜用5%脫脂奶粉室溫封閉2 h。孵育HJURP、E-cadherin、N-cadherin、AKT、p-AKT、GSK3β及p-GSK3β一抗與GAPDH內參抗體(均為1∶10 000稀釋)于4℃過夜,TBST洗膜10 min×3次,室溫孵育二抗(1∶10 000稀釋)2 h,再用TBST洗膜10 min×3次,ECL顯影、拍照后計算目的蛋白的相對表達量,實驗重復3次。
1.10 統計學方法采用Graphpad Prism 8軟件對數據進行統計分析,符合正態分布的計量資料使用均數±標準差(±s)比較。2組間比較采用獨立樣本t檢驗,多組比較使用單因素方差分析,組間多重比較使用LSD-t檢驗,以P<0.05為差異有統計學意義。
2 結果
2.1HJURP在腎癌中的表達及其對生存預后的影響從TCGA數據庫獲取533例腎癌患者基因測序數據的基因表達信息,經生物信息學統計分析顯示HJURP在腎癌組織中明顯上調(t=12.623,P<0.05),見圖1A。進一步分析不同腫瘤分期HJURP的表達情況,發現隨著腎癌病理分期升高,HJURP表達量也增高(F=15.868,P<0.05),見圖1B。Kaplan-Meier生存曲線分析顯示HJURP高表達組患者總生存率和無病生存率較HJURP低表達組降低(P<0.01)。見圖1C、D。
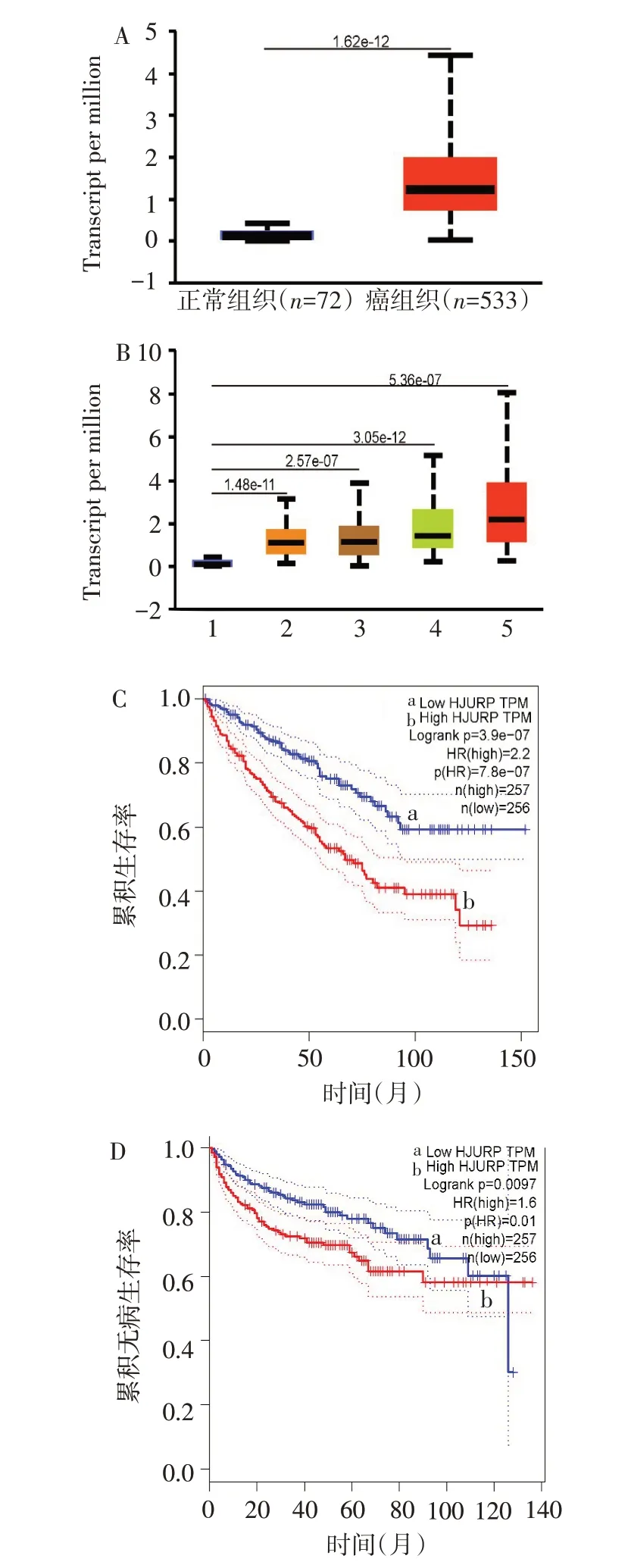
Fig.1 The expression of HJURP in renal cancer and its effect on prognosis analyzed by TCGA database圖1 TCGA數據庫分析HJURP在腎癌中的表達及對預后的影響
2.2 腎癌組織及細胞中HJURP的表達水平qPCR結果顯示HJURP在腎癌組織中的表達水平顯著高于癌旁組織(1.606±0.610vs.0.996±0.160;t=3.758,P<0.01),見圖2A。同樣,腎癌細胞株A498、786O和CAKI-1中HJURP基因表達水平高于正常化腎小管上 皮HK2細 胞株(F=14.484,P<0.01),且 在CAKI-1細胞系中表達量最高(圖2B),因此后續細胞實驗采用CAKI-1細胞系。
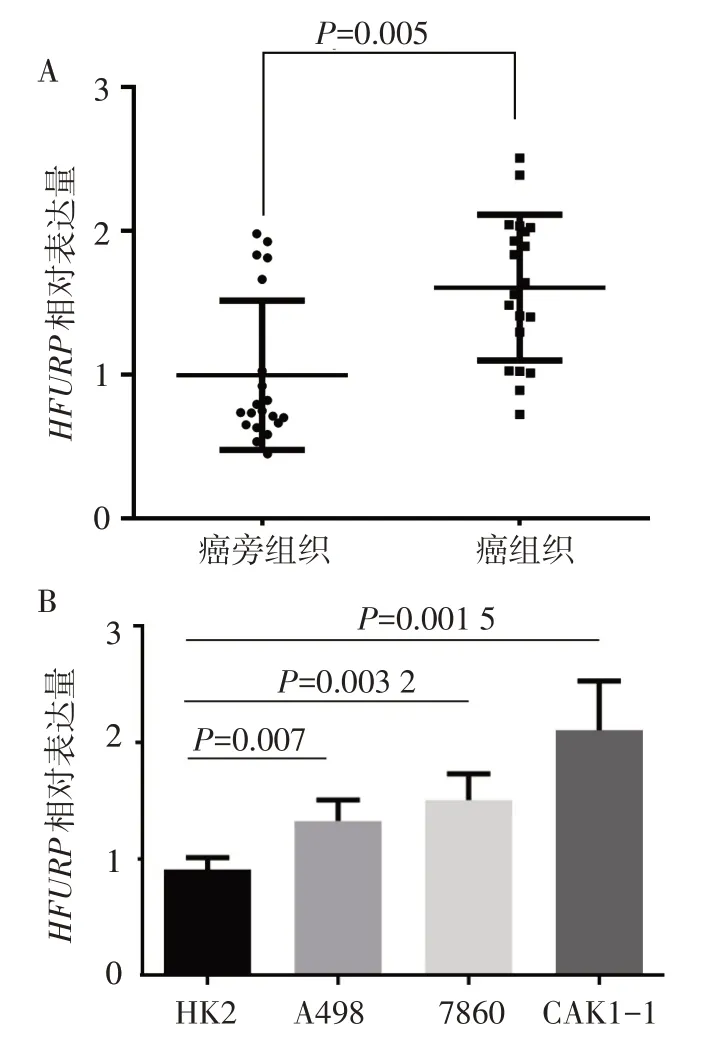
Fig.2 The expressions of HJURP in renal cancer tissues and renal cancer cells圖2組織及細胞驗證HJURP在腎癌中的表達
2.3HJURP基因敲減后CAKI-1細胞活力及克隆形成受限qPCR結果顯示3個HJURP特異性siRNA序列轉染CAKI-1細胞后,HJURP表達水平顯著低于NC組(F=205.121,P<0.01)。其中siRNA-1的敲減效果最佳,因此后續實驗采用siRNA-1進行HJURP敲減。MTT實驗結果顯示,si-HJURP組轉染后第2天開始CAKI-1細胞活性顯著低于NC組。細胞克隆形成實驗也表明,與NC組相比,si-HJURP組CAKI-1細胞的克隆形成能力顯著降低,細胞克隆形成集落顯著減少(個/視野:86.33±7.37vs.168.00±9.54,t=11.732,P<0.01)。見圖3。
2.4HJURP基因影響CAKI-1細胞周期HJURP敲減后CAKI-1腎癌細胞的G2期細胞比例明顯升高[(20.48±6.24)%vs.(53.33±7.53)%,t=14.122,P<0.01];G1期的細胞比例下降[(58.67±3.25)%vs.(33.32±5.33)%,t=16.848,P<0.01];S期的細胞比例差異 無統 計學 意 義[(18.29±1.22)%vs.(16.33±2.01)%,t=1.819,P=0.143],見圖4。
2.5HJURP影響CAKI-1腎癌細胞的體外遷移能力Transwell實驗結果顯示,si-HJURP組的細胞遷移數量顯著低于NC組[(103.0±3.5)個/視野vs.(152.3±8.7)個/視 野,t=4.714,P<0.01],見 圖5。Western blot結果顯示,HJURP敲減后CAKI-1細胞促進腫瘤細胞遷移相關的蛋白N-cadherin明顯下降,抑制腫瘤細胞遷移相關的蛋白E-cadherin明顯上調,同時AKT通路關鍵蛋白p-AKT及p-GSK3β明顯下調,見圖6。
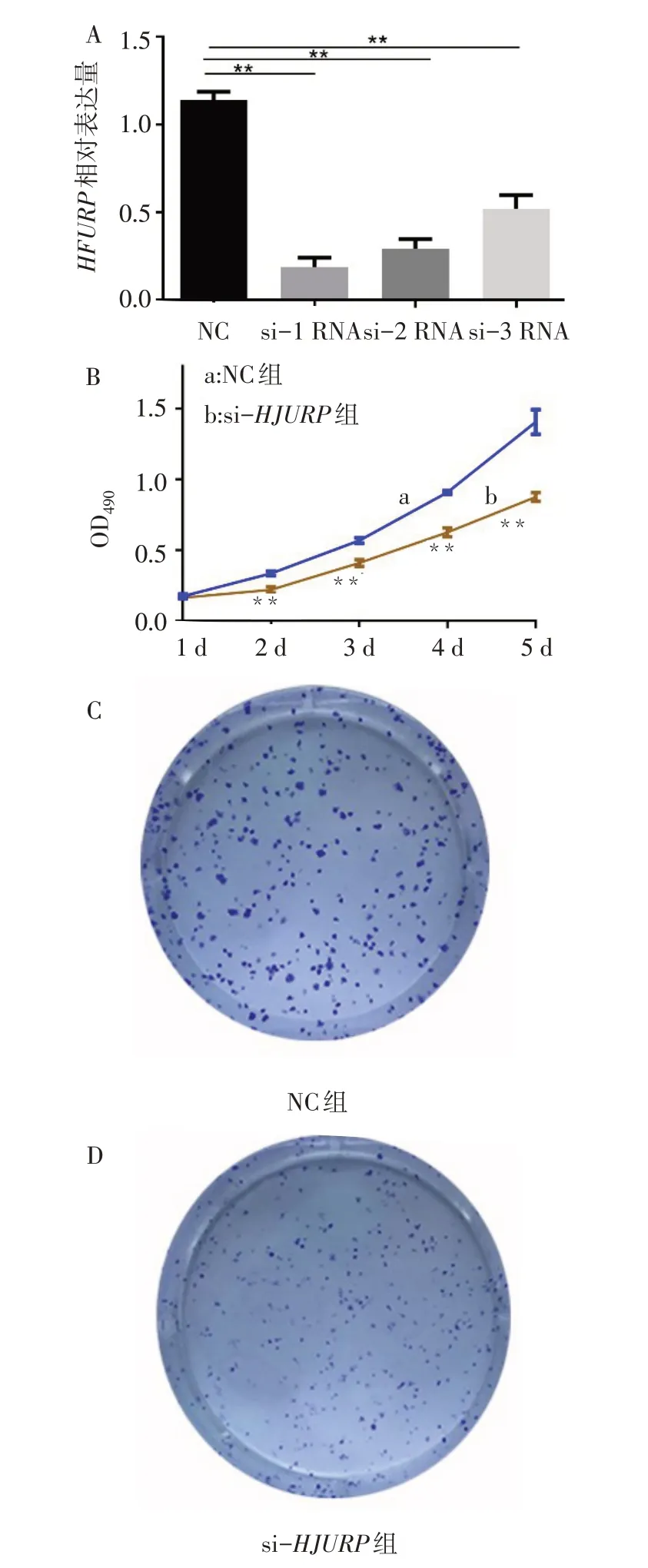
Fig.3 HJURP knockdown represses CAKI-1 viability and clone formation圖3 HJURP基因敲低降低CAKI-1細胞活力及克隆形成
3 討論
HJURP是近年來發現的一個原癌基因,在多種腫瘤中發揮重要作用。Chen等[11]發現HJURP在肝癌組織中高表達,且與肝癌患者的預后密切相關,同時通過體外細胞實驗證明HJURP通過MAPK/ERK1/2及AKT/GSK3β信號通路破壞抑癌基因P21蛋白的穩定性,從而促進肝癌細胞增殖、遷移和侵襲。在膀胱癌的研究中發現,HJURP作為原癌基因促進腫瘤的生長及轉移,敲除HJURP基因后,腫瘤細胞發生周期阻滯,增殖減緩,進一步的機制研究顯示HJURP可能通過PPARγ-SIRT1通路影響腫瘤的代謝[12]。此外另一項研究顯示HJURP在人乳腺癌細胞系和原發性乳腺癌中高表達,HJURP的表達水平影響乳腺癌患者的預后且與其放射治療結果相關,HJURP水平高的患者對放療更為敏感;體外實驗同樣表明,高表達HJURP的細胞對放射治療更為敏感,并且其凋亡率更高[13]。然而HJURP在腎癌的發生發展中的作用機制目前尚不清楚。
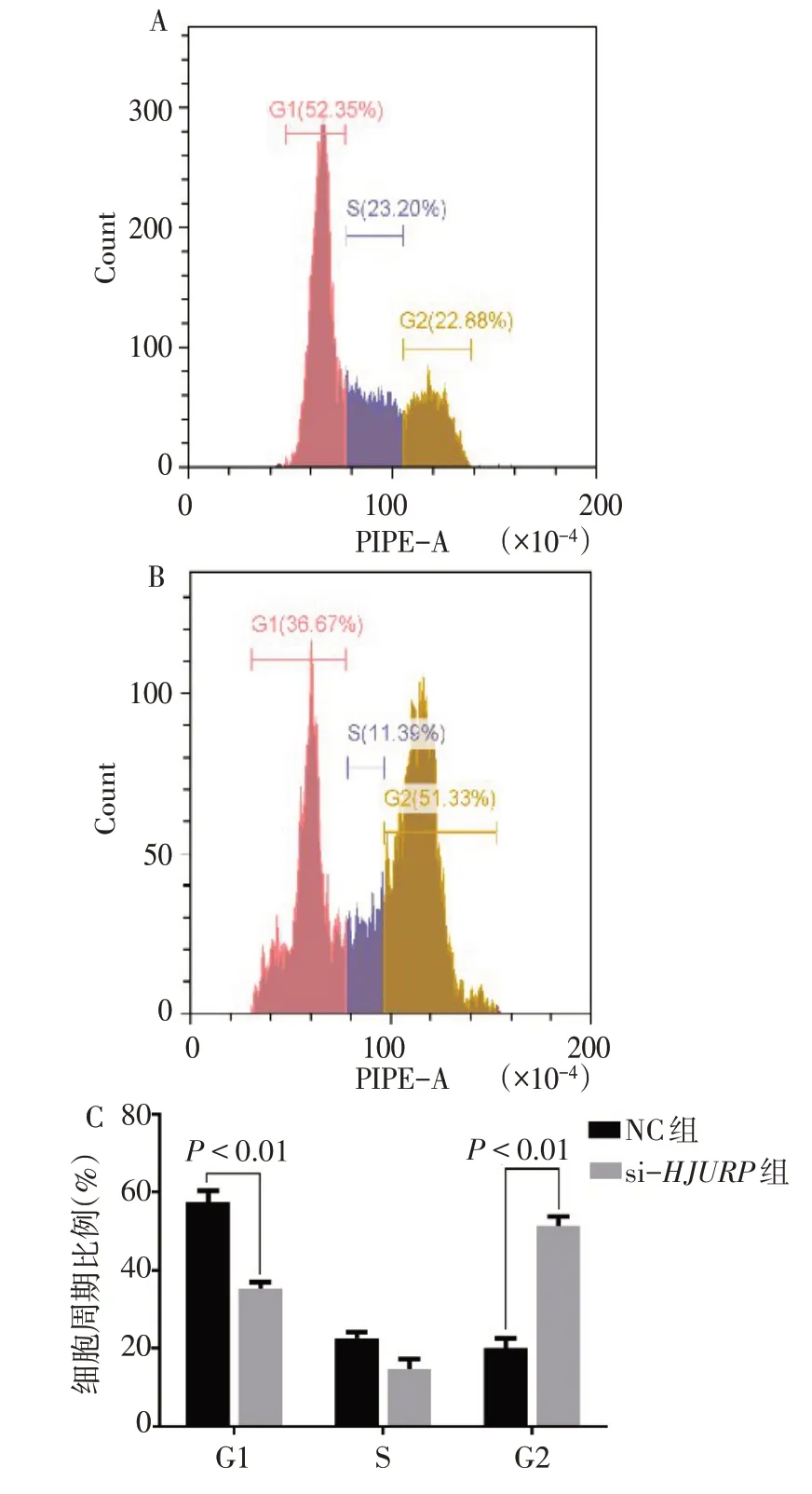
Fig.4 Flow cytometry results of HJURP knockdown in two groups圖4 si-HJURP組和NC組細胞周期分布流式圖
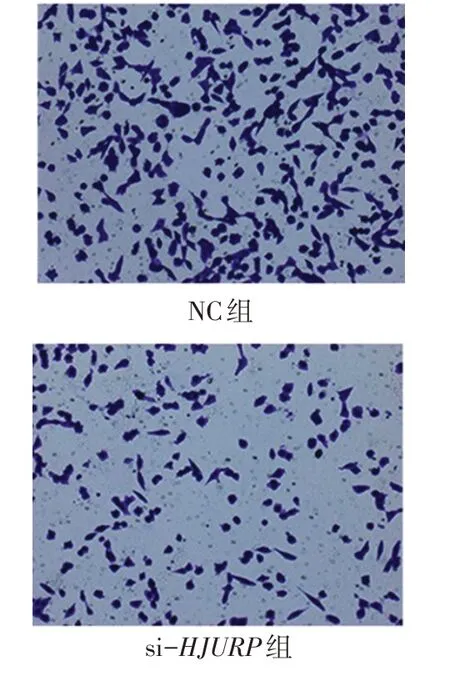
Fig.5 Changes of cell migration ability in si-HJURP group and NC group(×200)圖5 si-HJURP組和NC組細胞遷移能力變化(×200)

Fig.6 The key protein changes of EMT and AKT pathway after HJURP knockdown圖6 HJURP敲減后EMT及AKT通路蛋白表達變化
本研究利用TCGA腫瘤數據庫分析發現HJURP在腎癌中顯著高表達,生存及預后分析結果顯示,HJURP高表達組患者較低表達組預后明顯較差。進一步的臨床腎癌組織標本結果同樣證實,HJURP在腎癌組織中表達明顯高于癌旁組織,提示HJURP可能作為原癌基因參與腎癌腫瘤的形成及進展。
為進一步確定HJURP是否作為功能性的原癌基因參與腎癌進展,筆者檢測了HJURP過表達對腎癌細胞增殖和遷移的影響。HJURPsiRNA轉染CAKI-1細胞株后,發現沉默HJURP表達后腎癌細胞的增殖、侵襲和克隆能力明顯下降。細胞周期與細胞增殖密切相關,細胞周期的異常調節對腫瘤發生機制具有重要意義[14]。本研究中,沉默HJURP后引起腎癌細胞G2期阻滯,表明HJURP表達下調后可將腎癌細胞阻滯在G2期,從而抑制其在體內外的增殖能力。蛋白免疫印跡實驗結果顯示si-HJURP組促進腫瘤細胞遷移相關的蛋白N-cadherin明顯下降,抑制腫瘤細胞遷移相關的蛋白E-cadherin明顯增加,蛋白水平驗證了HJURP的表達水平確與腎癌細胞的遷移能力相關,提示HJURP可能通過EMT途徑促進腎癌細胞的侵襲遷移。AKT是在腫瘤細胞代謝、凋亡及細胞遷移等多種過程中起到重要作用的一種絲氨酸/蘇氨酸特異性蛋白激酶[15-16],AKT過度激活可促進各種腫瘤的生長與轉移。本實驗通路蛋白檢測結果顯示,HJURP基因下調后AKT通路關鍵功能蛋白p-AKT及p-GSK3β明顯下調,說明HJURP可能通過AKT通路促進腎癌細胞的侵襲遷移。
綜上所述,腎癌組織中HJURP的表達水平較癌旁正常組織升高,敲減HJURP表達可以抑制腎癌細胞的增殖轉移,其可作為腎癌的潛在臨床診治靶點及預后標志物。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
