高產阿魏酸酯酶菌株的篩選鑒定及其酶活測定條件優化
2020-11-02 08:50:38王祥鋒汪喬袁慧君王麗
生物技術通報 2020年10期
王祥鋒 汪喬 袁慧君 王麗
(山東農業大學植物保護學院 山東省農業微生物重點實驗室,泰安 271018)
阿 魏 酸 酯 酶(EC 3.1.1.73,Ferulic Esterase,FAE),屬于羧酸酯水解酶亞類,該酶可以水解植物細胞壁中半纖維素之間、以及半纖維素與木質素之間由阿魏酸連接形成的酯鍵,提高木質纖維素降解效率的同時還釋放出阿魏酸、p-香豆酸等抗氧化物質,FAE也具有參與合成酯類物質的能力[1],在食品、造紙、飼料、醫藥、化妝品等行業具有很高的應用價值和廣闊的市場前景[2-6]。
目前已報道的阿魏酸酯酶主要來源于微生物,其中研究最多的是黑曲霉Aspergillus niger,已經從中分離到多種阿魏酸酯酶,但酶活力普遍較低[7-9]。已報道的其他產阿魏酸酯酶的微生物菌株主要是從人和動物的腸道、反芻動物瘤胃中分離得到,大部分為厭氧微生物,分離培養較為困難,而且產生的酶活力較低,不能滿足工業應用的要求[10-12]。鑒于阿魏酸酯酶具有的廣泛用途,獲得廉價和高活力的阿魏酸酯酶是推動其應用的前提,因此篩選容易培養且高產阿魏酸酯酶的菌株顯得尤為重要。
大型真菌是菌物中能形成大型子實體的一類真菌,很多種類都具有較高的營養價值和藥用價值,資源豐富、取材方便。大型真菌在生長過程中,能分泌一系列降解纖維素、半纖維素和木質素的酶,如漆酶、木素過氧化物酶、錳過氧化物酶、纖維素酶、木聚糖酶等[13-15],由此可以推測,其中可能蘊藏著豐富的阿魏酸酯酶資源。基于此,本研究通過對實驗室保藏的106株大型真菌菌株進行篩選并獲得2個高產阿魏酸酯酶的菌株,進一步對其進行了分子生物學鑒定,并分別測定了2個菌株以米糠為培養基產阿魏酸酯酶的酶活曲線,優化了其粗酶液的酶活測定條件,為下一步阿魏酸酯酶的分離純化和應用提供理論基礎。
1 材料與方法
1.1 材料
1.1.1 樣品 106株大型真菌菌株為山東農業大學菌物實訓實踐基地實驗室保藏的菌株,其中包括采集于泰山周邊的野生菌株和部分栽培食用菌菌株。
1.1.2 化學試劑 阿魏酸甲酯、阿魏酸乙酯購自Sigma公司。Taq DNA 聚合酶、dNTP等分子試劑購自北京全式金生物技術有限公司。其他常規試劑為國產分析純。
1.1.3 培養基 PDA培養基:馬鈴薯(去皮)200 g,葡萄糖20 g,KH2PO42 g,MgSO4·7H2O 0.5 g,維生素B110 mg,瓊脂19 g,水1 000 mL,pH自然。
初篩培養基:用分析天平稱取酵母粉10 g,KCl 0.5 g,MgSO4·7H2O 0.5 g,K2HPO41.0 g,瓊脂20 g,配制成1 L培養基,pH值自然;在高壓滅菌鍋中121℃滅菌20 min;冷卻至60℃左右添加0.1%的阿魏酸乙酯(溶于N,N-二甲基甲酰胺溶液,過濾除菌),搖至均勻的乳白色倒平板[16]。
固體發酵培養基:麥麩或米糠10 g,加入無機鹽溶液(g/L,KH2PO42 g,MgSO4·7H2O 0.5 g)13 mL,攪拌均勻后于121℃下滅菌30 min,冷卻至室溫。
1.2 方法
1.2.1 平板法初篩 將106株大型真菌的菌株進行活化,之后用0.5 cm直徑的打孔器打孔,接種到以阿魏酸乙酯為唯一碳源的初篩培養基平板上,25℃恒溫培養3 d,觀察是否出現透明水解圈,并測定透明圈直徑,重復3次。若菌株可以產生阿魏酸酯酶,則會分解培養基中的底物阿魏酸乙酯而形成透明圈,透明圈的直徑大小反應阿魏酸酯酶的活性高低,根據透明圈大小初步篩選出阿魏酸酯酶活力高的菌株。
1.2.2 固體發酵培養復篩 將初篩得到的16個菌株,在PDA平板上活化后用直徑1 cm的打孔器打孔,分別接種3塊圓餅到裝有無菌固體麥麩和米糠培養基的三角瓶中,25℃恒溫培養6 d。每個處理3個重復。培養結束后向每個三角瓶中加入50 mL磷酸緩沖液,攪拌均勻后置于40℃恒溫水浴鍋內,浸提25 min;過濾收集浸提液后于4℃、10 000 r/min離心15 min,上清液即為粗酶液,用于酶活的測定。
1.2.3 阿魏酸酯酶酶活的測定 取10 μL粗酶液加入到400 μL阿魏酸甲酯溶液(50 μmol/L,pH 5.0)中,混合均勻,置于40℃水浴鍋中反應30 min,反應完成后在分光光度下測定340 nm處的吸光度[17],對照組在測定前加入10 μL酶液,其余條件相同。該方法測定原理為阿魏酸甲酯在340 nm下具有最大光吸收,阿魏酸酯酶可以降解阿魏酸甲酯中的酯鍵使吸光度下降,通過測定340 nm下的吸光度差值(ΔA340)來計算不同樣品的相對酶活。
1.2.4 菌株的ITS分子鑒定 將經過初篩和復篩后獲得的兩株高產阿魏酸酯酶的菌株在PDA平板中培養,收集菌絲體,采用液氮研磨法提取基因組DNA,采用ITS通用引物,ITS1:5'-T C C G T A G G T G A A C C T G C G G-3',I T S 4:5'-TCCTCCGCTTATTGATATGC-3',進行PCR擴增。PCR擴增程序:94℃ 5 min;95℃ 0.5 min,58℃ 0.5 min,72℃ 1 min,35個循環;72℃ 10 min。將PCR產物送華大基因科技有限公司純化后測序。
1.2.5 序列同源性分析及系統發育樹的構建 使用Blast功能將測得的ITS序列在GenBank中進行同源性比對,下載同源性高的序列并使用Mega 5.2 軟件進行系統發育樹的構建。
1.2.6 發酵米糠產阿魏酸酯酶的酶活曲線測定 將篩選獲得的兩株高產阿魏酸酯酶的大型真菌菌株接種到PDA平板上培養,菌絲長滿后用直徑1 cm的打孔器打孔,分別接種3塊圓餅狀菌塊到滅菌的米糠培養基中。25℃培養,每隔1 d取樣進行酶活測定,共培養14 d。
1.2.7 粗酶液酶活測定條件優化 采用單因素法,分別考察反應體系中溫度、酶液添加量和反應時間等對兩個菌株粗酶液阿魏酸酯酶酶活測定的影響。
分別收集兩個菌株粗酶液活性最高的發酵液,以20 μL粗酶液、400 μL阿魏酸甲酯溶液、40℃和30 min反應時間作為基礎反應條件。單一改變基礎反應條件中的變量,分別測定粗酶液添加量為10、20、30、40、50 μL時,反應時間為10、15、20、25、30、35、40 min時,反應溫度為20、30、40、50、60℃時,兩個菌株粗酶液中阿魏酸酯酶的活性。以每個因素中酶活力最高者為100%,其他各梯度下與該酶活的比值為該單因素下的相對酶活。計算確定兩個菌株粗酶液阿魏酸酯酶活性的最佳測定條件。
2 結果
2.1 產阿魏酸酯酶大型真菌菌株的初篩
將106個大型真菌菌株通過透明圈法進行產阿魏酸酯酶的初篩,發現92個菌株都可以產生透明圈,其中有16株菌株產生的透明圈直徑較大(≥3 cm),結果如圖1所示。從圖中可以看出,菌株M產生的透明圈最大,C、D、E、F、N和P等6個菌株產生的透明圈大小相近。通過透明圈大小反應了各個菌株產阿魏酸酯酶的活性高低。
2.2 高產阿魏酸酯酶大型真菌菌株的復篩
將初篩獲得的16株高產阿魏酸酯酶的菌株分別接種到麥麩培養基和米糠培養基中進行固體發酵復篩,測定各菌株的阿魏酸酯酶活性,其結果見圖2、3。由圖中可以看出,不同菌株在兩種培養基上表現出不同的產酶情況,A、C、D、F、H、I、K、M、N、O等10個菌株在麥麩和米糠培養基中都能產生較高活力的阿魏酸酯酶;而菌株L只在麥麩上可以產生高活力阿魏酸酯酶;菌株J只在米糠上可以產生高活力阿魏酸酯酶;菌株B、E、G、P在兩種培養基上產酶活性都比較低。最后選擇在兩種培養基上活性都較高且長勢較快的菌株D和M進行分子鑒定。
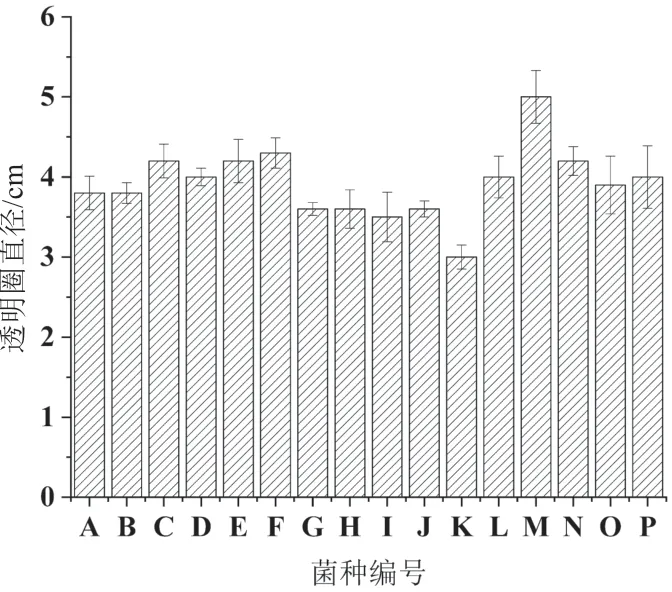
圖1 16株菌株的透明圈直徑大小
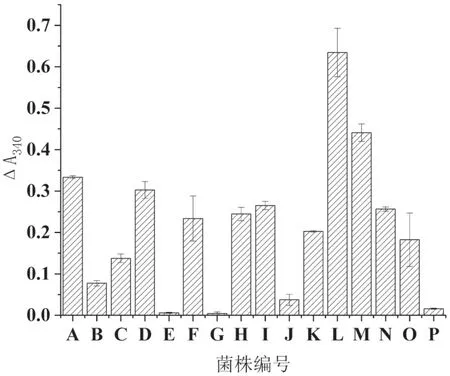
圖2 發酵麥麩培養基中16個菌株的阿魏酸酯酶活性
2.3 菌株分子鑒定
對菌株D和M分別進行ITS序列的擴增并進行電泳檢測。PCR產物擴增條帶單一,經測序,菌株D和M的ITS序列長度分別為630 bp和662 bp,使用Blast功能將測得的ITS序列在GenBank中進行同源性比對,并與其他同源性高的菌株進行系統發育樹的構建,結果如圖4所示。結果顯示菌株D與一色齒毛菌Cerrena unicolor的相似性最高,達99%;菌株M與血紅密孔菌Pycnoporus sanguineus的相似性最高,達99%。結合子實體形態(圖5),最終鑒定菌株D為一色齒毛菌,菌株M為血紅密孔菌。
2.4 兩個菌株發酵米糠產阿魏酸酯酶的酶活曲線
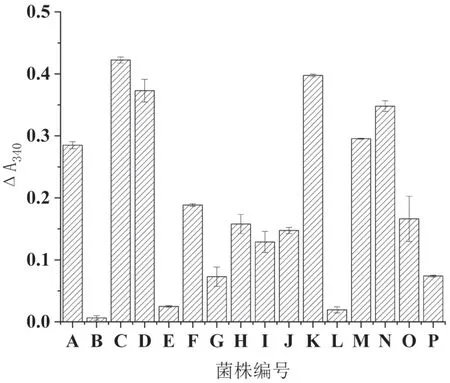
圖3 發酵米糠培養基中16個菌株的阿魏酸酯酶活性
分別對兩個菌株固體發酵米糠產阿魏酸酯酶情況進行了測定,酶活力隨發酵時間變化曲線見圖6,由圖中可以看出,菌株D從第2天開始產酶,第4天達到產酶高峰,隨著培養時間增加,酶活力開始緩慢下降;菌株M從第4天開始產酶,第6天酶活最高,之后隨著培養時間增加,酶活力開始下降。發酵14 d,兩個菌株粗酶液中依然具有較高的阿魏酸酯酶活性。
2.5 粗酶液阿魏酸酯酶酶活測定條件優化
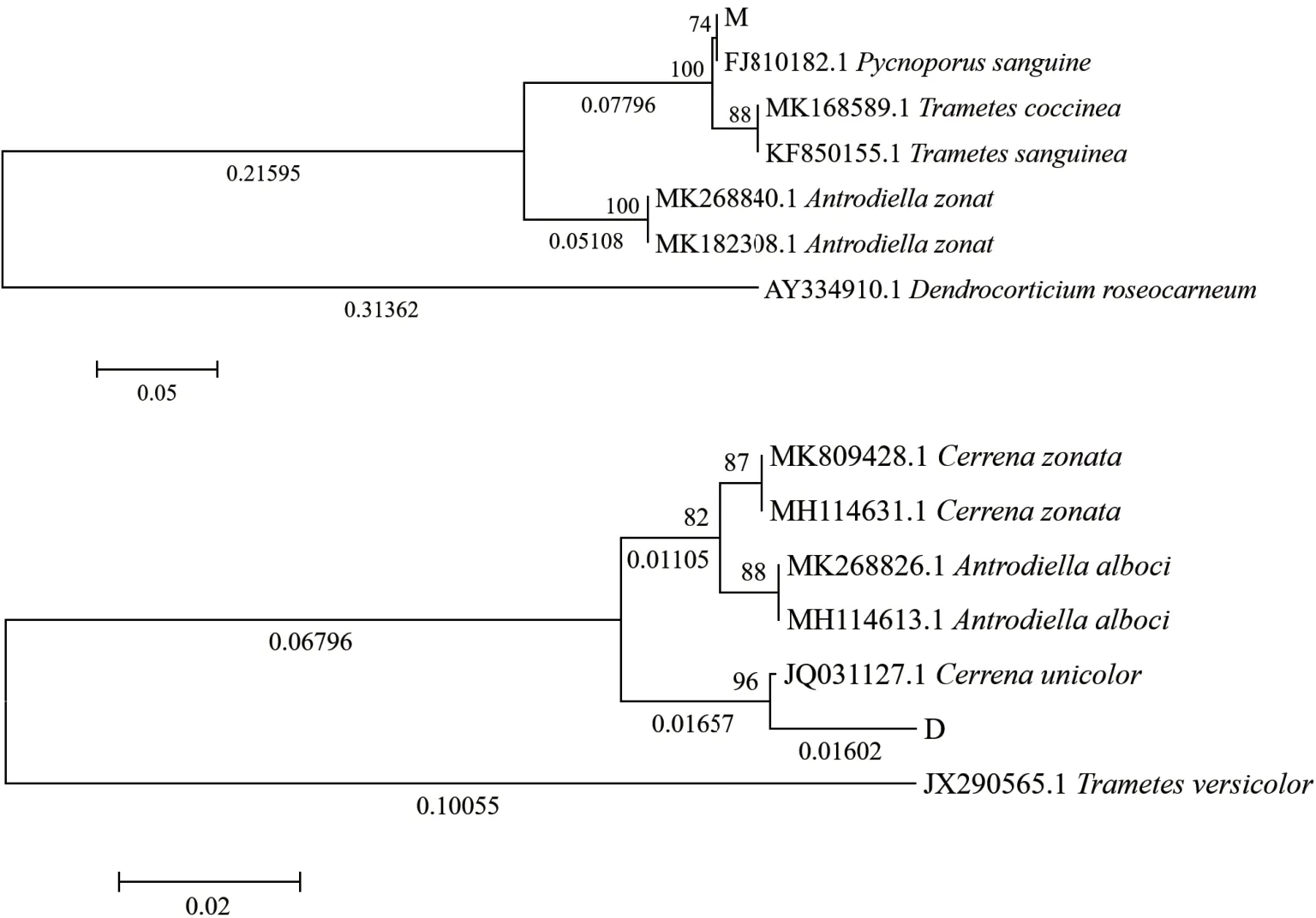
圖4 菌株D、M的ITS序列系統發育樹分析

圖5 兩個菌株的野外子實體圖片
分別收集兩種菌發酵米糠產酶活性最高的粗酶液(一色齒毛菌發酵4 d,血紅密孔菌發酵6 d)進行阿魏酸酯酶活性測定條件的優化,主要包括最佳粗酶液添加量、最適反應溫度、最佳反應時間等指標,具體結果如下所述。
2.5.1 粗酶液酶活的最適反應溫度測定 分別在20-60℃測定兩個菌株粗酶液中阿魏酸酯酶的活性,結果見圖7。由圖中可以得出,一色齒毛菌的阿魏酸酯酶最適反應溫度為40℃,在30℃和50℃分別保留73%和94%的活性;血紅密孔菌的阿魏酸酯酶最適反應溫度為50℃,隨著溫度升高,活性迅速下降,60℃時僅剩余40%的活性。
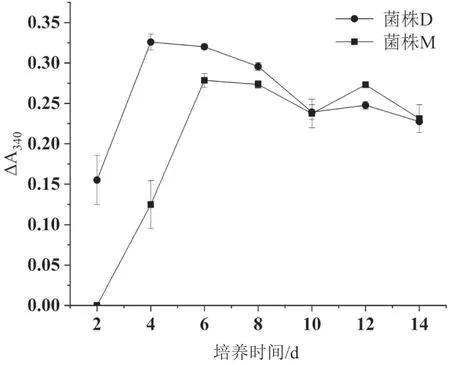
圖6 兩個菌株的酶活力隨發酵時間變化曲線
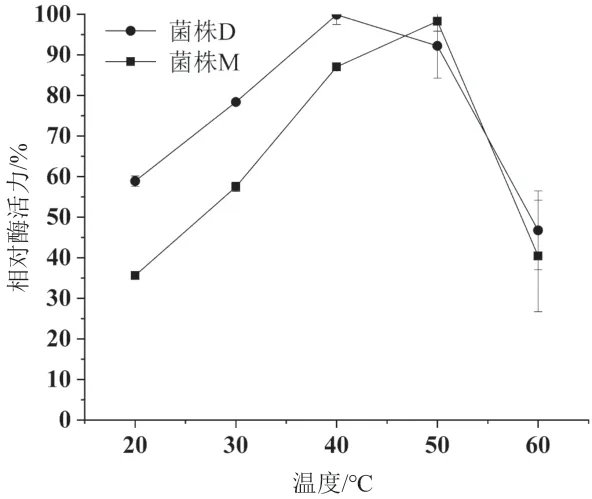
圖7 兩個菌株粗酶液中酶活最適反應溫度測定
2.5.2 粗酶液酶活的最佳反應時間測定 分別測定了反應時間為10、15、20、25、30、35、40 min時,兩個菌株粗酶液中阿魏酸酯酶的活性,結果見圖8。由圖中可以看出,隨著反應時間的延長,兩個菌株粗酶液中的酶活性逐步增加,當反應時間達到35 min時,酶活性達到最大。之后隨著時間增加,酶活性開始下降。
2.5.3 粗酶液添加量對酶活力的影響 分別測定了不同粗酶液添加量(10、20、30、40、50 μL)對酶活測定結果的影響,結果見圖9。由圖中可以看出,兩個菌株的酶活力變化趨勢相似,當粗酶液添加量較低時,隨著粗酶液的增加,酶活力也增加,當粗酶液添加量為20 μL時兩個菌株的酶活力最高,并趨于穩定,當添加量大于30 μL時酶活力開始下降。
3 討論
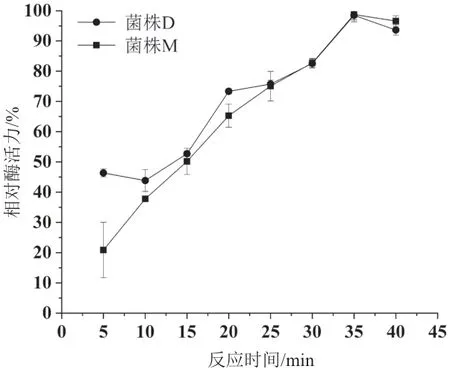
圖8 兩個菌株粗酶液中酶活最佳反應時間測定
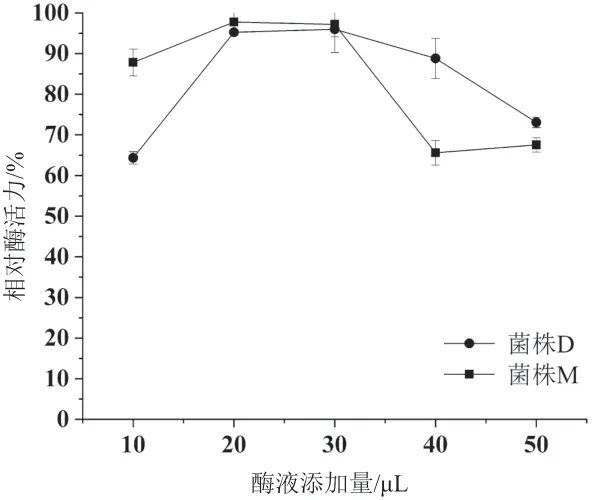
圖9 粗酶液添加量對酶活力的影響
阿魏酸酯酶可以降解由阿魏酸形成的酯鍵,因此通常采用是否可以降解阿魏酸乙酯產生透明圈來進行酶活初篩[16,18-19]。本研究通過平板透明圈法初篩得出86.8%的供試大型真菌菌株都具有降解阿魏酸乙酯的能力,根據透明圈大小進一步篩選出16個降解活性高的菌株,由此反映出大型真菌中阿魏酸酯酶的普遍性。這與Haase-Aschoff等[20]在擔子菌中的檢測結果一致,他們以阿魏酸甲酯、肉桂酸甲酯和3,4,5-三甲氧基肉桂酸甲酯3種阿魏酸酯衍生物為底物檢測了41株擔子菌發酵產阿魏酸酯酶的情況發現,61%的菌株對至少一種阿魏酸酯衍生物表現出降解活性。
阿魏酸酯酶是誘導酶,而且有多種不同類型,只有當培養基中含有阿魏酰酯鍵類結構時才會被誘導產生。研究發現,不同真菌的阿魏酸酯酶誘導培養基是不同的,而且同一菌株經不同培養基誘導可以產生不同類型的阿魏酸酯酶[5,21]。麥麩、米糠等谷物的殼中含有大量的阿魏酰酯鍵結構,可作為阿魏酸酯酶的誘導底物[22]。本實驗中,在對初篩獲得的16株高效降解阿魏酸乙酯的菌株進行固體發酵麥麩和米糠產阿魏酸酯酶復篩時也發現,這些菌株在3種底物上表現出了不同的產酶活性,推測原因為有些菌株在不同的誘導培養基上分泌了不同類型的阿魏酸酯酶,具體驗證還需要首先獲得各菌株在不同底物上分泌的阿魏酸酯酶的基因序列。
對篩選到的在阿魏酸乙酯、麥麩和米糠等3種培養基上都可以產生高活力阿魏酸酯酶的2個菌株進行分子鑒定,并結合野外子實體形態得出,2個菌株分別為一色齒毛菌Cerrena unicolor和血紅密孔菌Pycnoporus sanguineus,目前還未有關于這兩種大型真菌中阿魏酸酯酶的報道。這兩種菌在自然界中分布廣泛且生長迅速,推測這與其生長過程中分泌的豐富木質纖維素降解酶是密切相關的[23-24]。
阿魏酸酯酶是木聚糖支鏈降解酶,經該酶降解后才進一步暴露出木聚糖和纖維素主鏈,因此在木質纖維素的降解中發揮“先鋒軍”作用,通常在發酵早期就開始分泌[4-5,25]。本實驗中,2個菌株發酵米糠時均是在發酵前期就可以產生阿魏酸酯酶,且分別在第4天和第6天達到高峰,發酵14 d時依然保持較高活性,反映出阿魏酸酯酶在米糠降解中發揮關鍵的作用。分別選取2個菌株發酵米糠產酶活性最高的粗酶液對阿魏酸酯酶活性測定條件進行了優化,在粗酶液添加量的優化實驗中發現,兩個菌株的酶活趨勢相似:當粗酶液添加量較低時,隨著粗酶液的添加,酶活性先增加后趨于穩定,但當粗酶液高于一定值時,酶活性開始下降,推測粗酶液中含有其他影響酶活測定的成分,當增加到一定量時會對酶活測定結果產生影響,所以在酶活測定中應嚴格控制酶液的添加量。以上實驗為后續開展該酶的分離純化和進一步的應用提供了理論依據。
阿魏酸酯酶在木質纖維素高效降解和阿魏酸的生物法生產中發揮重要作用[5,26],本研究發現該酶在大型真菌中普遍存在,而且篩選到2株高產阿魏酸酯酶的菌株,這不僅豐富了大型真菌中的阿魏酸酯酶資源,而且還為揭示大型真菌對木質纖維素的高效降解機制提供了一定的思路。
4 結論
本研究從106個大型真菌菌株中篩選獲得2個高產阿魏酸酯酶的菌株,經分子鑒定分別為一色齒毛菌和血紅密孔菌。兩個菌株發酵麥麩和米糠都可以產生較高活力的阿魏酸酯酶,2個菌株發酵米糠時分別在第4天和第6天達到高峰,分別收集活性最高的粗酶液進行阿魏酸酯酶的酶活條件測定,結果表明:一色齒毛菌在40℃、反應時間為25 min、添加400 μL阿魏酸甲酯溶液和30 μL粗酶液條件下降解效率最高;血紅密孔菌在30℃、反應時間為35 min、添加400 μL阿魏酸甲酯溶液和20 μL粗酶液條件下降解效率最高。
