來源于P. bacterium 1109的甘露醇脫氫酶的重組純化及酶學性質研究
2020-10-23 11:34:18逯付之張文立光翠娥沐萬孟
食品工業科技 2020年19期
關鍵詞:甘露醇
逯付之,徐 煒,吳 昊,張文立,光翠娥,沐萬孟
(江南大學食品科學與技術國家重點實驗室,江蘇無錫 214122)
甘露醇又名D-甘露糖醇,是一種晶體狀不吸濕的功能性稀有己糖醇[1]。與蔗糖相比,甘露醇具有50%的甜度和較低的卡路里,目前在食品工業中主要用作功能性甜味劑[2],在制藥、化學和醫療行業中也得到了廣泛應用[3]。
甘露醇可以通過多種方法獲得,目前的合成方法主要包括有天然提取法、化學合成法、生物轉化法[4],其中生物轉化法又包括酶轉化法和微生物細胞轉化法。天然提取法主要是從海帶中提取,包括水重結晶法和電滲析器法[4]。在工業上,甘露醇主要是通過葡萄糖和果糖的混合物催化加氫制得,但是生產效率不高,果糖∶葡萄糖(質量比1∶1)混合可得到甘露醇∶山梨醇(質量比1∶3)的混合產物[5]。生物轉化法主要包括全細胞[6]和游離酶,酶的催化特異性強,催化底物的轉化率高,且出現副產物比例低,因而近年來利用生物轉化法合成甘露醇受到越來越多的關注[7]。利用生物轉化法把底物D-果糖轉化為甘露醇過程中,起關鍵作用的是甘露醇脫氫酶(MDH),同時需要輔酶因子NAD(P)H或NADH參與[8],在此基礎上,MDH還可以與甲酸脫氫酶聯用,構成NAD(P)H或NADH再生的的雙酶系統生產甘露醇[9]。目前已經有來自乳酸菌包括L.sanfranciscensis[10]、L.intermedius[11]、L.brevi[12]、L.reuteri[13]、L.mesenteroide[14]、L.pseudomesenteroide[15]的MDHs,還有來自其他細菌包括C.magnolia[16]、P.fluorescens[17]、R.sphaeroides[18]、T.maritima[19]和T.neapolitana[20]的MDHs被報道。
研究生物轉化法合成甘露醇過程中,關鍵是研究MDH的催化效率和催化條件。目前已經報道的MDHs中,大多數來源于中溫微生物,它們的熱穩定性相對較弱,不利于大規模工業生產中,特別是在長時間中生物轉化[21]。P.bacterium1109屬于耐熱菌屬,它的全基因組已經被測序并公布在Genebank上(登錄號為:LDJB01000003),經功能注釋存在一個編碼MDH的基因片段(Genebank登錄號為KLU39398.1)。本研究將來自P.bacterium1109的目的基因導入到E.coliBL21(DE3)中誘導表達,分離純化重組酶,同時研究了該重組酶的酶學性質以及反應條件,為酶法工業化生產甘露醇奠定理論基礎。
1 材料與方法
1.1 材料與儀器
E.coliBL21(DE3)、E.coliDH5α上海生工(生物工程)有限公司;載體pET-22b(+) 上海捷瑞生物工程有限公司;氨芐霉素(Amp)、異丙基硫代半乳糖苷(IPTG)、SDS-PAGE凝膠電泳試劑、標準蛋白Marker、D-果糖、D-核糖、D-木糖、L-山梨糖、D-阿洛糖、D-甘露糖、D-半乳糖、D-甘露糖醇、木糖醇、山梨醇、D-阿拉伯糖醇、半乳糖醇、阿洛糖醇、煙酰胺腺嘌呤二核苷酸(NAD+)、還原型輔酶煙酰胺腺嘌呤二核苷酸(NADH) Sigma-Aldrich公司;液體LB培養基、固體LB培養、脫色液 依據文獻[20]進行配制;其它試劑 均為國產分析純。
PE20型pH計 梅特勒-托利多儀器上海有限公司;WFH-201B7型蛋白電泳設備、MP3型凝膠成像系統 美國Bio-Rad公司;Waters2695型高效液相色譜儀(HPLC)、Sugar-PakTM鈣離子色譜柱 美國Waters公司;恒溫水浴鍋 上海生工有限公司;紫外/可見分光光度計 美國瓦里安儀器公司。
1.2 實驗方法
1.2.1 重組MDH的質粒構建和氨基酸序列分析P.bacterium1109為耐熱菌屬,其全基因組序列已公布在NCBI數據庫(GenBank:LDJB01000003),調取其已注釋MDH基因序列(GenBank:KLU39398.1)并委托上海捷瑞公司合成。MDH基因用限制性內切酶NdeI和XhoI雙酶切后,連接到同樣雙酶切的pET-22b(+)載體上,獲得重組質粒pET-22b-Peba-MDH,且重組基因C端加上6個組氨酸標簽便于目的蛋白的純化。具體實驗操作步驟參考已報道文獻[22]。
重組MDH與其它已報道MDHs的氨基酸序列比對和同源性分析,通過Clustal-Omega和Espript在線網址完成。
1.2.2 重組MDH的誘導表達和分離純化 將重組質粒導入到E.coliBL21(DE3)中,并涂布在固體LB培養基平板上(含Amp)37 ℃過夜培養12 h,由于pET-22b(+)載體上含有抗Amp基因片段,成功轉化的菌體會在平板上形成菌落。挑取成功轉化的重組菌的單菌落,接種到4 mL的LB液體培養基(含Amp)37 ℃搖床(200 r·min-1)過夜培養(12 h),然后將4 mL種子液轉移到200 mL的LB液體培養基中(含Amp),在此條件繼續下繼續培養到細胞光密度0.6~0.8(OD600)時,添加異丙基硫代半乳糖苷(IPTG)至終濃度為1 mmol/L,將搖瓶轉移至28 ℃搖床(200 r·min-1)誘導培養6 h。之后通過離心機(8000 r·min-1,5 min)收集菌體,用緩沖液(50 mmol/L Tris-HCl緩沖液,100 mmol/L NaCl,pH7.0)洗滌菌體兩次。將洗滌后的菌體進行超聲破碎,離心(12000 r/min,10 min)收集上清液。同時上清液用孔徑0.45 μm的水相濾膜過濾,使用填充有螯合Ni2+的層析柱進行純化[23]。
所有純化步驟均在4 ℃條件下進行。重組酶的純化分三個步驟[20],第一步:用5個柱體積的上樣緩沖液(50 mmol/L Tris-HCl,500 mmol/L NaCl,pH7.0)平衡柱子,將上述收集的上清液(粗酶)泵入裝有螯合Ni2+的層析柱;第二步:用洗滌緩沖液(50 mmol/L Tris-HCl,500 mmol/L NaCl,50 mmol/L咪唑,pH7.0)除去柱內雜蛋白;第三步:用洗脫緩沖液(50 mmol/L Tris-HCl,500 mmol/L NaCl,500 mmol/L咪唑,pH7.0)洗脫并收集目的蛋白。將收集的目的蛋白用含10 mmol/L EDTA的透析液(50 mmol/L Tris-HCl,pH7.0)透析6 h,去除金屬離子,最后用不含EDTA的透析液(50 mmol/L Tris-HCl,pH7.0)去除EDTA,透析完成后將目的蛋白溶液放在低溫冷藏。
1.2.3 重組MDH相對分子量和全分子量的測定 SDS-PAGE電泳檢測,分離膠和濃縮膠的丙烯酰胺以12%(w/v)和4%(w/v)比例配制。電泳結束后用考馬斯亮藍染色1~3 h至深藍色,然后用脫色液脫色至透明,通過蛋白酶條帶與Marker的對比,確定其相對分子量。
使用高效液相色譜(HPLC)檢測系統進行全分子量的測定,繪制全分子質量的標準曲線,根據標準曲線得出重組酶的全分子質量。
HPLC條件:流動相:0.1 mol·L-1Na2SO4,0.05% NaN3和0.1 mol·L-1KH2PO4;柱溫:25 ℃;流速:1 mL·min-1;紫外檢測器:280 nm。
1.3 重組MDH的酶學性質研究
1.3.1 重組MDH的酶活檢測 純酶溶液的蛋白濃度利用Lowry法檢測并計算[24]。使用紫外/可見分光光度計(紫外340 nm)通過測量NADH吸光值的變化量來測定重組MDH的酶活[23]。反應體系1 mL:50 mmol/L Tris-HCl(80 ℃,pH8.5),0.5 mmol/L NADH,50 μL純酶和100 mmol/L D-果糖。
比酶活:每毫克酶蛋白所含的酶活單位(U/mg)。
1.3.2 重組MDH最適pH和pH穩定性的測定 MDH是一種氧化還原酶,可以催化甘露醇和D-果糖之間的可逆反應。配制pH為6.0~9.5的Tris-HCl(50 mmol/L)緩沖液。
還原反應研究:將反應體系1 mL:50 mmol/L Tris-HCl,0.5 mmol/L NADH,50 μL純酶和100 mmol/L D-果糖,置于不同pH條件下進行反應,并將結果中的最高酶活設定為相對酶活100%。
氧化反應研究:將反應體系1 mL:50 mmol/L Tris-HCl,0.5 mmol/L NAD+,50 μL純酶和100 mmol/L甘露醇,置于不同pH條件下進行反應,并將結果中的最高酶活設定為相對酶活100%。
取pH分別為7.0、8.0、9.0的Tris-HCl(50 mmol/L)緩沖液各800 μL,分別加入200 μL純酶溶液,并將其置放于4 ℃條件下,分別每隔2 h取樣以D-果糖為底物測定酶活力變化,并分別將0 h測定的酶活設定為相對酶活100%。
1.3.3 重組MDH最適溫度和溫度穩定性的測定 同1.3.2研究重組MDH的還原反應和氧化反應。將pH8.5的還原反應體系(1 mL:50 mmol/L Tris-HCl,0.5 mmol/L NADH,50 μL純酶和100 mmol/L D-果糖)分別在55、60、65、70、75、80、85、90 ℃的水浴鍋環境中反應,并將結果中的最高酶活設定為相對酶活100%。將pH8.5的氧化反應體系(1 mL:50 mmol/L Tris-HCl,0.5 mmol/L NAD+,50 μL純酶和100 mmol/L甘露醇)分別在上述的水浴鍋環境中反應,并將結果中的最高酶活設定為相對酶活100%。
將純酶溶液置于75、85、90 ℃條件下保溫,定時取樣加入到1.3.1反應體系中,測定不同溫度取樣的酶活,并將初始酶活設定為相對酶活100%,利用一級動力學模型,作出相對酶活的圖像,研究重組MDH的熱穩定性。
1.3.4 金屬離子對重組MDH的影響 將金屬離子Co2+、Ba2+、Ni2+、Ca2+、Mg2+、Zn2+、Mn2+和Cu2+分別加入到1.3.1反應體系中,至終濃度為1 mmol/L。在最適條件下,將測得的酶活性進行比較,并將不添加金屬離子的一組設定為相對酶活100%。
1.3.5 重組MDH的底物特異性 將可能存在還原反應的不同底物(包括D-果糖、D-核糖、D-木糖、L-山梨糖、D-阿洛糖、D-甘露糖、D-半乳糖)分別添加到1 mL反應體系(50 mmol/L Tris-HCl,0.5 mmol/L NADH,50 μL純酶)中,至終濃度50 mmol/L,將測得的酶活性進行比較,其中底物為D-果糖的反應體系測定的酶活設定為相對酶活100%。
將可能存在氧化反應的不同底物(D-甘露醇、木糖醇、山梨醇、D-阿拉伯醇、半乳糖醇、阿洛糖醇)分別添加到1 mL反應體系(50 mmol/L Tris-HCl,0.5 mmol/L NAD+,50 μL純酶)中,至終濃度50 mmol/L,將測得的酶活性進行比較,其中底物為D-甘露糖醇的反應體系測定的酶活設定為相對酶活100%。
1.3.6 重組MDH以D-果糖為底物的動力學參數測定 使用不同濃度的D-果糖(10~200 mmol/L)在最適條件下測定酶活性,研究重組酶的動力學參數。利用Lineweaver-Burk雙倒數法繪圖,計算重組MDH的動力學參數,包括米氏常數(Km)、催化常數(kcat)、催化效率(kcat/Km)。
1.3.7 重組MDH在甘露醇生產中的應用 研究D-果糖轉化生產甘露醇,在pH8.5和溫度80 ℃條件下,加酶量為20 U/mL,加入NADPH的終濃度為200 mmol/L,使用初始濃度100、200和400 mmol/L的D-果糖作為底物,定時從反應體系中取樣,使用HPLC檢測甘露醇的生成量。
甘露醇檢測:將定時所取樣品離心(4 ℃,12000 r/min,5 min)取上清液,用0.22 μm微濾膜過濾,將過濾液稀釋進行HPLC檢測,根據反應體系產物與甘露醇標準樣品的出峰時間進行定性分析,根據反應體系與甘露醇標準樣品的出峰面積進行定量分析。檢測條件:Waters2695型高效液相色譜儀,Waters 示差折光檢測器(RI),Sugar-PakTM鈣離子色譜柱(10 μm,D7.8 mmol/L×300 mmol/L),流動相為0.22 μm 濾膜過濾后的超純水,柱溫85 ℃,流速0.6 mL/min。
1.4 數據處理
Clustal-Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/?tdsourcetag=s_pctim_aiomsg)和Espript(http://espript.ibcp.fr/ESPript/ESPript/)在線網址進行氨基酸序列比對分析。采用Origin 7.0 Pro進行試驗數據圖表分析,所有試驗至少重復三次,結果表示為平均值±標準偏差。
2 結果和討論
2.1 重組MDH的質粒構建和在E.coli BL21(DE3)中的表達
經測算重組基因片段全長1047 bp,可編碼349個氨基酸,翻譯蛋白理論相對分子質量為37.59 kDa(包含6個組氨酸)。經過重組蛋白的純化步驟,將純化的酶蛋白溶液進行SDS-PAGE蛋白電泳檢測(圖1),得到的蛋白條帶單一,分子量大小約為37 kDa,與理論值基本一致,表明重組MDH成功表達,純化效果較好。
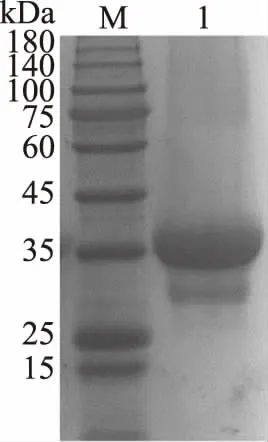
圖1 重組MDH純化后的SDS-PAGE電泳圖
使用裝備有紫外檢測器的TSKgel G2000SWXL凝膠色譜柱,測定來自P.bacterium1109的MDH的蛋白全分子質量。繪制全分子質量的標準曲線(圖2),根據標準曲線得出重組酶的全分子質量約為150 kDa,SDS-PAGE結果顯示該酶的單亞基相對分子質量約為37 kDa,表明一個酶分子含有四個單亞基,推測該酶是一個四聚體結構。目前已報道來源于C.hydrothermalis[25]的MDH是唯一的二聚體結構,來源于L.mesenteroides[14]、R.sphaeroides[26]的MDHs是單體結構,來源于C.magnoliae[16]、T.neapolitana[20]、A.bisporus[27]的MDHs為四聚體結構,其中來源于C.magnoliae[16]、T.neapolitana[17]的MDHs單亞基相對分子質量與本實驗的重組酶相近,為35~36 kDa。
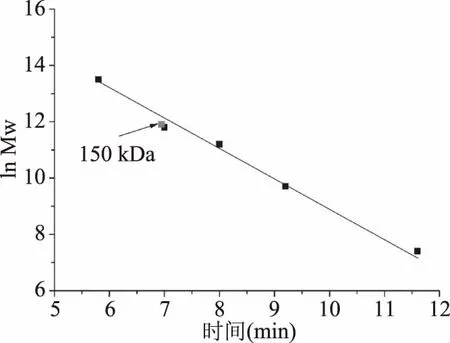
圖2 重組MDH全相對分子質量
2.2 重組MDH的氨基酸序列分析
目前,已經有十多種來自不同微生物的MDH編碼基因被克隆鑒定,并在NCBI官網中公布了蛋白質登錄號。表1顯示了來自不同微生物的MDHs的氨基酸序列同源性分析結果。來源P.bacterium1109的MDH和來源C.hydrothermali[25]的MDH有最高同源性(46.77%),其次與來源耐熱菌T.maritima[19]、T.neapolitana[20]的MDH同源性分別為32.38%和32.59%。此外,來源P.bacterium1109的MDH和來源R.sphaeroides[18]的MDH的同源性最低(17.94%)。雖然P.bacterium1109來源的MDH與大多數MDHs的同源性較低(<40%),但通過氨基酸序列比對鑒定其仍顯示出幾個關鍵殘基(圖3)。例如,協調大多數中鏈MDH與Zn2+結合的殘基(Cys37,His58,Glu59,Glu138,Cys142),催化殘基(Cys84,Cys87,Cys90)和輔因子結合殘基(Gly162,Gly164和Gly167),但通過P.bacterium1109 MDH與其它MDHs對比,發現R.sphaeroides[26]和P.fluorescens[20]的MDHs是例外,因為它們兩者均屬于長鏈MDH,不嚴格依賴Zn2+和其他金屬輔因子,不同于其它多元醇脫氫酶。
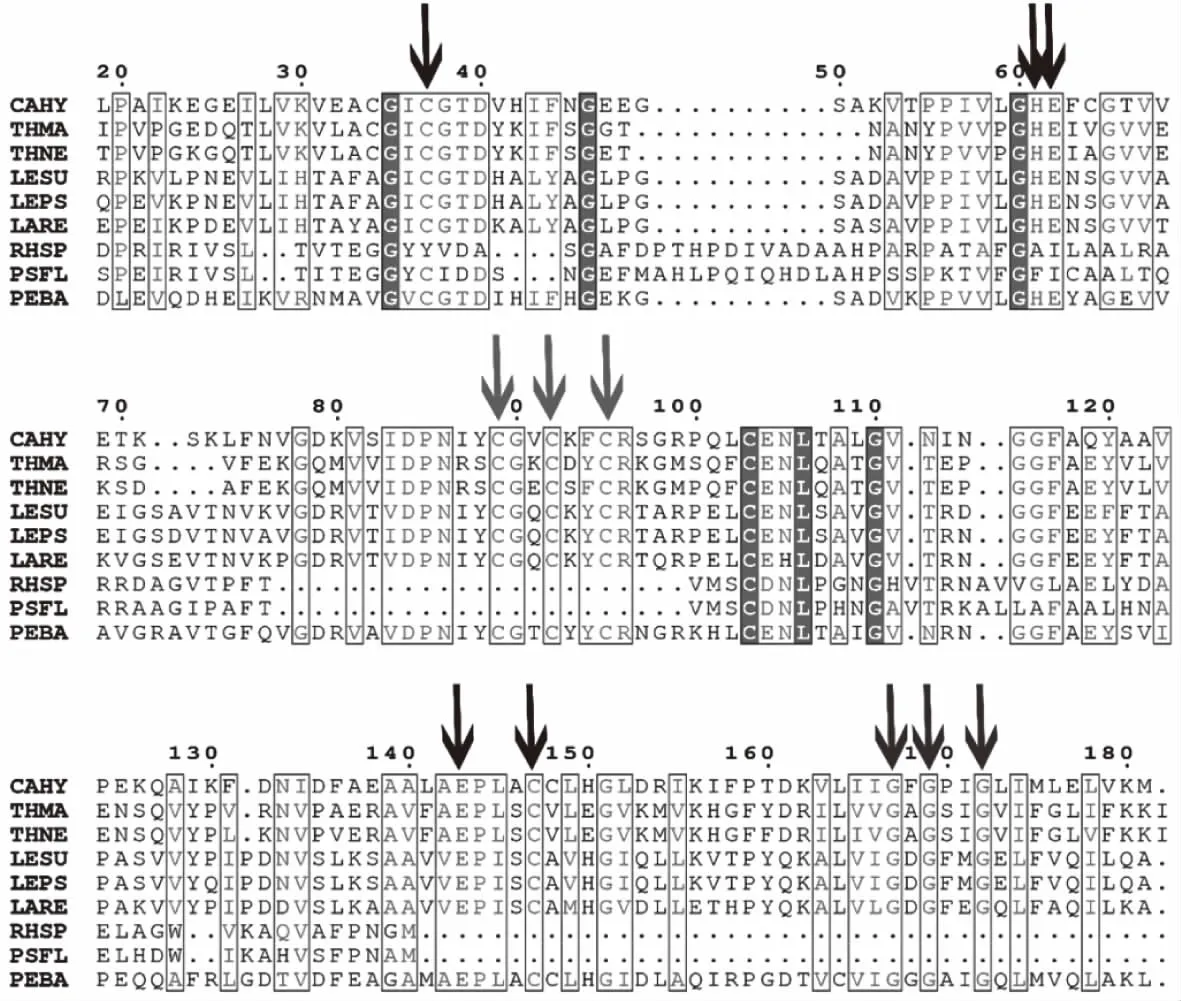
圖3 MDHs的氨基酸序列比對
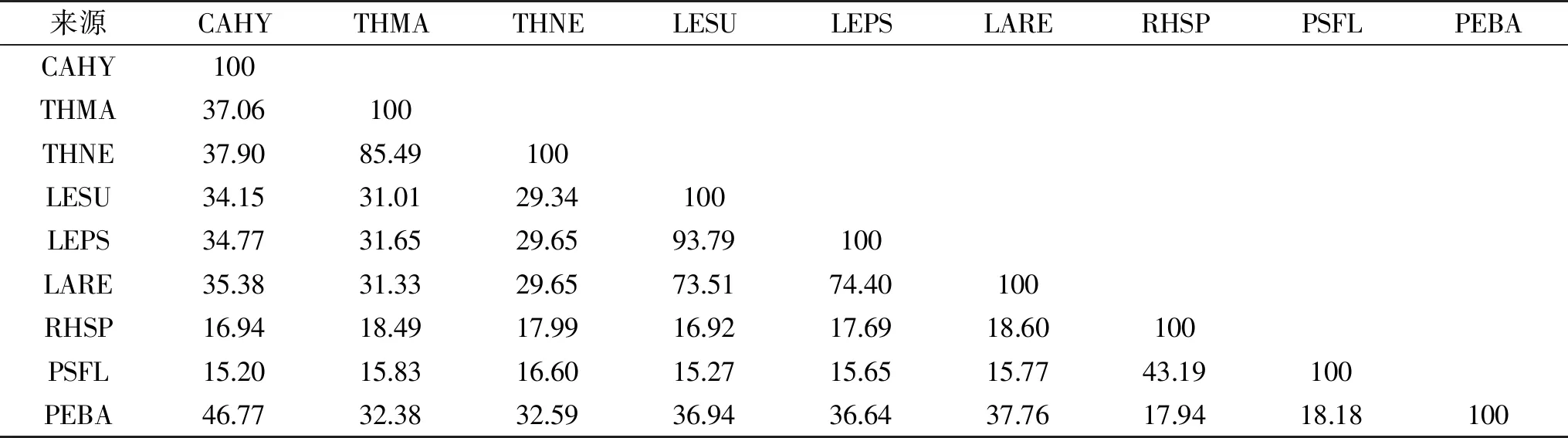
表1 不同來源MDHs的同源性對比(%)
大多數中鏈MDH都保留了用于協調金屬離子結合(顯示為黑色的向下箭頭)和催化作用(顯示為紅色的向下箭頭)和輔因子結合(顯示為藍色的向下箭頭)的殘基。
2.3 重組MDH的酶學性質鑒定
2.3.1 重組MDH的最適pH和pH穩定性 酶的催化活性受pH的影響很大。本文研究了來源于P.bacterium1109 MDH的酶活力在D-果糖還原和甘露醇氧化過程中受pH的影響。如圖4,來自P.bacterium1109的MDH,當以D-果糖和甘露醇為底物時,最適還原、氧化pH分別為8.5和9.0,表明該酶在堿性條件下比其他MDH顯示更高的活性,此外當以D-果糖為底物時,該酶在pH6.5的弱酸性條件下仍可保留接近50%的相對活性,表明該酶具有較寬的pH譜。在工業化大規模生產中往往要求酶具有寬的pH譜以適應體系環境的變化,在糖酶的催化反應中,略帶酸性的環境可以有效抑制碳水化合物的美拉德(褐變)反應[28],防止產生非酶褐變和減少副產物的產生[29-30]。在本文研究中,來源P.bacterium1109的MDH在pH分別為6.0和6.5的弱酸性條件下,表現出相對較高的活性,這有利于其在甘露糖醇工業生產中的應用。
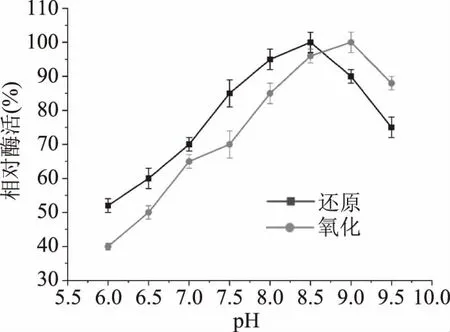
圖4 pH對重組MDH催化活性的影響
由圖5可知,重組酶在不同pH環境下的穩定特性。在長時間置于不同pH環境后,重組酶的活力變化不大,pH7.0條件下6 h,酶活為96%,pH8.0條件下6 h,保持92%的酶活力,在pH9.0環境下,酶活雖有下降,但6 h后仍能保持89%的活力,綜上所述該酶有比較好的pH穩定性。
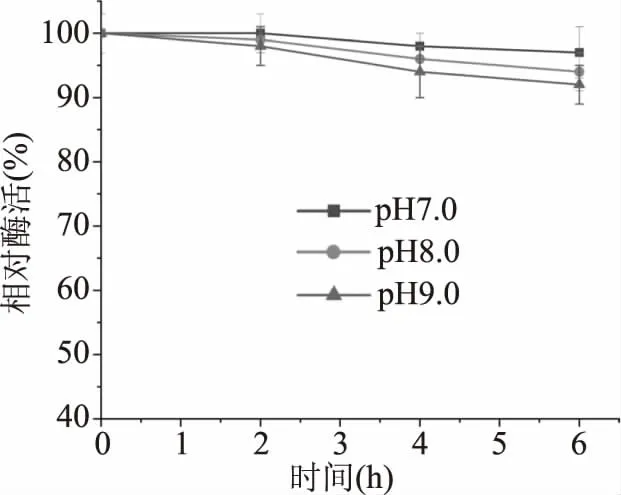
圖5 重組MDH的pH穩定性
2.3.2 重組MDH的最適溫度及溫度穩定性 溫度是影響酶催化效率的重要條件之一。如圖6,來自P.bacterium1109的MDH,當以D-果糖和甘露醇為底物時,最適還原和氧化溫度分別是80和85 ℃。當以D-果糖為底物時,在55~85 ℃間,該酶的催化活力隨著溫度的升高而逐漸增加。80 ℃時,酶活力達到最大。在70~90 ℃的范圍內保持最大活性的70%以上,表明該酶可以在較寬的溫度范圍內發揮其催化活性。相較于其它來源的MDHs,大多數MDHs的最適溫度都低于40 ℃,其中包括來源L.sanfranciscensis[10]、L.intermedius[11]和C.magnolia[16]的MDH,最適溫度分別是35、35和37 ℃,然而來自Thermotoga的兩個菌屬T.maritime[19]和T.neapolitana[20]的MDH具有較高的最適溫度,分別為95 和90 ℃(表4)。
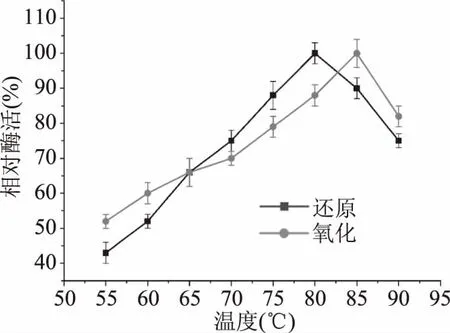
圖6 溫度對重組MDH催化活性的影響
由圖7可知,75 ℃條件下溫育6 h,保持大于80%的酶活。85和95 ℃條件下分別溫育6和4 h,仍然能分別保持52%和48%的酶活。由表2可知,在75、85、90 ℃條件下,來自P.bacterium1109的MDH的半衰期分別為28.5、5.3、3.0 h,另外兩個具有較高最適溫度的MDH,來自于T.maritime[19]和T.neapolitana[20]的MDH的半衰期分別是1.0、0.5、0.25和24.6、2.0、1.6 h,相較而言,來自P.bacterium1109的MDH有更長的半衰期,熱穩定性更好。綜上所述,該酶的熱穩定性較高,當酶在較高溫度下進行長時間反應時,良好的熱穩定性是實際應用中評估的重要標準。
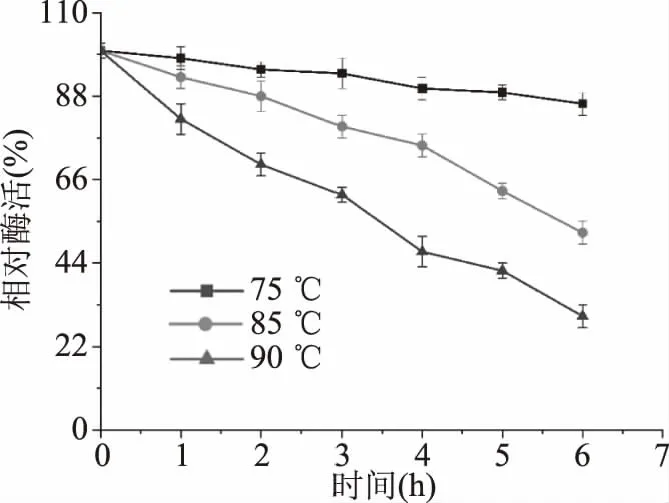
圖7 重組MDH的熱穩定性

表2 三種耐熱MDHs的熱穩定性比較
2.3.3 重組MDH的金屬離子的依賴性 金屬離子在催化過程中通常作為輔助因子或激活劑,在酶的催化反應中起著非常關鍵的作用。在作用于單糖的氧化脫氫酶中,金屬離子對酶的催化活力通常會有很大的影響[10]。本文研究了不同二價金屬離子對重組MDH催化活力的影響(圖8)。以純化后經EDTA處理的原始酶作為對照組,可以看出,金屬離子對酶的催化活力有比較大的影響。其中Cu2+、Zn2+、Ca2+、Mn2+都可以提高酶的催化活力,分別達到未添加金屬離子酶活的160%、260%、150%、170%。同時發現,添加Mg2+、Ba2+、Ni2+對酶活產生了不同程度的抑制,添加Co2+對酶活沒有顯著影響(P>0.05)。
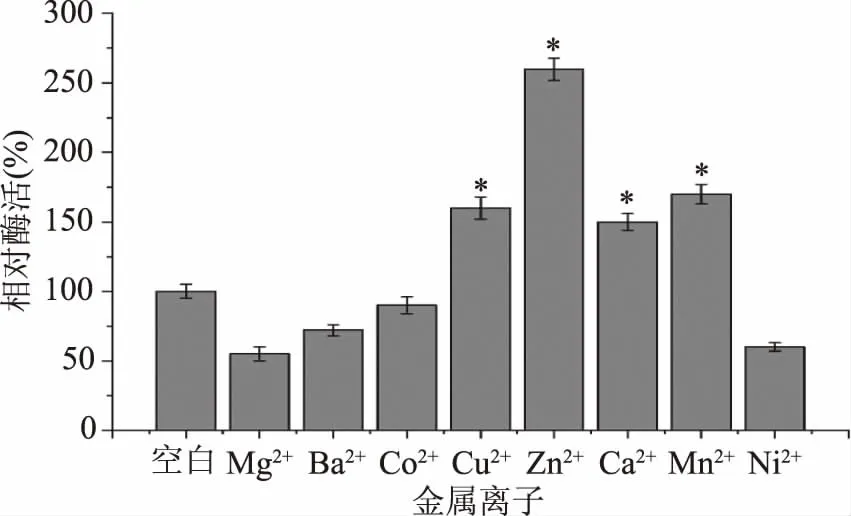
圖8 金屬離子對重組MDH催化活性的影響
MDH屬于中鏈多元醇脫氫酶,之前的研究表明Zn2+可能對其的活性有很大的影響,通過測定來自C.herbarum[31]的MDH晶體結構,證實晶體中的一些關鍵殘基被識別為金屬離子結合位點。由表4可知,目前報道的大部分MDH對金屬離子Zn2+有依賴型,Zn2+對酶的催化活力都有顯著提高。然而來自P.fluorescens[17]和R.sphaeroides[18]的兩個長鏈MDH沒有顯示出對金屬離子的依賴性。在本文研究中,Zn2+極大增強了MDH的酶活力,這表明酶結構中可能存在金屬離子的結合位點。
2.3.4 重組MDH的底物特異性研究 通過使用NADH和NAD輔因子,分別從還原和氧化兩方面研究重組MDH的底物特異性(表3)。當提供不同的己糖作為底物時,重組MDH對D-果糖具有較高的還原活性,比酶活為97 U/mg,對D-阿洛糖的活性很小,相對活性為17%,而對其他己糖(D-塔格糖、D-木酮糖和L-山梨糖等)沒有顯示任何活性。當與不同的己醇反應時,重組MDH對D-甘露醇表現出最大的氧化活性,并且對D-阿拉伯醇表現出一定的催化能力,相當于最大活性的25%。此次研究證明了來自P.bacterium1109的MDH有很強的底物專一性,并且重組MDH對D-果糖的還原活性高于目前報道的大多數MDH(表4)。
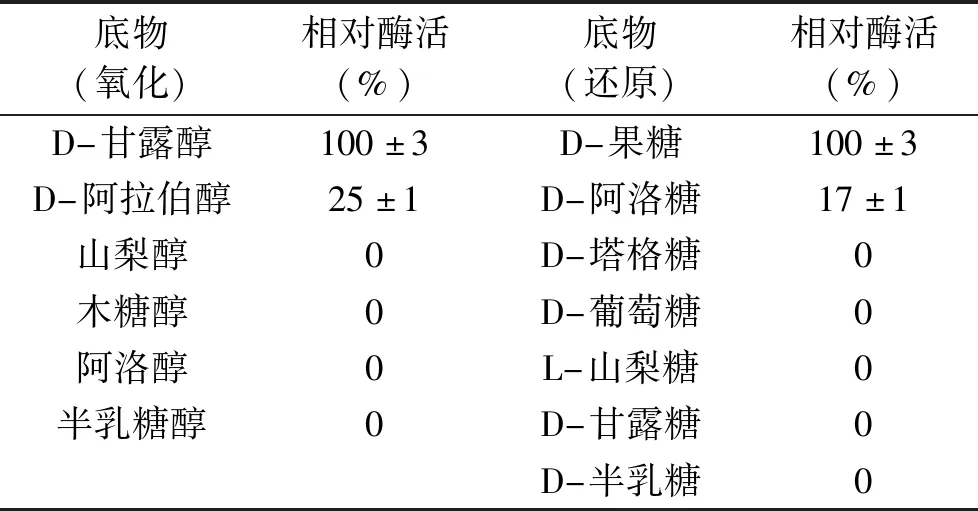
表3 重組MDH的底物特異性
2.3.5 重組MDH以D-果糖為底物的動力學參數測定 由表4可知,來自P.bacterium1109的MDH的Km值和kcat/Km值分別為20 mmol/L和7.5 L/(mmol·min)。相比而言,重組MDH和底物D-果糖的親和程度高于大多數MDHs。就其催化性能而言,到目前為止,已經報道的MDHs中(表4),以D-果糖為底物來自P.bacterium1109的重組MDH的催化性能也高于大多數MDHs,已知來自L.pseudomesenteroides[15]的MDH比酶活表現最高的是450 U/mg,但是其非常不穩定,在20 ℃條件下48 h,喪失了60%的酶活。在這項研究中,考慮到來自P.bacterium1109的MDH在底物親和度和催化性能表現出來的綜合能力,表明它在D-果糖工業生產甘露醇中有很高的價值。
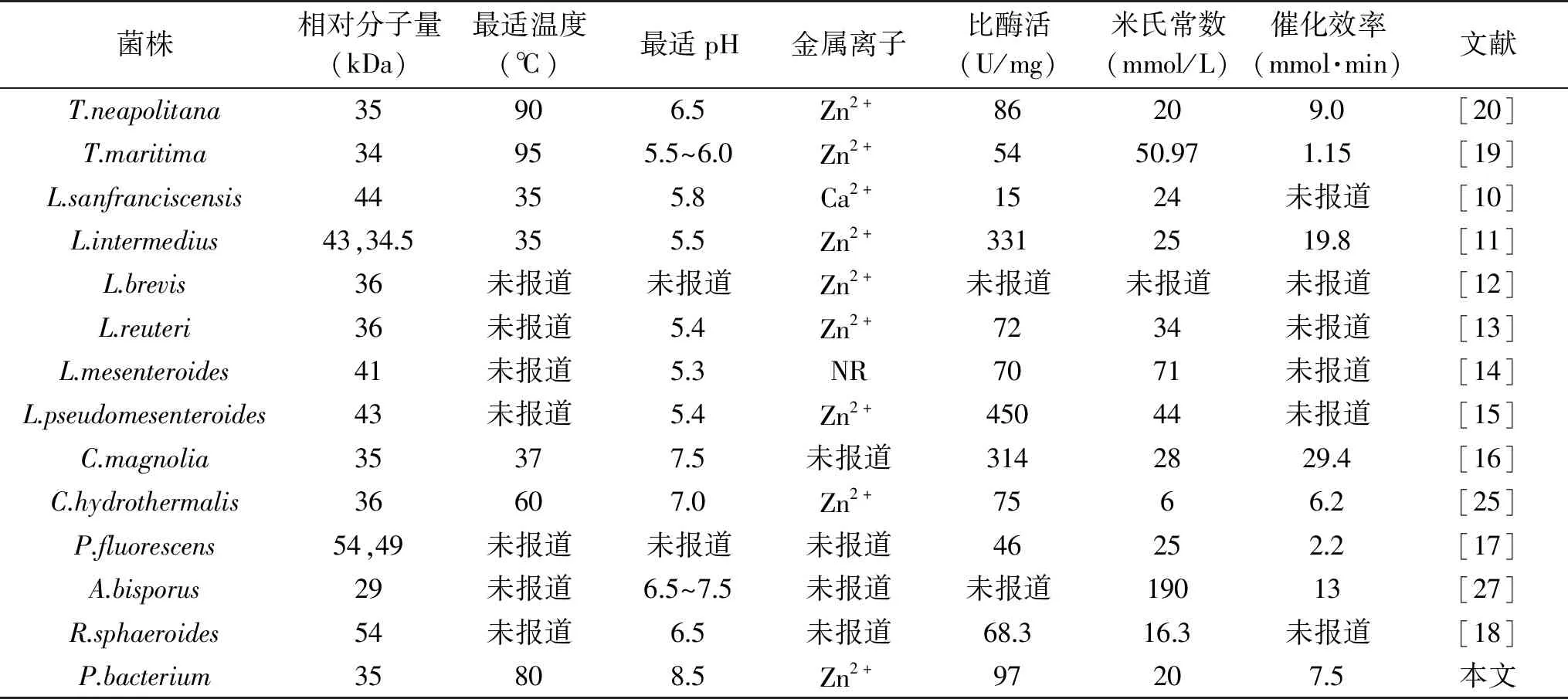
表4 不同來源MDHs的性質比較
2.3.6 重組MDH在甘露醇生產中的應用 研究甘露醇的轉化結果表明(圖9),在最適反應條件下,當以初始濃度100、200和400 mmol/L的D-果糖作為底物進行甘露醇轉化過程中,產率隨著時間的增加而增加,甘露醇的轉化率在最初的1 h內急劇增加,最終以100 mmol/L的D-果糖作為底物產率達到95%,以200、400 mmol/L的D-果糖作為底物產率也分別達到80%以上。值得注意的是,上述三個不同D-果糖濃度的反應體系中,甘露醇的轉化率都沒有達到100%,目前尚無100%轉化率的文獻研究,例如,來自L.mesenteroides[14]的MDH以100 mmol/L的D-果糖作為底物生產甘露糖醇達到了95%以上的轉化率,來自T.maritima[16]的MDH生產甘露糖醇的轉化率最高達到了80%以上。推測可能是輔酶因子NADH的不可逆消耗[32]和酶活力降低等原因導致。
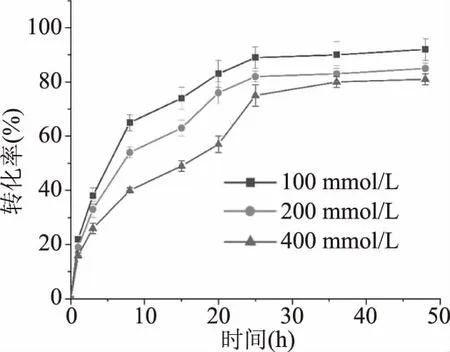
圖9 甘露糖醇的生物轉化
3 結論
本文鑒定了一株來源于耐熱菌屬P.bacterium1109的MDH,并可以應用于甘露醇的酶法合成。將來源于耐熱菌屬P.bacterium1109的MDH在大腸桿菌中表達,然后將表達的重組MDH目的蛋白進行分離純化,進行酶學性質研究。以D-果糖為底物時,重組表達的MDH的酶活為97 U/mg。該重組MDH最適pH和溫度分別是8.5,80 ℃,在堿性環境下有較好的熱穩定性,在75、85、90 ℃條件下,來自P.bacterium1109的MDH的半衰期分別是28.5、5.3、3.0 h。Zn2+存在時可以極大提高MDH的酶活力。在重組酶底物特異性的研究中發現,該酶對底物D-果糖有較強的親和力,并且具有相對窄的底物譜,底物專一性較高。在該酶的最適反應條件下,以400 mmol/L的D-果糖作為底物甘露醇產率達80%以上。總而言之,本文鑒定的重組MDH具有較高的熱穩定性和底物專一性等特點,在工業生產甘露醇中有很大的應用前景。
猜你喜歡
云南醫藥(2021年6期)2022-01-08 04:07:38
中國民間療法(2021年5期)2021-06-09 09:21:26
華聲文萃(2019年12期)2019-09-10 07:22:44
中西醫結合心血管病電子雜志(2017年17期)2017-11-17 15:54:41
上海農業學報(2017年4期)2017-04-10 12:40:22
中西醫結合心血管病雜志(電子版)(2017年17期)2017-01-11 06:37:25
食藥用菌(2016年6期)2016-03-01 03:24:27
中國繼續醫學教育(2015年4期)2016-01-07 07:38:11
護士進修雜志(2015年5期)2015-03-18 19:22:53
中國藥業(2014年24期)2014-05-26 09:00:09
