氮磷肥料的添加對鹽堿旱田有機碳礦化和激發效應的影響
2020-11-24 08:20:36陳靜書李昭陽王靜靜
科學技術與工程 2020年29期
湯 潔, 陳靜書, 李昭陽, 王靜靜, 楊 平
(吉林大學新能源與環境學院, 長春130021)
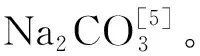
有研究發現,在農田生態系統中,施肥會影響SOC的礦化速率,影響有機質結構的穩定性,進而影響有機質的降解和積累[7]。在農業生產中,依賴大量的無機氮肥投入來維持作物產量[8],磷肥的用量雖少,但必不可缺,也不能過量[9]。氮和磷是植物生長所需的基本元素,施加氮肥和磷肥能夠改變植物多樣性以及群落組成[10],提高微生物活性,促進碳循環過程[11],增加SOC積累。當前關于添加氮、磷對SOC礦化的影響研究仍有爭議。有研究表明,氮添加會抑制原有SOC的分解[12]。而有些研究表明,氮添加會促進SOC分解或對SOC分解沒有影響[13-14]。有研究發現,磷對微生物活動有直接影響,添加磷使得微生物群落規模增加[15],從而促進SOC礦化。而有研究發現,在低生產力森林土壤中,磷對微生物呼吸有抑制作用[16-17]。盡管氮、磷的利用可能是有機碳礦化的重要機制,但這種可能性仍然需要廣泛實驗研究[1]。
激發效應是加入外源物后SOC周轉的強烈變化,加入外源物可能促進SOC的礦化,也可能抑制SOC的礦化[18]。這一現象是土壤元素、土壤理化性質與外源物質交互作用的結果。激發效應理論認為,在貧瘠的土壤中,添加外源物會增加天然有機碳的分解,從而為微生物提供所需的額外營養物質[19]。氮肥和磷肥是農田土壤中較為常見的外源添加物,SOC激發效應對不同氮、磷添加物的響應不同。基于此,通過實驗室模擬實驗,研究氮、磷添加對鹽堿旱田SOC礦化和激發效應的影響,對揭示SOC礦化機制和了解碳循環過程有重要意義。
1 材料與方法
1.1 研究區概況
于2018年5月進行現場采集,研究區位于吉林省西部,采樣點地理坐標如表1所示。吉林西部屬于溫帶大陸性季風氣候,夏季濕熱多雨,冬季寒冷降雪少、冰凍期長,年平均氣溫為6.5 ℃,年平均降水量約為400 mm,年蒸發量遠高于降水量,平均相對濕度約為60%[20],日照總時數為2 366.8~2 617.7 h。
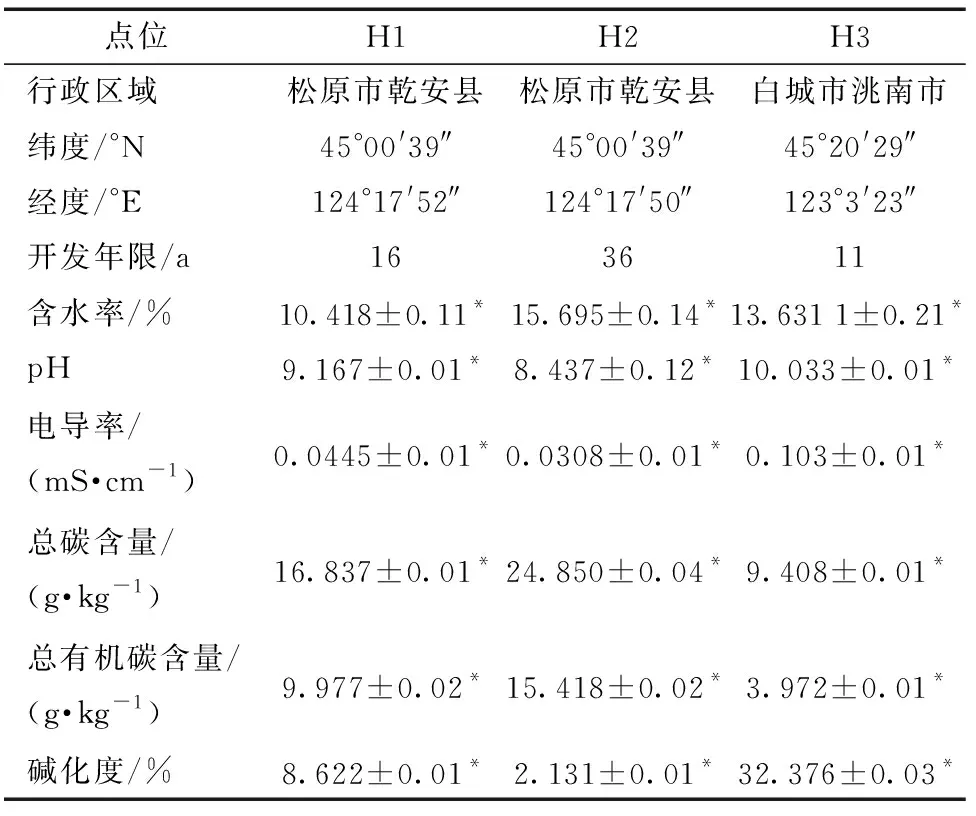
表1 采樣點位置及土壤基礎性質Table 1 Sampling point location and basic properties
1989年和2018年的吉林西部土地利用變化如圖1所示。研究區在歷史上曾是豐美的大草原,游牧民族廣泛居住在此。近30年來,在自然和人為因素的雙重驅動下,土地利用類型發生了巨大變化,土地景觀破碎[21],草地退化,耕地面積增加,土壤鹽堿化嚴重,鹽堿草地逐漸演變為鹽堿旱田。研究區旱田以種植玉米為主,為滿足糧食增產的需求,氮肥和磷肥的施用大大增加,SOC礦化作用增強,土壤環境進一步惡化。
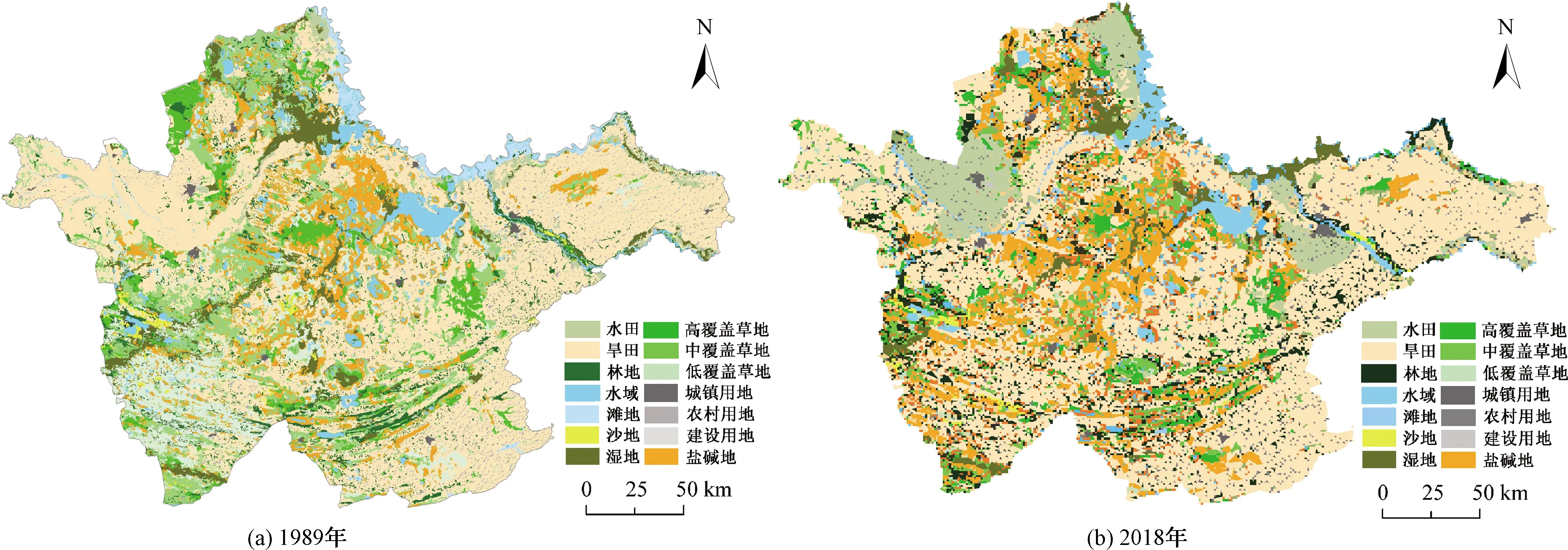
圖1 吉林西部1989年與2018年土地利用變化Fig.1 Land use change in Western Jilin from 1989 and 2018
1.2 土壤樣品預處理
采集0~15 cm表層土壤,去除肉眼可見的動植物殘體、混勻,一部分于4 ℃冷藏保存用于培養試驗,另一部分在室溫下自然風干,研磨過篩(2、0.15 mm),用于測量土壤理化性質。
1.3 培養實驗設計
使用堿液吸收法測定SOC礦化產生的CO2含量[22]。稱取定量的風干土樣,調節含水量至40%,均勻鋪于500 mL廣口瓶底部。隨后進行外6組添加處理,分別為200 mgN/kg干土的(NH4)2SO4(N1)、1 000 mgN/kg干土的(NH4)2SO4(N2)、200 mgN/kg干土的KNO3(N3)、200 mgP/kg干土的KH2PO4(P1)、1 000 mgP/kg干土的KH2PO4(P2)以及空白對照(CK)。其中,當地凈氮輸入量分別為200 mg/kg,與低濃度添加設置對應。將添加物用少量水溶解后,均勻加入廣口瓶中,調節含水量至45%。再將盛有10 mL 0.1 mol/L NaOH溶液吸收瓶懸置于廣口瓶中,密封廣口瓶,于25 ℃恒溫箱內培養。在試驗開始后的第1、2、4、9、18、32 d時更換吸收瓶,吸收瓶中加入1 mol/L BaCl2溶液2 mL,加1滴酚酞指示劑,用標準酸(約0.05 mol/L HCl)滴定至褪色,測得NaOH吸收的CO2含量,同時用稱量法進行土壤含水量的調整。將培養分為3個階段,分別為培養前期(0~9 d)、培養中期(9~18 d)和培養后期(18~32 d)。在培養后期,CO2釋放量大量增加,可適當調節吸收瓶中NaOH溶液濃度,避免CO2吸收不完全。
1.4 土壤鹽堿程度的測定
選用電位法測量酸堿度(potential of hydrogen,pH)和電導率(electroconductibility,EC),EDTA-乙酸銨鹽交換法測量陽離子交換量(cation exchange capacity,CEC),火焰光度法測量可交換性鈉離子濃度(Na+),并通過式(1)計算堿化度(exchangeable sodium percentage,ESP):

(1)
1.5 SOC礦化指標的計算
1.5.1 SOC礦化量
CK組和添加組SOC礦化量計算公式為
MC=(V0-V)CHCl22m
(2)
式(2)中:MC為CK組SOC礦化量,mg/kg;CHCl為鹽酸濃度,mol/L;V0為空白滴定值,mL;V為消耗鹽酸的體積,mL;22為1/2MCO2,MCO2為CO2的物質的量;m是實際培養土樣質量換算值。
Ma=(CNaOHV′-CHClV)22m
(3)
式(3)中:Ma為添加組SOC礦化量,mg/kg;V′為小燒杯NaOH體積,mL。
1.5.2 SOC礦化速率
SOC礦化速率計算公式為
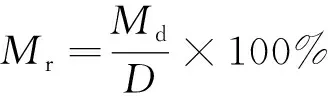
(4)
式(4)中:Mr為礦化速率,mg/(kg·d);Md為培養期間有機碳礦化量,mg/kg;D為培養天數,d。
1.5.3 激發效應
激發效應計算公式為[23]
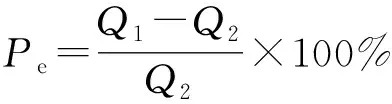
(5)
式(5)中:Pe為激發效應,%;Q1為來自加入外源物后SOC礦化釋放的CO2量,mg/kg;Q2為空白的SOC礦化釋放的CO2量,mg/kg。
2 結果與分析
2.1 土壤理化性質
H1、H2和H3表示研究區3個樣地,土壤基礎理化性質如表1所示。作為評價鹽堿程度的重要指標,pH、EC和ESP在3個樣地的大小分布均為H3>H1>H2。土壤總碳和總有機碳含量與鹽堿程度成反比,其大小分布均為H2>H1>H3。H1、H2、H3樣地土壤含水率分別為10.4%、15.7%、13.6%,土地開發年限分別為16、36、11 a。有研究認為,土壤鹽堿程度隨著開發年限的增加而減弱[24-25],這與本研究實驗結果相同,3樣地鹽堿程度與開發年限呈反比。由此可見,研究選擇樣地土壤的理化性質完全不同。
2.2 表面形態結構分析
使用激光共聚焦掃描電鏡,對土樣進行掃描,土壤表面形態如圖2所示,分別為H1、H2、H3樣地土壤顆粒放大至40 μm時的形態。從整體來看,研究區土壤顆粒形狀不規則,大小不一;從局部來看,H1、H2樣地土壤顆粒較大,土壤結構呈塊狀,土壤顆粒表面粗糙,有細塊狀、薄片狀顆粒物附著,附著物大多表現出一定的聚集;H3樣地土壤顆粒較小,表面比較光滑,附著細小的層狀或片狀顆粒物較多。

圖2 供試樣地土壤的掃描電子顯微鏡圖Fig.2 Scanning electron micrograph of soil samples
2.3 不同處理對SOC礦化量的影響
不同處理的SOC礦化累積隨培養時間的變化規律如圖3所示,添加氮、磷使得SOC礦化累積顯著增加。隨著培養時間延長,SOC礦化量逐漸增加。在培養前期,各樣地不同添加處理的SOC礦化沒有明顯差距。在培養中期,SOC礦化累積迅速增加。在培養后期,CK組SOC礦化累積達598.7、373.5、538.33 mg/kg,添加氮、磷使得礦化累積增加了211.0%~876.5%,H1、H2樣地N2組礦化累積略微降低,不同處理H1的礦化累積的大小為P2>N3>N2>P1>N1>NP>CK,H2為N3>P2>N2>P1>N1>NP>CK,H3為N2>P2>N3>NP>P1>N1>CK,N2、P2和N3組表現出明顯優勢。
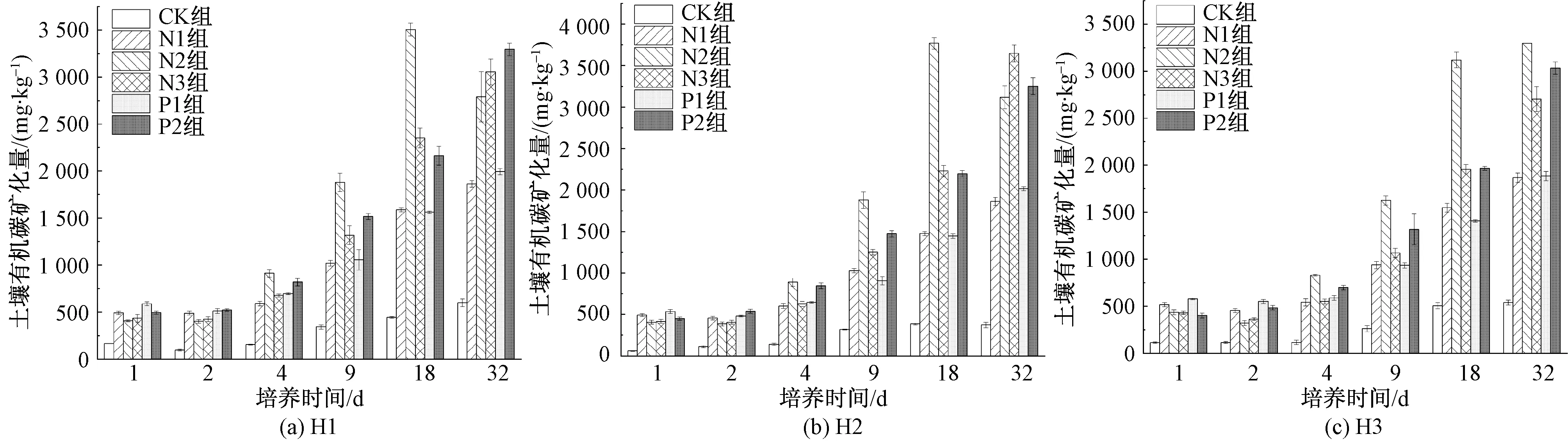
圖3 培養期間不同處理組SOC礦化累積Fig.3 Changes of SOC mineralization in different groups during incubation
2.4 不同處理對SOC礦化速率的影響
培養期間不同處理SOC礦化速率變化規律如圖4所示。添加氮、磷強烈刺激了SOC的礦化速率,培養0~1 d,CK組的礦化速率為164.8、60.4、112.6 mg/(kg·d),添加組礦化速率增加了146.7%~786.3%。SOC礦化速率在培養前期迅速下降,P1組的礦化速率和斜率最大[圖4(e)],但P2組在第2天略微升高[圖4(f)],N2組隨培養時間延長速率曲線略微波動[圖4(c)]。隨著培養時間延長,各組SOC礦化速率逐漸下降趨于平緩,培養結束后,CK組的礦化速率降至42.8、26.7、38.4 mg/(kg·d),各樣地礦化速率仍增加了211.0%~770.5%。
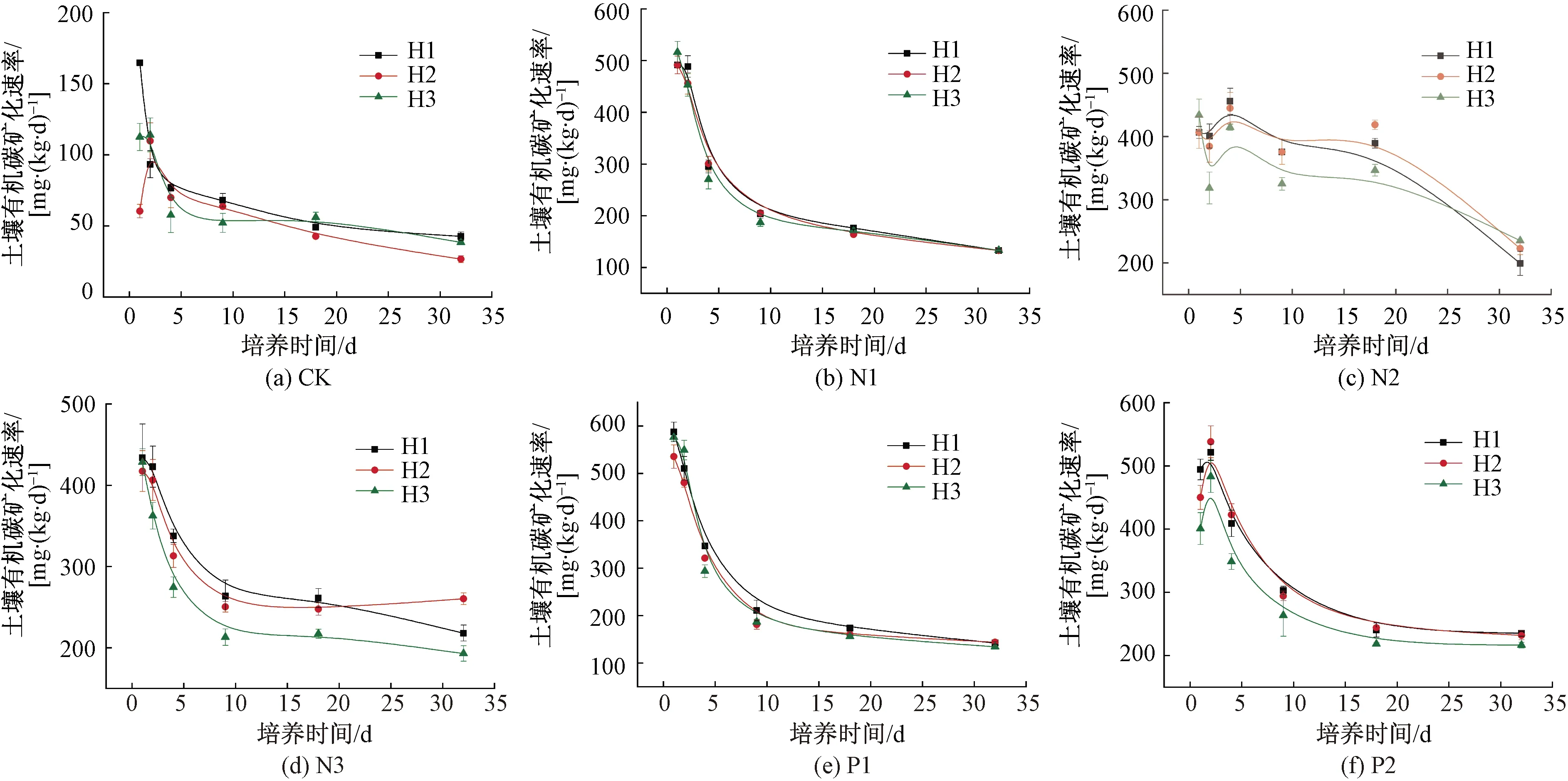
圖4 培養期間不同處理組SOC礦化速率Fig.4 Changes of SOC mineralization rate in different groups during incubation
2.5 不同處理對激發效應的影響
通過定量化SOC礦化速率的大小和方向,得到激發效應的變化規律如圖5所示。在培養期間,各樣地不同添加的激發效應均產生了正激發效應。與培養前期相比,N1、P1組激發效應在培養后期略微減弱[圖5(a),圖5(d)],其他組略微升高無明顯趨勢。培養結束后,各處理組激發效應大小均為:H2>H3>H1。在培養期間內,各樣地激發效應大小均為,N2組最大。說明添加N2對SOC礦化的激發效應響應更明顯。
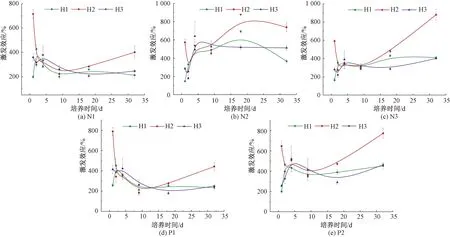
圖5 培養期間不同處理組激發效應的變化Fig.5 Changes of priming effect of different treatments during incubation
各樣地在不同添加下的激發效應變化規律相似,H1樣地在培養的0~4 d迅速上升后趨于平緩;H2樣地在培養的0~4 d迅速下降,隨著培養時間延長,激發效應逐漸增加;H3樣地在0~1 d略微下降,隨培養時間延長逐漸上升趨于平緩。
2.6 鹽堿程度對SOC礦化和激發效應的影響
通過相關性分析,研究原生土壤鹽堿程度對SOC礦化作用和激發效應的影響,培養期間不同處理組SOC礦化作用與鹽堿程度的顯著相關性結果如表2所示,激發效應與鹽堿程度的顯著相關性結果如表3所示。
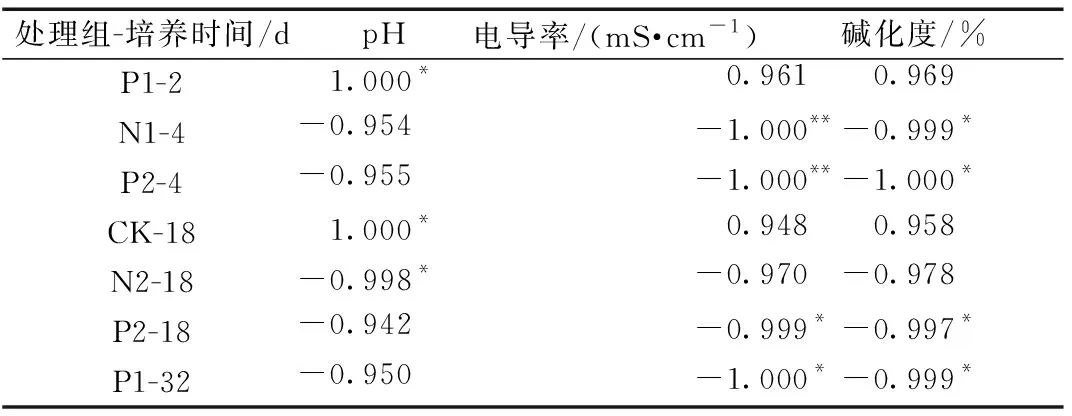
表2 各處理組SOC礦化與鹽堿程度的顯著相關性結果Table 2 The significance correlation between the SOC mineralization and soil salinization in different treatment groups

表3 各處理組激發效應與鹽堿程度的顯著相關性結果Table 3 The significance correlation between priming effect and soil salinization in different treatment groups
培養2~4 d,N1、P2組SOC礦化作用與EC、ESP顯著負相關(P<0.05、P<0.01);在培養的9~18 d,N2組SOC礦化作用與pH顯著負相關(P<0.05),P2組SOC礦化作用與EC、ESP顯著負相關(P<0.05);在培養的18~32 d,P1組SOC礦化作用與ESP顯著負相關(P<0.05)。激發效應與土壤鹽堿程度的顯著相關性結果集中于培養的9~18 d,N2、P2組激發效應與pH呈顯著負相關(P<0.05、P<0.01),N3、P1組激發效應與EC、ESP呈顯著負相關(P<0.05)。
3 討論
近年來,隨著人口增長,糧食需求大大增加。氮和磷是農作物生長的必要元素,為了提高作物產量,過量施用氮肥、磷肥的現象普遍存在。有研究發現,過量施用氮肥會刺激SOC產生大量礦化,導致SOC損失[26]。在本研究中,添加氮使得SOC礦化累積增加了2~8倍,這是因為氮的可利用性被增強,受氮限制的微生物活性提高,使得SOC礦化累積增加[26]。當前大多數研究認為,土壤缺磷會導致土壤礦化能力顯著降低,添加磷有利于SOC礦化[27],這與本研究的結果一致。在本研究中,添加磷使得SOC礦化累積增加了2~7倍,這是由于添加磷使得土壤中磷的含量增加,可溶性有機碳含量提高,SOC礦化累積增加[26, 28]。添加氮比添加磷對SOC礦化累積的刺激更大,這是因為添加氮使得堿性土壤pH降低,改善了土壤環境,促進了有機碳的分解[19];此外,研究區土壤含有大量的碳酸鹽(如Na2CO3、CaCO3等),磷添加后容易形成難溶的三鈣磷酸鹽,導致磷釋放緩慢[27],對有機碳礦化的響應減弱。
有研究認為,少量氨態氮的輸入會促進SOC礦化累積,而高濃度的氨態氮輸入會抑制SOC的礦化累積[29]。在本研究中,高濃度的氮輸入并沒有抑制SOC礦化作用,雖然N2組礦化累積量略有降低,但這是由于高濃度的氮和硫酸根離子對微生物活性有抑制作用,在一定程度上削減了SOC礦化累積;此外,由于研究區土壤堿化程度高,在培養后期,(NH4)2SO4發生硝化反應,造成氮素損失,SOC>礦化累積略微降低。
在本研究中,各樣地SOC礦化速率大體呈先快速降低,后緩慢下降趨于平緩的變化規律。在培養前期,土壤中易分解的組分含量較高,大量營養物質迅速釋放,提高了土壤中微生物活性[30],SOC產生正激發效應,礦化速率增加;隨著培養時間的延長,微生物開始利用惰性有機碳,CO2排放量減少,礦化速率隨之減緩[31];在培養后期,活性有機碳消耗殆盡[32],礦化速率趨于平緩。
有研究表明,貧瘠的土壤更易產生激發效應[19]。當土壤氮、磷含量較高時,SOC礦化的激發效應增大[33-34]。在本研究中添加氮、磷產生了正激發效應,SOC礦化速率顯著提高(圖4、圖5)。原因可能是:一方面,氮、磷作為作物生長的必要元素,添加氮、磷刺激了SOC分解,導致正激發效應明顯;另一方面,由于研究區土壤鹽堿程度大,有機碳含量低,一些微生物在維持“代謝警覺”的細胞狀態方面投入的能量很低,它們對底物的反應比對休眠細胞的反應更快,添加營養物使細菌的更替加快,從而觸發了正激發效應[35],SOC礦化速率增強。
水分通過影響微生物活性和微生物與有機質的有效接觸,進而影響土壤SOC礦化作用[36]。N2和P2組礦化速率略微波動,這可能是由于土樣重新加水后,增加了土壤通氣性,細胞破裂,滲透物釋放,SOC產生了激發效應,有機碳礦化速率短暫增強[37]。
已有研究證明,鹽堿會抑制SOC礦化作用[38]。在本研究中,鹽堿程度、SOC礦化作用和激發效應表現出顯著負相關(P<0.05)。原因如下:①土壤鹽堿程度越強,土壤表面越光滑(圖3),土壤表面粗糙程度與吸附能力強弱有一定相關性[39],在一定程度上影響了土壤對氮、磷的吸附;②土壤有機質是土壤團聚化作用的主要影響因子,而研究區土壤鹽堿程度大,土壤有機質含量低,土壤結構較差,不利于土壤團聚體的形成,阻礙了氮、磷水溶液與土壤的細小顆粒結合形成團聚體[40-41],SOC礦化作用減弱;③大量研究表明,pH是影響微生物群落結構的重要因子,pH過高或過低都會抑制微生物的活性[42],而EC過高可能對SOC礦化、微生物種群產生負面影響[43]。綜上所述,當鹽堿程度較高時(pH>8.5,EC>1.81 mS/cm,ESP>20%),會破壞土壤的物理性,土壤結構較差,微生物活性受到抑制,SOC礦化作用減弱。
添加氮、磷會提高鹽堿土壤pH,改善土壤環境,增加SOC礦化作用,但大量施用氮肥和磷肥,會加速SOC的分解,導致土壤肥力降低,溫室氣體過度釋放。因此,對于SOC含量較低、鹽堿化程度較高的土壤CO2排放問題應被關注。
4 結論
(1)不同氮、磷添加處理,均促進了鹽堿旱田SOC礦化累積,且同一添加物濃度越高,礦化累積越大,N2、P2和N3對SOC礦化作用的刺激更大。
(2)在培養前期,添加氮、磷使得鹽堿旱田SOC礦化速率大大增加,正激發效應劇烈變化,隨著培養時間延長,各樣地礦化速率迅速降低后趨于平緩,礦化速率增幅降低,正激發效應趨于平緩。
(3)在不同培養階段,不同處理組的SOC礦化、激發與鹽堿程度顯著負相關(P<0.05),pH、EC和ESP越大,SOC礦化作用越小,正激發效應越弱。
(4)添加氮肥和磷肥,改善了鹽堿旱田的土壤環境,為作物生長提供了更多的養分,但過度施用氮肥和磷肥,會加速SOC礦化作用,導致SOC儲量降低,土壤肥力下降,溫室效應加劇。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
核科學與工程(2021年4期)2022-01-12 06:30:26
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中學物理·高中(2016年12期)2017-04-22 11:53:03
