麋鹿鹿角脫落、群主更替、產仔的年節律及其環境影響因子
2020-11-12 07:52:40程志斌劉定震白加德鐘震宇林潤生田東曉王麗斌
生態學報 2020年18期
程志斌, 劉定震, 白加德,*, 鐘震宇, 林潤生, 田東曉, 王麗斌, 劉 田
1 北京麋鹿生態實驗中心, 北京 100076 2 北京師范大學生命科學學院, 教育部生物多樣性與生態工程重點實驗室, 北京 100875 3 北京市氣象信息中心, 北京 100089
地球上的光、溫度、濕度等生態因子具有明顯的周期性,因而在生物的進化和發展中形成了晝夜節律、月節律、年節律等生物節律[1-3]。動物的年節律如繁殖、脫角、換羽(毛)、遷徙、冬眠等行為是一種復雜的生物學現象,它是動物對光照、溫度、濕度、水等非生物條件,食物、種內關系和種間關系等生物條件的適應,每個物種均有各自的生物節律特點,來適應外界復雜而變動的外界環境條件[4-8]。因此,通過動物的生物節律來研究動物與環境之間的適應性關系始終是生態學家和物候學家的研究熱點[9-12]。繁殖活動是哺乳動物生命中最為重要的組成部分,已有大量關于鳥類[13-15]和獸類[16-18]繁殖年節律會著溫度、降水等環境變化而產生適應性變化的報道,有蹄類作為哺乳動物中的重要類群,其繁殖的年節律受光周期、氣候、營養、緯度、種群密度等環境因子的影響[19-20]。然而結合具體的平均氣溫和降水量來探討氣候對哺乳動物繁殖特征年節律影響方面的報道較少。
各物種采取的繁殖策略是多種多樣的,包括從高度季節性到完全無季節性的繁殖模式[21]。生活在高緯度地區的鹿科動物屬于前者,雄鹿每年的繁殖周期包含發情期、身體恢復期、鹿角脫落期和長茸骨化期四個階段,雌鹿每年的繁殖周期包含發情期、懷孕期、產仔期和哺乳期四個階段[22]。首先,鹿角是鹿亞科(Cervinae)和空齒鹿亞科(Odocoileinae)動物特有的性二型特征[23],是繁殖期雄性爭奪交配權和雌性選擇雄性的重要指標[24-25],它是哺乳動物唯一可再生的器官[23],高緯度地區鹿科動物的鹿角每年周期性脫換[23,26]。其次,占群繁殖,是一雄多雌的哺乳動物雄性繁殖期的重要行為特征之一,如川金絲猴(Rhinopithecusroxellana)[27]、象海豹(MiroungaLeonina)[28]、藏羚羊(Pantholopshodgsonii)[29]、普氏野馬(Equusprzewalskii)[30]等。第三,雌性鹿類動物的每次的發情持續時間較短,通常只有幾個小時[31],不易于觀察,因此更為直觀的產仔周期成為研究雌鹿的繁殖年節律特征的首選[17,32]。
盡管關于鹿類動物年節律特征的研究有許多,但主要為單一或者兩個繁殖特征,如馬鹿[17, 33-34](Cervuselaphus)、狍[35](Capreoluspygargus)、馴鹿[36](Rangifertarandus)等廣布物種的角生長、發情或者產仔的年節律,以及白尾鹿(Odocoileusvirginianus)血液中激素的年節律變化[37-38]。而同一群體的多個繁殖特征的變化差異、相互之間的關系及其環境影響因子等方面的研究甚少。麋鹿(Elaphurusdavidianus)為我國特有物種,國家一級保護動物,IUCN紅皮書列為野外滅絕,屬于典型的季節性繁殖動物,雄性具角,具有自身獨特的脫角周期[23]。麋鹿的婚配制度為典型的一雄多雌的后宮制[39-40],雄性麋鹿采用序列占群的繁殖策略[40],通過頂角爭斗獲得群主地位和交配權[41-43],目前尚未見到麋鹿年節律研究的相關報道。本文通過研究麋鹿鹿角脫落、群主更替、產仔等繁殖特征的年節律及其環境影響因子,為深入了解哺乳動物在進化過程中對環境的適應性,以及為保護麋鹿這一珍稀物種提供科學依據。
1 材料與方法
1.1 研究地概況
研究地為北京南海子麋鹿苑(39°50′N, 116°30′E),位于北京城南10 km,占地60 hm2,麋鹿半散放活動的保護區面積約40 hm2,為古代元明清皇家苑囿南海子的一部分,是麋鹿模式種產地、我國最后滅絕地和我國第一處重引入地。苑內優勢植物有牛筋草(Eleusineindica),大畫眉草(Eragrostiscilianensis),馬唐(Digitariasanguinalis)和狗尾草(Setariaviridis)等;年均氣溫為13.1℃,1月平均氣溫為-3.4℃,7月平均氣溫為26.4℃;年降雨量約為600 mm[44]。苑內無河流經過,為人工湖,水域靠降雨和抽取地下水補給。
北京南海子麋鹿種群是麋鹿在國內滅絕后建立的第一個半散放遷地種群。1985年和1987年北京南海子麋鹿苑從英國烏邦寺共引入38只麋鹿。回歸35年來,北京南海子麋鹿苑分別向湖北石首、江西鄱陽湖、浙江慈溪、河北灤河上游等地輸送536只麋鹿,建立了39個遷地保護種群,在長江中下游建立了麋鹿野生種群。北京南海子麋鹿種群維持在130—210只之間(雄雌性比約1∶1),采取半散放的飼養方式,由于夏秋兩季地面草本植物生物量有限,一年四季均有人工補飼。
1.2 數據采集及處理
1.2.1鹿角脫落數據采集及處理
2014—2016年,每一年度掉角期12月—翌年2月,每天上午和下午各一次在麋鹿生活區域巡視,收集麋鹿自然脫落的角,且在角上標記脫落日期。選擇鹿角脫落開始時間、鹿角脫落高峰期、鹿角脫落結束時間作為鹿角脫落年節律因子。
脫角數量達到半數為脫角中位值日期[45],由于脫角中位值日期處于脫角集中時間段,為了便于討論,將其定義為鹿角脫落高峰期;由于2歲齡鹿角(稱筆桿角,無分叉)在脫落后掉于草叢中難以收集,因此本文將2叉(3歲齡)及2叉以上的鹿角全部脫落完畢的日期定義為鹿角脫落結束時間。
鹿角回收率(%)=(鹿角回收數/2歲以上雄性數×2)×100%
1.2.2群主更替數據采集及處理
2015—2017年,每年發情期5—9月,一周觀察4—7 d,每天觀察0.5—2.5 h,使用雙筒望遠鏡(SWAROVSKI 8×42WB)、相機(Canon550)記錄群主的特征,通過鹿角形狀、耳標、耳缺、項圈、體型等區分個體和年齡。群主為占有和控制發情雌鹿群(后宮)的等級序位高的雄鹿[41]。群主有兩種方式控制發情雌鹿群:一是直接控制發情雌鹿群,二是控制發情場[40]。由于北京南海子麋鹿為半散放種群,發情場地有限,在水邊泥灘的高大喬木下,占有發情場地是群主最喜歡的方式。本研究中群主為占有固定發情場的群主,而占有臨時雌性群(指離開固定發情場外出覓食形成的臨時雌性群,它們通常1 h內會被群主驅趕回固定發情場或者被群主控制)的臨時群主不計算在內。
將第一只群主開始占有和控制雌鹿群,且占有固定發情場的日期定義為第一次發情期開始時間(即群體而言雄性發情開始時間)[17];第一次發情期間群主更替次數達到半數的日期為群主更替中位值日期[46],由于群主更替中位值日期處于群主更替集中時間段,為了便于討論,將其定義為群主更替高峰期;對于整個種群而言,將當年第一次發情期的群主占群更替結束的日期定義為第一次發情期結束時間(通常群主全身為夏毛,未開始更換冬毛);將當年第二次發情期的群主開始占群的日期定義為第二次發情期開始時間(通常群主夏毛已經脫落完畢,正在長冬毛);將當年第二次發情期的群主占群更替結束的日期定義為第二次發情期結束時間(即群體而言雄性發情結束時間)。
選擇第一次發情期開始時間、群主更替高峰期、第一次發情期結束時間、第二次發情期開始時間、第二次發情期結束時間作為群主更替年節律因子。
1.2.3產仔數據采集及處理
2016—2018年,每一年產仔期3—7月,每天使用望遠鏡觀察麋鹿群,尤其是離群即將分娩或者正在分娩的母鹿,上午和下午各觀察鹿群及其活動場所一次,做好新生仔鹿出生記錄。選擇產仔期開始時間、產仔高峰期、產仔期結束時間作為產仔的年節律因子。
產仔數量達到半數的日期為產仔中位值日期[47],由于產仔中位值日期處于產仔集中時間段,為了便于討論,將其定義為產仔高峰期。
1.2.4環境數據采集及處理
2014年1月—2018年7月氣象數據由北京市氣象信息中心提供,站點為大興站(站號54594,國家一般氣象站),距離研究地點10.3 km;2018年7—12月的氣象數據取自中國地面國際交換站氣候資料日值數據集,站點為北京站(站號54511,國家基本氣象站),距離研究地點3.7 km。選擇氣溫(3—9月、12月的月平均氣溫,春、夏、秋、冬季4個季節的季平均氣溫,年平均氣溫)、降水(春季第一場雨、春季第一場透雨、雨季開始時間、雨季結束時間、年降雨量)作為氣候因子。
春季第一場透雨參照姚佩珍和張強[48]北京春季第一場透雨指標的方法,由于2014年、2015年、2017年、2018年春季降雨量均超過70 mm,降水充沛,第一場透雨日的標準按照降雨量超過或者約等于當旬輕旱的平均標準值統計。雨季開始和結束時間參照于曉澄等[49]華北雨季開始早晚的方法。
由于麋鹿半散放區域的面積不變,以年末種群數量作為種群密度因子。
1.3 作圖
按照實際日期制作每一年度麋鹿鹿角脫落、群主更替、產仔和日降水量情況變化圖。日降水量存在的缺失值在圖中不顯示。
2 結果
2.1 麋鹿鹿角脫落、群主更替、產仔的年節律
2.1.1鹿角脫落的年節律
由表1可知,2014—2016年麋鹿角的回收率為81.37%—90.70%,說明可根據實際所收集到的麋鹿角脫落日期統計鹿角脫落高峰期。圖1表明,整個種群而言,麋鹿角脫落的年節律存在差異,2014—2016年的鹿角脫落開始時間分別為12月16日、12月7日、12月3日,鹿角脫落高峰期分別為12月31日、12月26日、12月21日,鹿角脫落結束時間分別為2月7日、2月4日、1月17日。說明北京南海子麋鹿種群鹿角脫落時間開始于12月,結束于1月下旬或2月上旬;2015年、2016年的鹿角脫落開始時間、鹿角脫落高峰期、鹿角脫落結束時間與前一年比較均出現同步提前的現象。
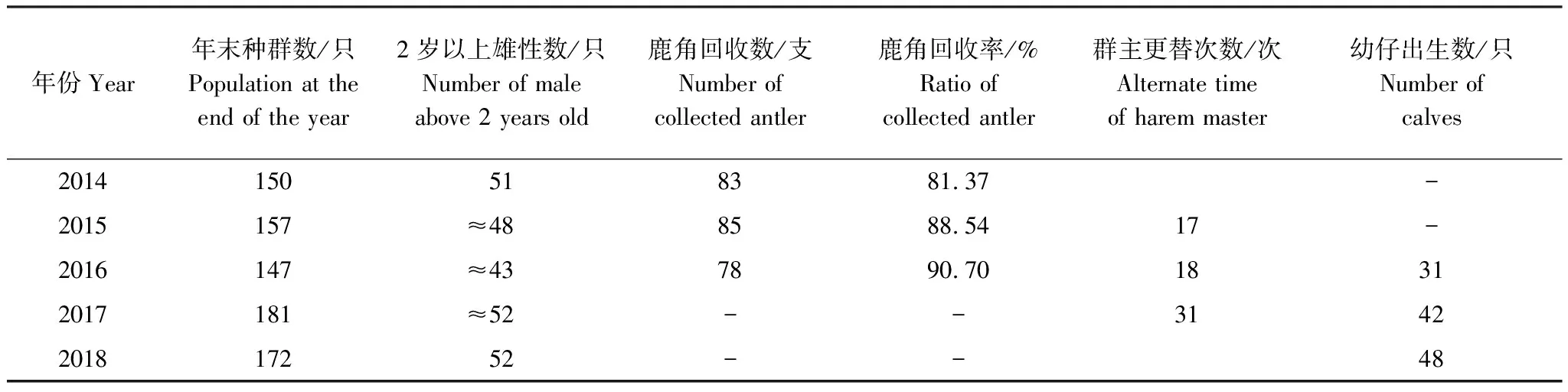
表1 北京南海子麋鹿苑2014—2018年麋鹿年末存欄、脫角、群主更替次數、產仔情況
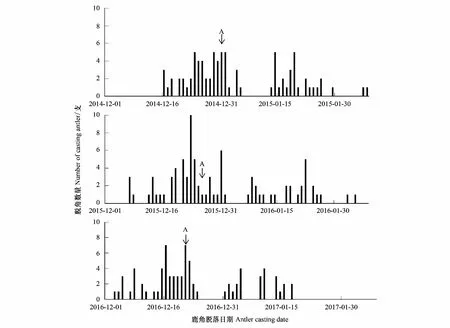
圖1 北京南海子麋鹿苑2014—2016年度麋鹿角脫落情況
2.1.2群主更替的年節律
圖2表明,雄性麋鹿存在第二次發情期的現象;2017年出現兩個固定發情場所,其中第二個固定發情場6月7日出現,6月28日消失。整個種群而言,群主更替存在年節律的差異,2015—2017年的第一次發情期開始時間分別為6月9日、5月30日、5月17日,群主更替高峰期分別為7月22日、7月4日、6月28日,第一次發情期結束時間分別為9月1日、8月12日、8月15日,第二次發情期開始的時間分別為9月1日、8月15日、8月17日,第二次發情期結束時間分別為9月8日、9月9日、9月4日;2015—2017年發情期時間分別為91d、102d、110d。說明北京南海子麋鹿苑麋鹿發情期開始于5月下旬或6月上旬,結束于9月上旬;2016年和2017年的第一次發情期開始時間、群主更替高峰期與前一年比較均出現同步提前的現象;而第一次發情期結束時間、第二次發情期開始時間和第二次發情期結束時間不存在同步提前的現象;從2015年至2017年,整個鹿群發情的時間有延長的趨勢。
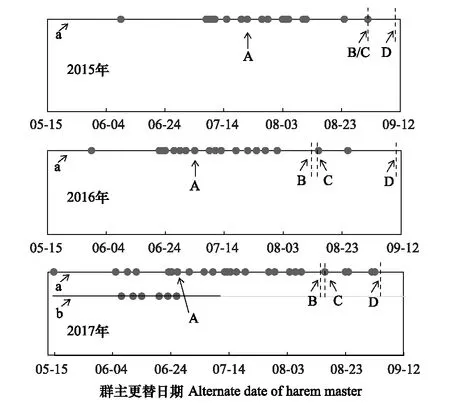
圖2 北京南海子麋鹿苑2015—2017年麋鹿群主更替情況
2.1.3產仔的年節律
由表1和圖3可知,整個種群而言,麋鹿產仔年節律存在差異,產仔期開始時間2016—2018年分別為4月12日、3月28日、3月16日;產仔高峰期分別為4月23日、4月17日、4月3日,產仔期結束時間分別為7月25日、7月15日、5月25日;產仔期分別長達104 d、109 d、70 d。2016年70.97%的幼仔集中在高峰期前后的13 d內出生,2017年64.29%的幼仔集中在高峰期前后的21 d內出生,2018年56.25%的幼仔集中在高峰期前后的19 d內出生。說明北京南海子麋鹿苑麋鹿產仔期開始于3月中旬至4月中旬,結束于5月下旬至7月下旬;2017年和2018年產仔期開始時間、產仔高峰期、產仔期結束時間與前一年比較均出現同步提前的現象;麋鹿產仔同期化變弱。
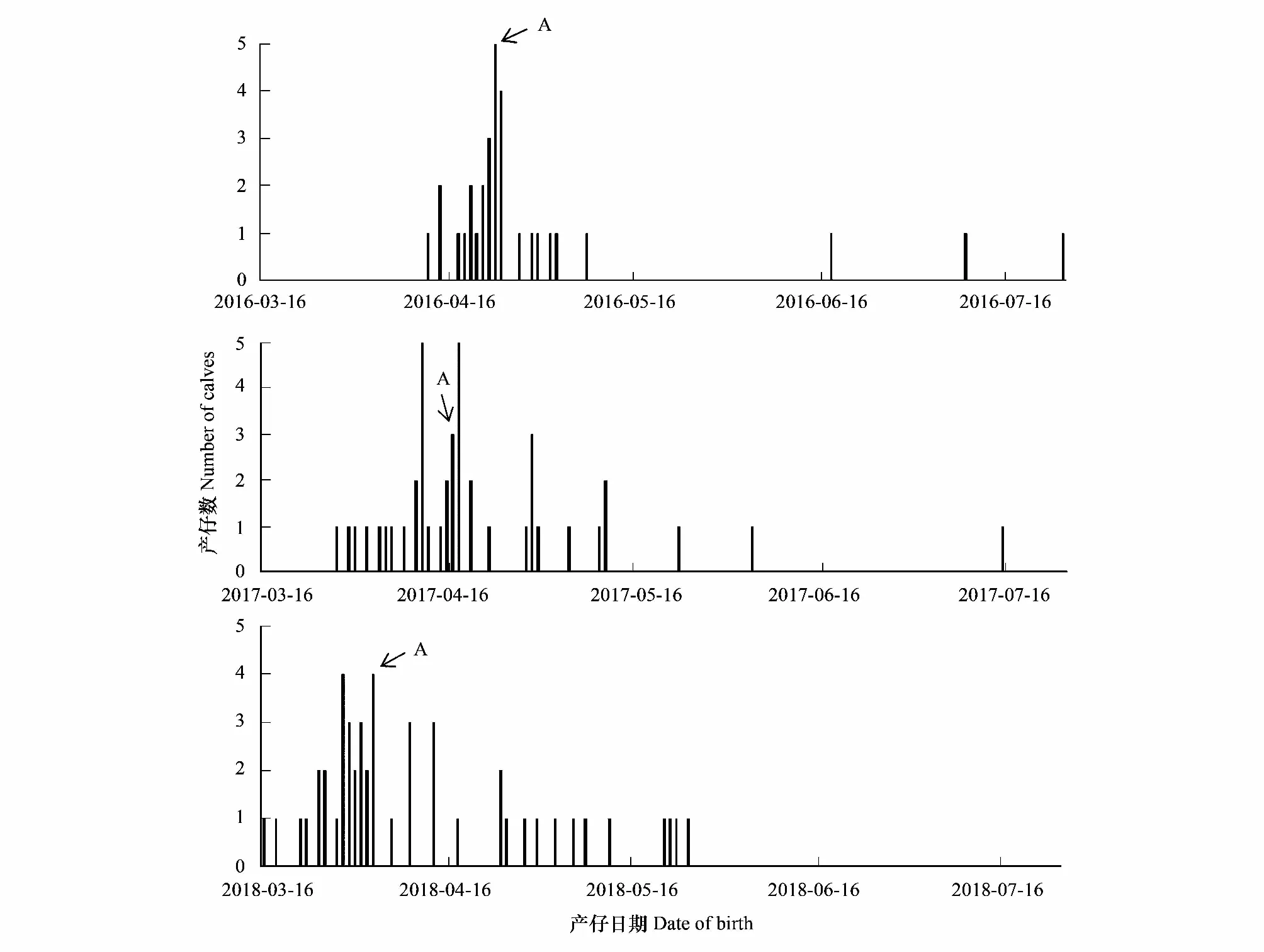
圖3 北京南海子麋鹿苑2016—2018年度麋鹿產仔情況
2.2 環境因子
由表1可知,北京南海子麋鹿苑2014—2018年麋鹿年末種群數量分別為150只、157只、147只、181只、172只。
2014—2018年的月平均氣溫見圖4;年平均氣溫分別13.88℃、13.62℃、13.36℃、13.65℃、13.34℃;春季平均氣溫分別為16.42℃、15.54℃、15.40℃、16.27℃、15.04℃,夏季平均氣溫分別為26.30℃、26.17℃、26.37℃、26.08℃、27.51℃,秋季平均氣溫分別為13.56℃、12.77℃、12.89℃、12.57℃、13.36℃。2014—2017年冬季(12月、翌年1—2月)平均氣溫分別為-0.13℃、-1.36℃、-0.11℃、-2.00 ℃。
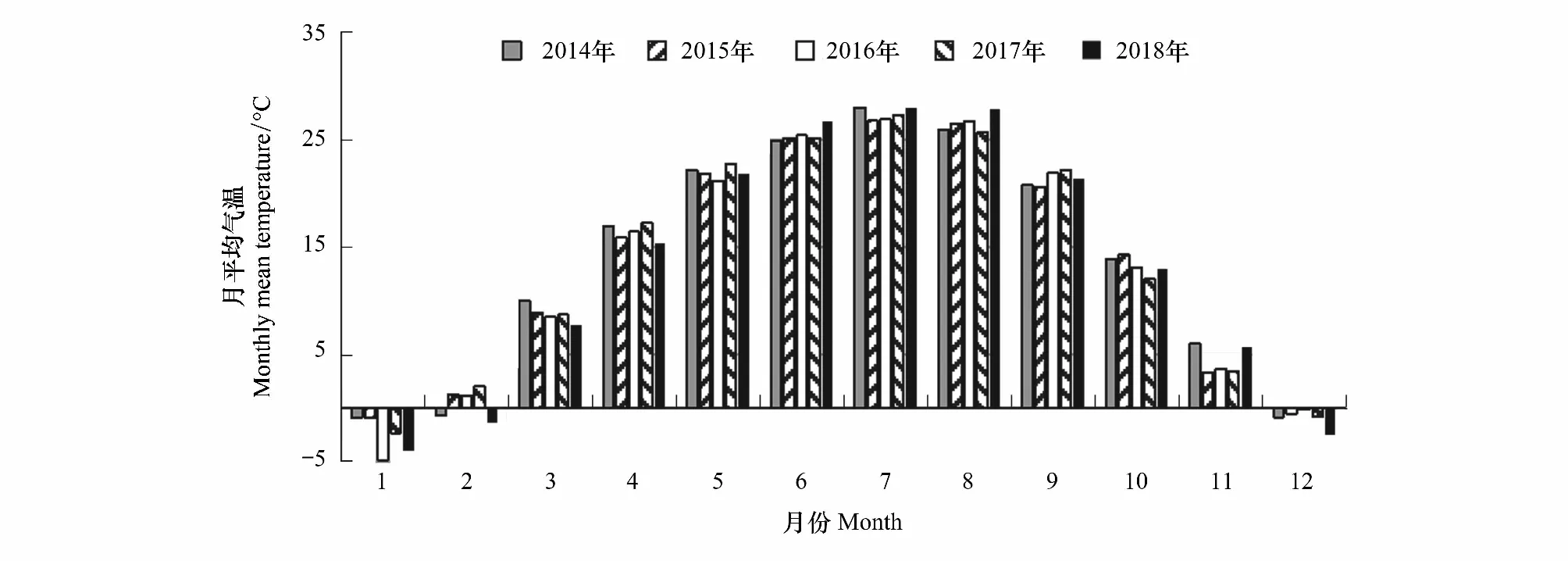
圖4 北京南海子麋鹿苑2014—2018年月平均氣溫
2016年春季 3—5月共7場降雨,降雨量總和為30.7 mm,屬于干旱,7場降雨均沒有達到春季第一場透雨的標準。2014—2018年的年降雨量分別416.8 mm、505.8 mm、722.7 mm、682.9 mm、537.1 mm。由圖5可知,2014—2018年的第一場春雨分別為3月28日、3月31日、4月2日、3月20日、3月17日;2014年、2015年、2017年、2018年的春季第一場透雨分別為4月17日、3月31日、3月23日、4月5日;2014—2018年的雨季開始時間分別為7月2日、7月17日、7月19日、7月4日、7月8日;2014—2018年的雨季結束時間分別8月13日、8月31日、7月28日、8月27日、8月13日。
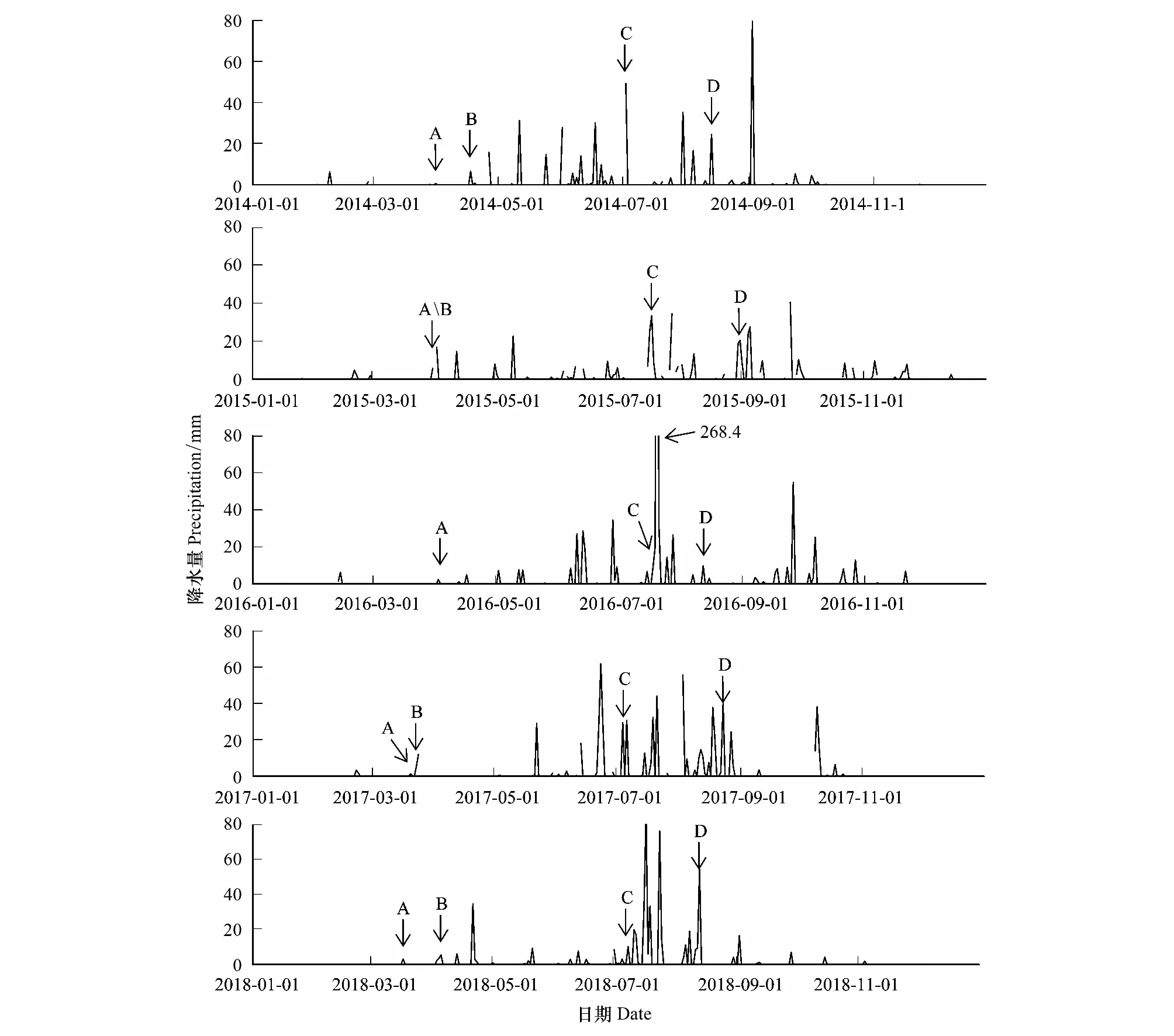
圖5 北京南海子麋鹿苑2014—2018年日降水量
2.3 鹿角脫落、群主更替、產仔的年節律之間的同步關系
由圖1—3可知,當2014—2016年麋鹿鹿角脫落開始時間、鹿角脫落高峰期和鹿角脫落結束時間逐年提前時,2015—2017年第一次發情期開始時間、群主更替高峰期也隨之逐年提前;2016—2018年的產仔期開始時間、產仔高峰期和產仔期結束時間也緊接著逐年提前。2015—2017年的第一次發情期開始時間至翌年產仔期開始時間間隔分別為308 d、302 d、303 d,平均間隔為304.3 d;2015—2017年的群主更替高峰期至翌年產仔高峰期間隔分別為276 d、287 d、279 d,平均間隔為280.7 d。說明2014—2016年的麋鹿鹿角脫落年節律因子(鹿角脫落開始時間、鹿角脫落高峰期和鹿角脫落結束期)、2015—2017年群主更替年節律因子(第一次發情期開始時間和群主更替高峰期)、2016—2018年的產仔年節律因子(產仔期開始時間、產仔高峰期和產仔期結束時間)存在明顯的同步關系。
2.4 鹿角脫落、群主更替、產仔的年節律與環境因子之間的關系
圖1—5可知,麋鹿鹿角脫落的年節律與秋季平均氣溫、冬季平均氣溫、年平均氣溫不存在隨著平均氣溫升高或降低而提前或者延遲的現象;2015—2017年群主更替的年節律與2014—2016年的年平均氣溫和秋季平均氣溫,與本年度夏季平均氣溫不存在隨著平均氣溫升高或降低而提前或者延遲的現象;2016—2018年產仔的年節律與2015—2017年秋季平均氣溫、冬季平均氣溫和年平均氣溫,與本年度春季平均氣溫不存隨著平均氣溫升高或降低而提前或者延遲的現象。產仔期開始時間與3月平均氣溫,產仔期開始時間和產仔高峰期與4月平均氣溫,第一次發情期開始時間與5月平均氣溫,第一次發情期開始時間和群主更替高峰期與6月平均氣溫,群主更替高峰期與7月平均氣溫,第一次發情期結束時間和第二次發情期開始時間與8月平均氣溫,第二次發情期結束時間與9月平均氣溫均不存在隨著平均氣溫升高或降低而提前或者延遲的現象。但鹿角脫落開始時間、鹿角脫落高峰期和鹿角脫落結束時間存在隨著12月平均氣溫升高而提前的現象;產仔期開始時間和產仔高峰期存在隨著前一年9月平均氣溫的升高而提前的現象。說明除群主更替年節律外,產仔年節律和鹿角脫落年節律表現出受氣溫因子影響的現象。
鹿角脫落開始時間、鹿角脫落高峰期和鹿角脫落結束時間均與年降雨量存在隨著年降雨量的增加而提前的現象。2015—2017年第一次發情期開始時間、群主更替高峰期的年節律存在隨著2014—2016年年降雨量的增加而提前的現象。2016—2018年產仔期開始時間和產仔高峰期不存在隨著2015—2017年年降雨量的增加或減少而提前或延遲的現象。產仔期開始時間、產仔高峰期和第一次發情期開始時間均不存在隨著春季第一場雨、春季第一場透雨的提前或者延遲而提前或者延遲的現象。群主更替高峰期不存在隨著雨季開始的提前或者延遲而提前或者延遲的現象;第一次發情期結束時間、第二次發情期開始時間和第二次發情期結束時間均不存在隨著雨季開始時間、雨季結束時間的提前或者延遲而提前或者延遲的現象。2015年的群主更替高峰期出現在雨季期間,2016年和2017年的群主更替高峰期出現在雨季開始之前。說明,產仔年節律沒有表現出與降水因子的變化而出現相應的同步變化,鹿角脫落年節律表現出隨著年降雨量的增多而提前的現象;第一次發情期開始時間、群主更替高峰期的年節律表現出隨著前一年度年降雨量的增多而提前的現象。
由圖1—3和表1可知,麋鹿鹿角脫落年的節律、群主更替的年節律、產仔的年節律均不存在隨著種群密度升高或降低而提前或者延遲的現象。
3 討論
3.1 麋鹿鹿角脫落、群主更替、產仔的年節律
高緯度地區的有蹄類,均具有各自的繁殖節律特征來適應高度季節性的生境。本研究中,北京南海子麋鹿種群鹿角脫落時間從12月開始,1月下旬或2月上旬結束,這與極地物種馴鹿、歐亞大陸高緯度地區的廣布物種狍的鹿角脫落時間相一致,而馬鹿、梅花鹿(Cervusnippon)、黇鹿(Damadama)等大部分鹿科動物為春夏之際開始脫角[50]。有蹄類喜歡選擇在春夏之際氣候和食物條件最適的時候集中產仔,研究顯示麋鹿產仔期為3—7月,集中于4—5月,這與溫帶地區和寒帶地區的其他鹿類相一致[50]。本研究表明,麋鹿發情期為5月下旬或6月上旬開始,9月上旬結束,麋鹿存在二次發情期現象;這與李春旺等[42]研究結果相同。本研究發現群主更替高峰期與翌年產仔高峰期間隔平均為280.7 d,而人工圈養的麋鹿妊娠期平均為280 d[51],因此可以根據前一年的群主更替高峰期推測第二年的產仔高峰期。有研究表明,黑尾鹿(Odocoileushemionus)發情高峰期可以預測翌年產仔時間的情況[52];可通過馬鹿鹿角脫落年節律來預測緊接著的雄性發情年節律[17]。
高緯度地區鹿類動物的鹿角脫落、發情、產仔等繁殖周期具有同步性[53]。雄鹿為了提高等級序位,增加自身繁殖后代的貢獻率,可通過使角提前骨化而先進入發情占群期[46]。南非一些反芻動物在反捕食策略上表現為產仔行為和交配行為同步[54]。本研究發現麋鹿鹿角脫落、群主更替、產仔的年節律存在明顯的同步關系,其中鹿角脫落開始時間、鹿角脫落高峰期、鹿角脫落結束時間、第一次發情期開始時間、群主更替高峰期、產仔期開始時間、產仔高峰期、產仔期結束時間與前一年度比較均出現同步提前的現象。這與馬鹿繁殖特征的年節律相似[33,55],Clements等[46]對野生馬鹿長達28年的研究表明,馬鹿群體鹿角脫落開始時間、鹿角脫落結束時間、雄性發情開始時間、雄性發情高峰期、雄性發情結束時間、雌性發情開始時間和產仔期開始時間等七個繁殖特征之間均呈顯著正相關關系,并呈線性逐年同步提前現象[17]。
3.2 環境影響因子
3.2.1氣候因子
光周期具有恒定性和規律性,隨著地理緯度和季節而很有規律地變動,它是引起動物繁殖活動年節律變化的直接原因[3, 19, 56],通過神經——垂體——性腺軸控制動物繁殖期[42, 57]。根據動物繁殖與光照長短的關系,可將動物分為長日照動物和短日照動物,羚羊類和鹿類只有在白晝逐步縮短的秋冬之際才開始性腺活動,它們屬于短日照動物[3],然而麋鹿是鹿類中唯一的夏季發情的短日照發情動物[42, 50, 53],這與本研究的結果相一致,麋鹿發情期為5月下旬—9月上旬,發情高峰期集中在光照時長最長的6—7月。通過適應光周期的變化,溫帶地區的鹿類動物產生繁殖年節律,它們可以優化調整產仔時間,使其與最適的環境氣候因子(如食物)相同步[58]。本研究表明,麋鹿的鹿角脫落、群主更替、產仔等繁殖特征存在明顯的年節律,恰好說明它們受光周期的影響,形成周期性變化的年節律。有研究表明,將南半球的普度鹿(Pudupudu)遷移至北半球,它的繁殖期會很快與當地的光周期同步[58]。
光在決定動物繁殖年節律中起著重要作用,但并不是說它是唯一的信號,溫度、濕度、降水等氣候原因對動物繁殖年節律也有重要的影響[3]。非洲的有蹄類如斑馬(Equusquagga)的產仔年節律同環境的氣溫、降水量的變動有關[59]。地中海馬鹿雌性發情開始時間在更加干旱和炎熱的春季會推遲;在更加涼爽和降雨量多的秋季會提前[60]。我們的研究中,麋鹿的鹿角脫落、群主更替、產仔等繁殖特征連續3個年度的年節律不是固定不變的,而是存在年際差異,說明它們受到其他因子的影響。
溫度可以影響動物的生活史特點,如使狍的產仔年節律提前[35]。氣候變暖伴隨著有效積溫的逐年升高,使得蘇格蘭Rum島植物的生長量增加,給當地馬鹿帶來更多的食物,導致鹿角脫落開始時間、鹿角脫落結束時間、產仔期開始時間、雄性發情開始時間、雄性發情結束時間呈逐年提前的趨勢[17, 46]。而且馬鹿鹿角開始脫落的時間與前一年秋季溫度有關,因為秋冬季發情結束之后的食物多少與秋季溫度密切相關[61],而且前一年9月和當年4月平均氣溫升高會使馬鹿鹿角脫落年節律提前[62]。我們的研究結果也顯示產仔期開始時間和產仔高峰期存在隨著前一年9月平均氣溫的升高而提前的現象,鹿角脫落年節律存在隨著每年12月平均氣溫升高而提前的現象。
降水是陸生動物的重要環境因子,降水會改變哺乳動物繁殖的年節律,本研究表明,麋鹿第一次發情期開始時間、群主更替高峰期的年節律表現出隨著前一年度年降雨量的增多而提前的現象。雌性馴鹿在4月降水量大的年份,表現出產仔期開始時間推遲[18];但是本研究中,產仔年節律未表現出隨著降水因子的變化而發生相應的同步變化。有研究認為,當冬季降雪量增多時,第二年馬鹿鹿角脫落開始時間會推遲[62-63],本研究中麋鹿鹿角脫落年節律表現出隨著年降雨量的增多而提前的現象。
3.2.2營養和種群密度因子
營養(或食物)也是動物繁殖年節律變化的重要原因[59]。植物提前生長或者增加生物量,可為食草動物在關鍵的繁殖期提供更多的食物[64-65]。增加或者提前的春季食物可使雌鹿妊娠期和雄鹿鹿角脫落時間提前;植物生長時間延長,可提高哺乳期和長茸期食物的質量,讓它們提前進入秋季的發情期[17]。動物能夠根據產仔的營養需要,調整產仔時間,將產仔時間與環境氣候最適時間(如食物供應高峰期)同步[35, 52, 63],提高幼崽存活率[32]。本研究中,2015—2017年麋鹿鹿群發情時間有延長的趨勢,2016—2018年麋鹿產仔同期化變弱,分析原因,可能與北京南海子麋鹿種群為半散放狀態,食物營養充分,食物壓力較小有關。圈養動物與野生動物的產仔年節律不同,受氣候和食物等因子的影響,圈養的馬麝(Moschuschrysogaster)產仔同期化比野生種群更弱[66]。白尾鹿的動物園種群產仔期開始時間為2月,而野生種群4月才開始產仔[25]。南美草原鹿(Ozotocerosbezoarticus),阿根廷干旱草原生境中的種群全年產仔,產仔80%集中在10月至翌年4月出生;而同緯度相鄰的烏拉圭半干旱大陸性氣候生境中的種群為季節性產仔,產仔同期化更加集中,高峰期推遲至夏季末或秋季初食物充沛的時期;而熱帶環境下的巴西種群,由于明顯的旱季和雨季,食物變化更為顯著,產仔同期化最顯著,產仔高峰期為8—9月,與當地的雨季完全同步[32]。但是,有學者認為,這一同步現象不是立即發生的[67-68],有時會存在時間誤差,對于狍而言它的胚胎滯育策略就顯得尤為重要[35]。
野外條件下食物和生存空間有限,種群密度大,意味著個體之間的競爭力大。地中海馬鹿雌性發情開始時間在種群密度大的年份會推遲;在種群密度小的年份會提前[60]。蘇格蘭Rum島馬鹿種群也存在高密度種群會推遲角脫落開始時間和角脫落結束時間的現象[62]。本研究表明,麋鹿鹿角脫落、群主更替、產仔的年節律均不存在隨著種群密度升高或降低而提前或延遲的現象,也許和半散放狀態下,每天人工補飼,不存在食物壓力有關。
3.2.3緯度
隨著緯度的變化鹿類動物繁殖年節律會隨之變化[52]。江蘇大豐麋鹿脫角開始時間和結束時間早于北京南海子種群[23]。本研究中,北京南海子麋鹿群主更替高峰期為6—7月,而低緯度地區的江蘇大豐麋鹿種群雄性發情高峰為提前為5—6月[42]。這與其他鹿類動物相似,墨西哥的白尾鹿發情開始時間為7月,而美國和加拿大地區為10月[25],高緯度和低緯度地區馬鹿發情和產仔年節律研究的結果也表現出這一特征[33]。然而,本研究發現麋鹿鹿角脫落開始時間、發情開始時間、產仔期開始時間逐年提前,而江蘇大豐麋鹿種群2004—2010年鹿角脫落開始時間、發情開始時間、產仔期開始時間出現了延后的現象[69],與本研究結果相反,也許與江蘇大豐為濱海濕地生境,而北京南海子為內陸濕地生境有關。
種群繁殖特征年節律是一個復雜的規律,受氣候、營養、種群密度等諸多因子影響,甚至種群內個體間的相互作用及各飼養區的管理模式也會對它造成影響[17, 60, 66]。本研究的數據為3個年度3種繁殖特征的年節律,時間相對較短,對麋鹿繁殖特征與環境因子之間的關系還需要更長時間的觀察研究。
致謝:數據收集中得到北京麋鹿生態實驗中心王建昌、包海元的幫助,氣象數據分析中得到國家氣候中心中國氣象局氣候研究開放實驗室趙俊虎的幫助,特此致謝。
