扶桑綿粉蚧交配行為及能力的研究
2020-11-11 08:51:06潘志萍陸永躍
環(huán)境昆蟲(chóng)學(xué)報(bào) 2020年5期
孫 峰,潘志萍,陸永躍*
(1. 華南農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院,廣州 510642;2. 廣東省科學(xué)院動(dòng)物研究所,廣東省動(dòng)物保護(hù)與資源利用重點(diǎn)實(shí)驗(yàn)室, 廣東省野生動(dòng)物保護(hù)與利用公共實(shí)驗(yàn)室,廣州 520620)
昆蟲(chóng)多以?xún)尚陨撤绞竭M(jìn)行后代繁殖,其特點(diǎn)是雌雄個(gè)體必須通過(guò)兩性交配使雄成蟲(chóng)的精細(xì)胞與雌成蟲(chóng)體內(nèi)的卵細(xì)胞結(jié)合形成受精卵以開(kāi)始新個(gè)體的發(fā)育。因此,交配是兩性生殖的昆蟲(chóng)選擇配偶及繁衍后代最重要、最基本的環(huán)節(jié) (Miyatake,1997)。交配行為的本質(zhì)就是刺激雌蟲(chóng)產(chǎn)卵 (Opp,1986),雄性個(gè)體通過(guò)交配增加其基因的傳播機(jī)會(huì),提高雄性的遺傳適合度 (張延華,2006) ,雌性個(gè)體在交配過(guò)程中可以得到雄蟲(chóng)的精液或者精囊中的營(yíng)養(yǎng)物質(zhì),提高雌蟲(chóng)的生殖能力(Tregenza,1998) 。因此,在復(fù)雜多變的外界環(huán)境下,雌蟲(chóng)、雄蟲(chóng)通過(guò)多次交配,對(duì)后代種群增長(zhǎng)和進(jìn)化具有重要作用(王超等,2014)。
扶桑綿粉蚧PhenacoccussolenopsisTinsley 是 2008年發(fā)現(xiàn)入侵中國(guó)大陸的重要害蟲(chóng) (陸永躍等,2008; 武三安和張潤(rùn)志,2009),并隨著入侵發(fā)生范圍擴(kuò)大,其寄主范圍呈逐步增大的趨勢(shì)(Arifetal.,2009; 張潤(rùn)志等,2010)。研究表明,由于該蟲(chóng)在我國(guó)潛在分布地區(qū)廣泛,我國(guó)大部分地區(qū)都適合扶桑綿粉蚧生存,若全面暴發(fā)將造成近100 億的重大損失,對(duì)農(nóng)業(yè)、林業(yè)生產(chǎn)構(gòu)成較大的威脅(王艷平等,2009)。
目前,扶桑綿粉蚧的生殖方式仍存在爭(zhēng)議。有報(bào)道認(rèn)為扶桑綿粉蚧可營(yíng)兩性生殖和孤雌生殖 (Vennilaetal.,2010; Sahitoetal.,2010) ,但也有一些研究認(rèn)為該蟲(chóng)只營(yíng)兩性生殖(Arifetal.,2012; Prasadetal.,2012 ) 。王超等(2014)對(duì)我國(guó)廣東、廣西、海南、福建、上海、浙江、安徽、湖北等地11個(gè)入侵種群生殖方式調(diào)查中僅發(fā)現(xiàn)了該蟲(chóng)的兩性生殖行為。筆者在試驗(yàn)過(guò)程中發(fā)現(xiàn)類(lèi)似情況,扶桑綿粉蚧種群在無(wú)雄蟲(chóng)情況下雌蟲(chóng)不能產(chǎn)卵。因此,在扶桑綿粉蚧兩性生殖種群中,雌雄成蟲(chóng)的交配行為、交配能力對(duì)種群的繁殖顯得非常重要。本文僅從扶桑綿粉蚧的交配行為及交配能力進(jìn)行觀察,以期明確其交配行為和交配能力,以期為進(jìn)一步了解扶桑綿粉蚧的交配行為和性信息素研究提供科學(xué)依據(jù)。
1 材料與方法
1.1 供試蟲(chóng)源及飼養(yǎng)方法
供試扶桑綿粉蚧采自廣州地區(qū)的扶桑,采回的扶桑綿粉蚧在室溫下接入扶桑植株飼養(yǎng)3代后備用。試驗(yàn)中扶桑綿粉蚧采用培養(yǎng)皿飼養(yǎng)進(jìn)行觀察,用細(xì)毛筆小心挑選2齡若蟲(chóng),將若蟲(chóng)置于備用的培養(yǎng)皿(直徑9 mm)里的扶桑葉片上。每培養(yǎng)皿中的葉片采自實(shí)驗(yàn)室種植的健康扶桑植株,葉片下加上一層培養(yǎng)基,以保持葉片新鮮。培養(yǎng)皿口用保鮮膜緊密覆蓋,保鮮膜上用昆蟲(chóng)針扎80~100個(gè)小孔,保持氣流通暢,保證試蟲(chóng)不能爬離培養(yǎng)皿。定期更換培養(yǎng)基和葉片。待觀察到雄蟲(chóng)進(jìn)入預(yù)蛹狀態(tài),則把雄蟲(chóng)與雌蟲(chóng)隔離飼養(yǎng),從而獲得未交配的扶桑綿粉蚧雄蟲(chóng)和雌蟲(chóng)。
1.2 扶桑綿粉蚧交配節(jié)律和行為的觀察
觀察培養(yǎng)皿中扶桑綿粉蚧雄蟲(chóng)的羽化時(shí)間,把羽化后的扶桑綿粉蚧雄成蟲(chóng)與雌成蟲(chóng)放置于同一個(gè)培養(yǎng)皿中,觀察其交配行為。同時(shí),把剛進(jìn)入成蟲(chóng)期的扶桑綿粉蚧雌蟲(chóng)與雄成蟲(chóng)放置于同一個(gè)培養(yǎng)皿中,觀察和記錄其交配時(shí)間和交配行為。
1.3 扶桑綿粉蚧雄蟲(chóng)交配能力的觀察
在每個(gè)培養(yǎng)皿中放置10頭扶桑綿粉蚧2日齡以上未交配的雌成蟲(chóng),然后用毛筆小心挑選扶桑綿粉蚧未交配的雄成蟲(chóng)置于各培養(yǎng)皿中,每皿引入1頭雄蟲(chóng),設(shè)30組重復(fù)。培養(yǎng)皿口用保鮮膜緊密覆蓋,保鮮膜上用昆蟲(chóng)針扎80~100個(gè)小孔,觀察扶桑綿粉蚧雄成蟲(chóng)的交配能力,至雄蟲(chóng)死亡,記錄交配時(shí)間和交配次數(shù)。
1.4 扶桑綿粉蚧雌蟲(chóng)交配能力的觀察
在每個(gè)培養(yǎng)皿中放置1頭扶桑綿粉蚧2日齡以上未交配的雌成蟲(chóng),用毛筆小心挑選扶桑綿粉蚧未交配的雄成蟲(chóng)置于各培養(yǎng)皿中,每皿引入5頭雄蟲(chóng),設(shè)30組重復(fù)。培養(yǎng)皿口用保鮮膜緊密覆蓋,保鮮膜上用昆蟲(chóng)針扎80~100個(gè)小孔,觀察扶桑綿粉蚧雌成蟲(chóng)的交配能力,至雄蟲(chóng)死亡,記錄交配時(shí)間和交配次數(shù)。
2 結(jié)果與分析
2.1 扶桑綿粉蚧的交配節(jié)律
扶桑綿粉蚧雄蟲(chóng)的羽化時(shí)間主要集中在凌晨3 ∶00-6 ∶00,雄蟲(chóng)羽化出來(lái)之后可以立刻尋找雌蟲(chóng)進(jìn)行交配。而雌蟲(chóng)經(jīng)第三次蛻皮進(jìn)入成蟲(chóng)后不會(huì)立刻進(jìn)行交配,當(dāng)雌蟲(chóng)蛻皮后約24 h后,雄蟲(chóng)才漸漸地靠近雌蟲(chóng),進(jìn)行交配。
將扶桑綿粉蚧未交配的雌蟲(chóng)和雄蟲(chóng)置于同一培養(yǎng)皿中,雄蟲(chóng)很快爬向雌蟲(chóng),可在任意時(shí)間進(jìn)行交配。交配前,雌蟲(chóng)始終處于被動(dòng)地位,大多數(shù)時(shí)間都固定在寄主植物上,而雄蟲(chóng)在交配中處于主動(dòng)性,當(dāng)雄蟲(chóng)感應(yīng)到雌蟲(chóng)后,會(huì)爬向雌蟲(chóng)。當(dāng)雄蟲(chóng)接觸到雌蟲(chóng)后,多數(shù)雌蟲(chóng)會(huì)很快感應(yīng)到雄蟲(chóng)接近,然后抬起尾部,以接受雄蟲(chóng)交配。雄蟲(chóng)找到雌蟲(chóng)后不能立刻交配成功,雄蟲(chóng)在雌蟲(chóng)背部來(lái)回爬動(dòng),腹部則不斷向下彎曲以尋找雌蟲(chóng)生殖器。當(dāng)雄蟲(chóng)生殖器與雌蟲(chóng)生殖器交合成功,即交配開(kāi)始,此時(shí)雄蟲(chóng)的足緊緊抓住雌蟲(chóng)尾部,幾乎呈靜止?fàn)顟B(tài)。當(dāng)交配結(jié)束,多數(shù)雄蟲(chóng)會(huì)撲打翅膀,然后爬向另一頭雌蟲(chóng),進(jìn)行下一次交配。
2.2 扶桑綿粉蚧雄蟲(chóng)的交配能力
當(dāng)把1頭雄蟲(chóng)放入有10頭雌蟲(chóng)的培養(yǎng)皿中,雄蟲(chóng)會(huì)很快尋找雌蟲(chóng),并與之交配。通常情況下,雄蟲(chóng)與1頭雌蟲(chóng)交配結(jié)束之后,會(huì)很快尋找下一頭雌蟲(chóng)進(jìn)行交配,1頭雄蟲(chóng)可以連續(xù)與多頭雌蟲(chóng)進(jìn)行交配,但也有少數(shù)情況,雄蟲(chóng)與同一頭雌蟲(chóng)進(jìn)行1次以上交配。在所觀察扶桑綿粉蚧30頭雄蟲(chóng)分別與10頭雌蟲(chóng)的交配中,雄蟲(chóng)交配次數(shù)為3~10次不等(圖1),其中以交配6次所占的比率最大(26.7%),其次為交配8次(20%),平均每頭雄蟲(chóng)交配6.9次。

圖1 扶桑綿粉蚧單頭雄蟲(chóng)的交配次數(shù)分布Fig.1 Distribution of mating frequency of single male of Phenacoccus solenopsis
所觀察扶桑綿粉蚧1雄多雌的交配中,每一頭雄蟲(chóng)交配時(shí)間差異較大,需18.1 ~228 min,平均每頭雄蟲(chóng)交配總時(shí)間為79.9 min,如果將單頭雄蟲(chóng)交配時(shí)間跨度按30 min一個(gè)等級(jí)進(jìn)行劃分時(shí)發(fā)現(xiàn)(圖2),單頭雄蟲(chóng)交配時(shí)間比較分散,不同蟲(chóng)體之間差異較大。
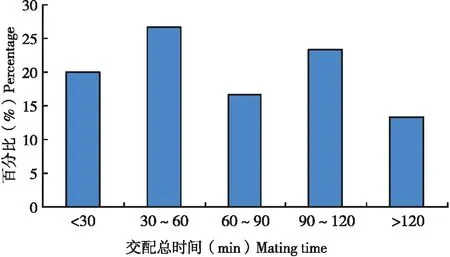
圖2 扶桑綿粉蚧單頭雄蟲(chóng)交配時(shí)間分布Fig.2 Distribution of mating time of single male of Phenacoccus solenopsis
試驗(yàn)中30頭雄蟲(chóng)總共交配208次,每一次交配持續(xù)時(shí)間長(zhǎng)度差異較大,從1.8~76.5 min不等,平均每次交配時(shí)間為11.5 min,如果將交配持續(xù)時(shí)間跨度按10 min一個(gè)等級(jí)進(jìn)行劃分時(shí)發(fā)現(xiàn)(圖3),持續(xù)時(shí)間在10 min以?xún)?nèi)的發(fā)生概率最大(68.8%),持續(xù)時(shí)間為10~20 min的發(fā)生概率次之(18.3%),這表明在扶桑綿粉蚧1雄多雌的交配中,大多數(shù)交配時(shí)間都在10 min以?xún)?nèi)完成。
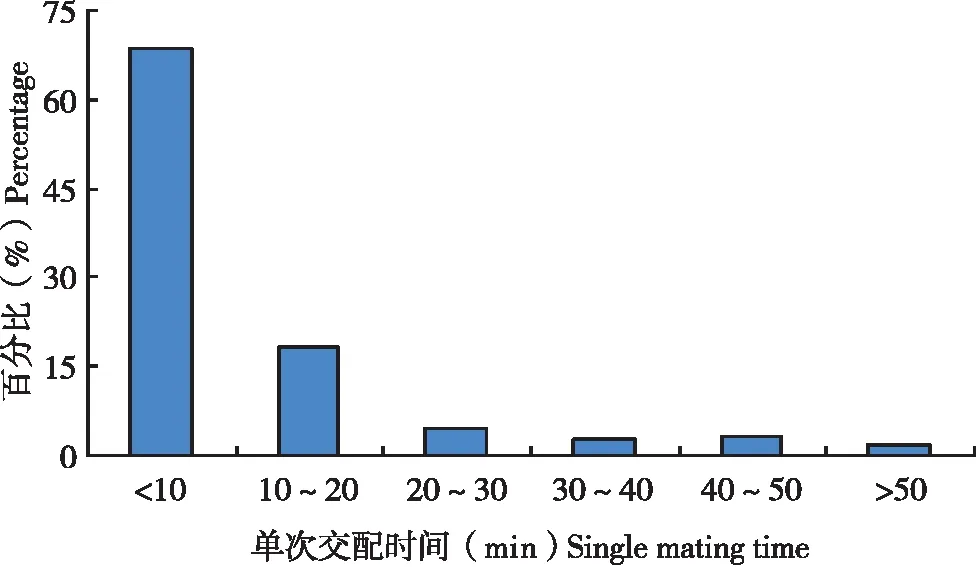
圖3 扶桑綿粉蚧雄蟲(chóng)單次交配時(shí)間分布Fig.3 Distribution of single mating time of Phenacoccus solenopsis male
2.3 扶桑綿粉蚧雌蟲(chóng)的交配能力
試驗(yàn)中,當(dāng)把5頭雄蟲(chóng)放入只有1頭雌蟲(chóng)的培養(yǎng)皿中,不多久就會(huì)有雄蟲(chóng)爬來(lái)與之交配,當(dāng)其中1頭雄蟲(chóng)與雌蟲(chóng)交配時(shí),其它雄蟲(chóng)因?yàn)閷ふ也坏酱葡x(chóng),也會(huì)爬向這1頭雌蟲(chóng),并且在雌蟲(chóng)背部來(lái)回爬動(dòng),雄蟲(chóng)交配器不斷向下彎曲,試圖與之交配,在多次試圖交配未果后,這些雄蟲(chóng)則爬離雌蟲(chóng)。在所觀察扶桑綿粉蚧30頭雌蟲(chóng)分別與5頭雌蟲(chóng)的交配中,單頭雌蟲(chóng)的交配次數(shù)為1~4次不等(圖4),以交配2次所占的比率最大(40.0%),其次為交配3次(33.3%),平均每頭雌蟲(chóng)交配2.3次。
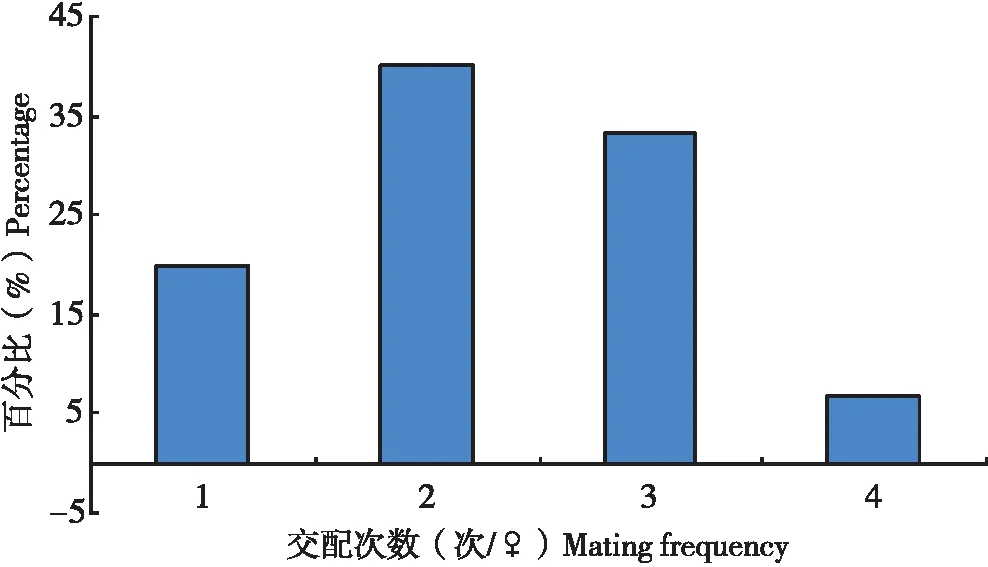
圖4 扶桑綿粉蚧單頭雌蟲(chóng)交配次數(shù)分布Fig.4 Distribution of mating times of single female of Phenacoccus solenopsis
在所觀察扶桑綿粉蚧1雌多雄的交配中,每一頭雌蟲(chóng)交配時(shí)間差異較大,從10.6 min到83.7 min不等,平均每頭雌蟲(chóng)交配總時(shí)間為36.3 min,如果將單頭雌蟲(chóng)交配總時(shí)間跨度按10 min一個(gè)等級(jí)進(jìn)行劃分時(shí)發(fā)現(xiàn)(圖5),單頭雌蟲(chóng)交配時(shí)間比較分散,不同蟲(chóng)體之間差異較大。

圖5 扶桑綿粉蚧單頭雌蟲(chóng)交配時(shí)間Fig.5 Mating time of single female of Phenacoccus solenopsis
30頭雌蟲(chóng)總共交配68次,每一次交配持續(xù)時(shí)間差異明顯,從4.4 min到51.5 min不等,平均每次交配時(shí)間為16 min,如果將交配持續(xù)時(shí)間跨度按10 min一個(gè)等級(jí)進(jìn)行劃分時(shí)發(fā)現(xiàn)(圖6),持續(xù)時(shí)間在10 min以?xún)?nèi)的發(fā)生概率最大(41.2%),持續(xù)時(shí)間為10~20 min的發(fā)生概率次之(30.9%)。這表明在單次交配中,1雌多雄的平均交配持續(xù)時(shí)間長(zhǎng)于1雄多雌的平均交配時(shí)間。

圖6 扶桑綿粉蚧雌蟲(chóng)單頭交配時(shí)間分布Fig.6 Distribution of single mating time of Phenacoccus solenopsis female
3 結(jié)論與討論
本研究通過(guò)觀察發(fā)現(xiàn)扶桑綿粉蚧雌雄蟲(chóng)均可以多次交配,其中雄蟲(chóng)的交配能力強(qiáng)于雌蟲(chóng),雄蟲(chóng)羽化后即尋找雌蟲(chóng)進(jìn)行交配,此時(shí),雄蟲(chóng)不再取食,當(dāng)雄蟲(chóng)完成交配任務(wù)后,會(huì)很快死去,而雌蟲(chóng)交配后對(duì)寄主的取食和危害更大,當(dāng)雌蟲(chóng)個(gè)體生物量達(dá)到最大時(shí),開(kāi)始繁殖產(chǎn)出下一代。這樣有利于所有雌蟲(chóng)得到順利交配,同時(shí)雌蟲(chóng)有巨大的繁殖力,從而有利于扶桑綿粉蚧種群數(shù)量的迅速擴(kuò)大,容易爆發(fā)成災(zāi)。扶桑綿粉蚧在交配中的每次交配時(shí)間差異較大,從1.8 min到76.5 min不等,但是多數(shù)時(shí)候都在10 min以?xún)?nèi)(68.8%),但雌蟲(chóng)交配時(shí)間與最終繁殖量是否有關(guān)聯(lián),需要作進(jìn)一步研究。
研究發(fā)現(xiàn),扶桑綿粉蚧雄蟲(chóng)對(duì)雌蟲(chóng)則有較強(qiáng)的搜索能力,即便把雌蟲(chóng)放置在隱蔽的地方,雄蟲(chóng)也可以通過(guò)嗅覺(jué)來(lái)尋找到雌蟲(chóng)并進(jìn)行交配。完全交配后的雌蟲(chóng)對(duì)雄蟲(chóng)沒(méi)有吸引作用,即使把雄蟲(chóng)放置在完全交配后的雌蟲(chóng)旁邊,雄蟲(chóng)并不會(huì)靠近雌蟲(chóng),推測(cè)雌成蟲(chóng)自身分泌性信息素有關(guān)。性信息素在性成熟的處女蚧中一直存在, 但其釋放不是連續(xù)不斷, 而是可以控制的。雌成蟲(chóng)只有在求偶時(shí)才釋放性信息素引誘雄成蟲(chóng)前來(lái)交配(周林等,1999)。交配可直接影響性信息素的釋放。松突圓蚧未交配雌成蚧和去殼處女蚧均對(duì)雄蚧具引誘作用,而交配1 h后的雌蚧逐漸失去引誘雄蚧的能力(杜家緯等,1991)。日本松干蚧雌蚧一旦交配,便馬上停止性信息素的分泌和釋放(祁云臺(tái)等,1983)。同樣,本研究也觀察到類(lèi)似的情況,扶桑綿粉蚧雌成蟲(chóng)一旦交配后,雄成蟲(chóng)就不再靠近雌成蟲(chóng),推測(cè)其停止性信息素的分泌和釋放,從而不再吸引雄成蟲(chóng)。
利用性信息素,應(yīng)用誘殺技術(shù)和交配干擾技術(shù),可以達(dá)到防治害蟲(chóng)目的。應(yīng)用昆蟲(chóng)性信息素,能有效地減少化學(xué)農(nóng)藥所造成的環(huán)境污染、害蟲(chóng)產(chǎn)生抗性以及殺傷天敵等問(wèn)題,保護(hù)生態(tài)系統(tǒng)平衡,在害蟲(chóng)的治理中具有十分重要的作用。目前,尚未見(jiàn)利用該誘殺技術(shù)大面積防治蚧蟲(chóng)的報(bào)道,但已有這方面研究報(bào)道。據(jù)以色列報(bào)道,交配干擾能成功地用作為紅圓蚧的防治措施(韓桂彪,1999)。祁云臺(tái)等(1989)采用屋脊式紙板誘捕器在江蘇無(wú)錫大浮松林中對(duì)日本松干蚧進(jìn)行小規(guī)模的誘殺試驗(yàn), 獲得了明顯的誘蟲(chóng)效果。本研究觀察到采自廣州的扶桑綿粉蚧種群的生殖方式僅為有性生殖,即無(wú)雄蟲(chóng)存在的情況下雌蟲(chóng)不能產(chǎn)卵,因此可以利用性信息素阻礙雌雄蟲(chóng)交配來(lái)進(jìn)行防治。有性生殖的生殖方式能使產(chǎn)卵期提前,但并沒(méi)有提高產(chǎn)卵量。雄性個(gè)體存在的意義可能只是為了適應(yīng)不良環(huán)境,有利于種群繁殖(魏婷等,2010)。但是扶桑綿粉蚧雌雄蟲(chóng)均能夠進(jìn)行多次交配,交配能力強(qiáng),對(duì)于利用性信息素大量誘捕進(jìn)行防治是一個(gè)不利的因素,仍需要進(jìn)一步加強(qiáng)多種蚧蟲(chóng)性信息素生物合成、生態(tài)遺傳學(xué)及進(jìn)化、在近緣種間生殖隔離的作用以及人工合成性信息素在種群監(jiān)測(cè)及防治中的應(yīng)用技術(shù)等研究。因此,如何利用性信息素對(duì)該蟲(chóng)進(jìn)行防治,仍有待進(jìn)一步深入研究。
