植物PHD蛋白的研究進展
2020-10-20 06:48:56姚依秀李玉林何艷軍高杰范敏
山東農業科學 2020年9期
姚依秀,李玉林,何艷軍,高杰,范敏
(1.浙江省農業科學院蔬菜研究所,浙江 杭州 310021;2.新疆農業大學林學與園藝學院,新疆 烏魯木齊 830052)
PHD(plant homeodomain)蛋白包含一個高度保守的PHD結構域,在基因的轉錄調控方面發揮重要作用。雖然關于動物中PHD蛋白的結構和功能研究已十分地廣泛和深入,但直到1993年,Schinder才首次在植物中發現并鑒定到PHD蛋白[1]。與動物相比,PHD基因的功能僅在少數模式植物中被研究。PHD蛋白與植物的各種生理生化過程有關,許多研究也表明了PHD基因在調節植物生長發育、病原體防御和對各種脅迫反應中具有重要作用[2]。先前的研究側重于PHD蛋白參與植物生長發育的調控,而關于其參與植物非生物脅迫及生物脅迫調控的報道較少。因此,對這一類重要的植物蛋白進行深入研究具有十分重要的意義。本文對植物PHD基因和蛋白的基本特征、在植物生長發育和逆境調控中的生物學功能及其相關的分子調控機制等進行綜述,并對今后的研究方向進行展望,以期為植物PHD基因家族在非生物(干旱、高鹽等)和生物(病原菌、害蟲等)脅迫等研究方面提供參考。
1 植物PHD蛋白的基本特征
PHD蛋白是一類真核生物中較為常見的具有一個或幾個PHD結構域的鋅指蛋白。PHD結構域由約60個氨基酸組成,具有Cys4-His-Cys3鋅結合基序特征[3],半胱氨酸殘基之間以及半胱氨酸與組氨酸之間的氨基酸數相對保守[4],且最后一對半胱氨酸前的第2個氨基酸殘基通常為色氨酸等芳香族氨基酸[5]。PHD結構域的三維結構通常為球狀[6]。進一步研究發現,PHD結構域是14種已知的鋅指結構域中的一種,存在于400多種真核生物蛋白質中,在進化過程中高度保守[3,7]。
不同植物中PHD家族的基因數目各不相同。如表1所示,大豆有45條[8]、水稻有59條[9]、毛果楊有73條[10]、擬南芥有70條[11]、胡蘿卜有106條[12]。大豆中PHD家族的基因數目最少,胡蘿卜中最多。蛋白長度范圍比較大,在74~2 336 aa之間。不同物種中PHD基因的內含子數各不相同,內含子數量在10個以內居多,也有個別物種基因中無內含子。等電點變化范圍為4.46~9.87,其中水稻(54%)和毛果楊(53%)中大部分為酸性蛋白。對PHD基因在擬南芥、水稻、大豆和毛果楊染色體上的分布進行分析(表2)發現,PHD基因都是不均勻地分布在各自物種的染色體上。植物PHD基因的重復事件共分為兩種,即串聯重復和大片段復制。其中擬南芥、水稻、毛果楊中基因的擴增主要依賴于大片段復制,在擬南芥和毛果楊中串聯重復基因只有兩對(表3)。
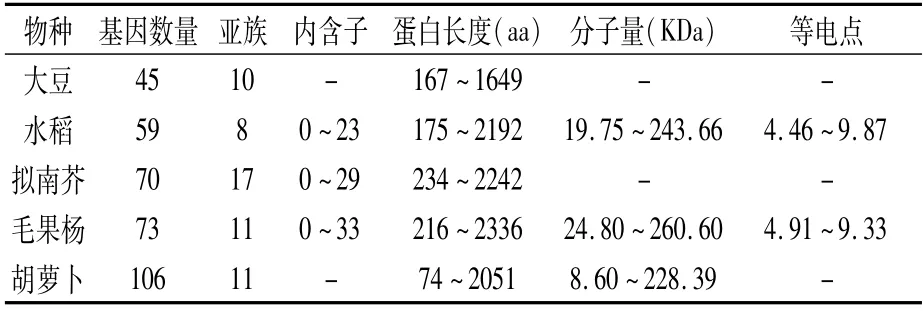
表1 擬南芥、水稻、大豆、胡蘿卜和毛果楊PHD家族基本信息

表2 擬南芥、水稻、大豆和毛果楊PHD基因在染色體上的分布
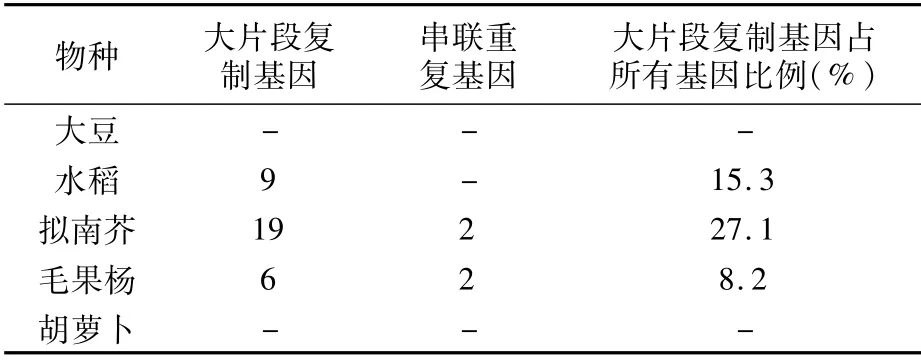
表3 大豆、水稻、擬南芥、毛果楊和胡蘿卜中PHD基因擴增情況
植物PHD蛋白結構比較復雜,除了共同擁有的PHD結構域外,同一物種中還包含其他多種結構域。如表4所示,擬南芥中有8種,水稻中10種,大豆中4種,胡蘿卜中3種,毛果楊中有6種。不同物種中PHD蛋白結構域類型也不同,但也有少數共有的結構域類型,如這5種物種中都具有PHD、BAH、DDT結構域。PHD蛋白中豐富且多樣的結構域很可能是導致其功能多樣性的決定因素。
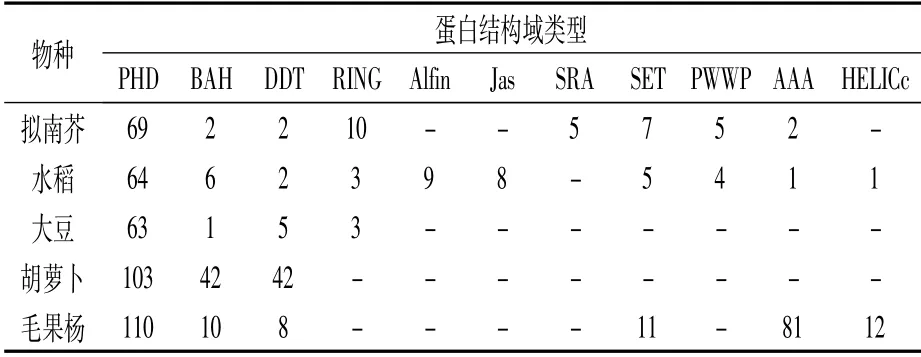
表4 擬南芥、水稻、大豆、胡蘿卜和毛果楊中 不同蛋白結構域的PHD蛋白數目
2 植物PHD蛋白的分子調控機制
2.1 植物PHD蛋白具有DNA結合能力
研究發現,植物PHD蛋白具有DNA結合能力,能夠在轉錄調控方面發揮作用。定位在細胞核上的雄性不育蛋白(MALE STERILITY 1,MS1)具有轉錄激活功能,在花粉絨氈層發育以及花粉壁生物合成中有重要作用,具有一個亮氨酸鋅指結構域和PHD 結構域。擬南芥MMD1、大麥MS1、玉米MS7和木薯MePHD1都是PHD結構域的轉錄因子。水稻PHD結構域鋅指蛋白OsTITANIA可以調控多種金屬離子轉運器基因的表達,在維持水稻正常生長發育中起重要作用[13]。木薯PHD結構域蛋白MePHD1對ADP-葡萄糖磷酸化酶亞基1(ADP-glucose pyrophosphorylase,AGPase)基因進行負調控,在淀粉代謝中發揮重要作用[14]。
2.2 植物PHD蛋白具有RNA結合能力
基因的轉錄后調控是真核生物適應生物和非生物脅迫的一種強有力的策略,該過程受到多種RNA結合蛋白(RNA-binding protein,RBPs)的調控,這些蛋白通過與靶mRNA的直接或間接相互作用,調控RNA代謝的多個方面,如RNA剪接、聚腺苷酸化、帽化、修飾、轉運、定位、翻譯和穩定性等。例如:PUF蛋白能與任何效應結構域結合,并且能夠選擇性結合一種特定的RNA目標,從而控制某一方面的新陳代謝作用。最早在果蠅和線蟲中分別發現的PUMILIO和FBF,并以此來命名的PUF蛋白就是這樣一類蛋白質[15,16]。它具有保守PHD基序,能夠識別高度保守的8~10個核苷酸核心基序,包括位于mRNA 3′UTR的UGUA核苷酸基序,以確保調控靶mRNA翻譯的穩定性和準確性。大多數已鑒定的PUF蛋白在哺乳動物、真菌、原生動物和植物的整個進化過程中都是保守的。在調控生長發育、逆境應答等多個方面發揮作用[17,18]。干旱脅迫誘導反義轉錄基因NFYA5,在擬南芥中被miR169靶向調控。NFYA5的反義轉錄基因蛋白NERF(NFYA 5 enhancing RING finger)具有PHD結構域的鋅指蛋白,可以通過重疊區域產生siRNA,并與miR169共同作用影響NFYA5轉錄。NERF蛋白作為泛素化的E3連接酶發揮作用,它的表達可以增強NFYA5的表達,并提高植物抗旱性[18]。
2.3 植物PHD蛋白具有組蛋白密碼解讀功能
各種組蛋白修飾(包括甲基化、乙酰化、磷酸化、泛素化等)的模式和組合的多樣化被稱為組蛋白密碼,與調節染色質狀態和基因轉錄活性有關。不同的組蛋白修飾狀態導致基因轉錄狀態的差異;PHD鋅指結構域能特異性識別組蛋白密碼,是組蛋白密碼的一種重要解讀器,具有招募下游被調控的效應物結合的能力,在調節染色質狀態和調控基因轉錄活性方面發揮作用[19]。
GmPHD5是大豆在鹽脅迫下組蛋白H3K4二甲基化與H3K14乙酰化互作的重要調控因子,在大豆抵抗非生物脅迫中發揮作用[20]。HBO1是組蛋白乙酰轉移酶(HAT)復合物的一部分,含有PHD結構域,支架式蛋白JADE1將HBO1與組蛋白H3-H4底物連接起來,在調節DNA復制、細胞增殖和發育方面發揮重要作用[21]。PHD結構域的泛素連接酶(E3s)與泛素偶聯酶(E2s)共同作用介導蛋白質泛素化,參與許多細胞過程[22]。
3 植物PHD蛋白的生物學功能
植物中的PHD基因家族成員眾多,蛋白結構各異,基因的表達特性也多種多樣,PHD蛋白已被證實在植物不同的生長發育和抗逆抗病等過程中發揮極其重要的作用。
3.1 PHD蛋白在植物生長發育中的生物學功能
研究表明,大量的PHD蛋白調控植物的生殖發育過程。GSR1編碼一個串聯的PHD蛋白,該基因可以通過生長素信號途徑調節種子萌發和休眠[23]。在擬南芥中,對PHD蛋白在春化作用途徑中功能的研究表明,FLOWERING LOCUS C(FLC)是開花網絡核心基因,VERNALIZATION INSENSITIVE 3(VIN3)是春化作用途徑上游的關鍵基因,長時間(大于20 d)的低溫處理會誘導VIN3的表達,并使FLC表達量降低[24]。PHD結構域可以結合2個鋅離子,以pygoupus蛋白為例,分別在C1、C2、H1和C5上結合第一個鋅離子以及在C3、C4、C6和C7上結合第2個鋅離子,使得在PHD結構域內形成2個環的穩定結構[19]。VIN3也具有與pygoupus蛋白類似的鋅蛋白交叉支架結構,該結構使得VIN3具有很多生理功能,例如參與蛋白與蛋白之間的相互作用、核小體組蛋白的修飾等。甲基化胞嘧啶結合蛋白VIM1編碼包含PHD、RING和SRA結構域的蛋白質,這些結構域共同存在于哺乳動物蛋白中,涉及染色質修飾、轉錄和細胞周期的調控,能與組蛋白結合,參與染色質狀態的調節[25]。小孢子母細胞減數分裂相關基因MMD1中存在一個PHD結構域,并且優先在雄性減數分裂細胞中表達,MMD1的突變表型和分子特征表明,它可能參與了染色質重構和/或通過減數分裂繼續進行轉錄,進一步說明了該蛋白可能調控減數分裂過程中相關基因的轉錄[26]。大麥PHD轉錄因子MS1在花粉發育中起關鍵作用[27]。此外,PHD基因家族還通過調節SOC1/FT染色質構象在調控開花過程中發揮作用[28]。
3.2 PHD蛋白在植物逆境脅迫中的功能
3.2.1 PHD蛋白在植物非生物逆境脅迫中的功能 qRT-PCR結果表明水稻PHD基因家族可能參與植物對不同環境脅迫的響應[29]。對水稻Os-PHD基因在脫落酸(abscisic acid,ABA)、高鎘和缺水下的表達模式進行檢測,結果表明,59個Os-PHD基因中有47個對高鎘脅迫有響應,只有5個基因響應低溫處理。大多數OsPHDs在高濃度的鎘、ABA和水分虧缺條件下表達水平上調。在玉米中,67個PHD基因中有15個響應干旱和高鹽脅迫[30];毛果楊中有9個PHD基因家族在鹽、干旱和冷脅迫下表現出差異表達[10]。
紫花苜蓿中的Alfin1蛋白是植物特異性PHD蛋白亞家族成員。研究表明,Alfin1是一種鹽誘導轉錄因子,可以調控MsPRP2基因的表達,從而提高植物耐鹽性[31,32],ALs(Alfin1-like PHD finger proteins)是擬南芥中的類Alfin1鋅指蛋白。ALs基因對鹽、低溫和氧化脅迫有不同程度的響應。擬南芥植株中過量表達AL5基因具有更強的鹽、干旱和低溫耐受能力,而AL5突變植株對這些脅迫的耐受能力降低[2]。大豆中GmPHD2基因在擬南芥中的異源表達提高了轉基因植株的耐鹽性[33]。也有研究表明PHD基因通過改變轉錄和閱讀表觀組蛋白修飾,在調控植物對非生物脅迫的反應中發揮重要作用[34,35]。逆境(干旱、高鹽和低溫)處理下,水稻內源基因OsPHD1的表達量明顯升高[35]。研究表明,OsMsr16作為PHD轉錄因子在植物響應鹽脅迫過程中發揮重要作用[36]。擬南芥中的PHD 蛋白可以與組蛋白H3K4me3/2結合[37],在鹽脅迫下誘導紫花苜蓿Aflin1和AL基因的表達[38,39]。大豆GmPHD5蛋白,作為甲基化H3K4的“編碼閱讀器”,調控乙酰化H3K14,從而控制鹽脅迫下靶基因的表達[20]。將來源于水稻的與耐逆性相關的PHDfinger家族轉錄因子基因KT496轉化水稻,可以顯著提高水稻對低溫、高鹽和干旱脅迫的耐受性[40]。
3.2.2 PHD蛋白在植物免疫反應調控中的作用
植物在生長發育過程中會面臨多種病原菌的侵染,在長期的進化中,植物演化出了兩道免疫防線來抑制病原菌的破壞。第一道防線是病原相關分子模式(pathogen-associated molecular patterns,PAMPs)所觸發的免疫反應(PAMP-triggered immunity,PTI)。植物使用模式識別受體(patternrecognition receptors,PRR)感知病原微生物的入侵并激發相應的抗病反應。植物PRR是表面定位的受體類蛋白(receptor-like proteins,RLPs)或受體激酶(receptor kinases,RKs)。植物受體激酶常富含亮氨酸的重復(leucine rich repeat,LRR),為LRR受體激酶(LRR receptor-like kinase,LRRRLK)。第二道防線是效應因子觸發的免疫反應(effector-triggered-immunity,ETI)。在ETI過程中,植物抗病基因編碼的抗病蛋白具有保守的核苷酸結合位點(nucleotide binding site,NBS)以及富含亮氨酸重復序列,它能夠直接或者間接的識別病原菌釋放的效應因子,引起效應因子觸發的免疫反應,使植物細胞主動死亡,即超敏反應(hypersensitive response,HR),進而抑制病原菌的侵染。植物NB-LRR R蛋白根據其N末端結構可以分為兩類,一類具有卷曲螺旋(CC)域CC-NBLRR,另一類具有Toll樣受體(TIR)域TIR-NBLRR[41]。PHD 蛋 白 既 可 與LRR 受 體 激 酶 如SERK3/BAK1、SnRK1類抗病蛋白互作,也可與TIR-NB-LRR、CC-NB-LRR類抗病蛋白互作,在植物免疫反應調控中發揮重要作用。
(1)PHD蛋白參與PTI反應:PHD蛋白與受體類激酶或受體類蛋白互作在植物免疫調控中發揮作用。PAMP觸發的防御反應需要受體蛋白、蛋白激酶(MAPK)、SUMO蛋白形成轉錄復合體,共同調節植物抗病免疫反應。具體通過三種方式來調節。
第一,PHD蛋白可以與SOBIR 1和SERK3/BAK1激酶互作。如模式植物擬南芥中富含亮氨酸重復的受體激酶 FLAGELLIN SENSING 2(FLS2)是模式識別受體(PRRs),FLS2受體感知細菌鞭毛蛋白并募集另一種共受體BAK1(brassinosteroid insensitive 1-associated kinase 1),BAK1和胞質激酶(BOTRYTIS-INDUCED KINASE-1,BIK1)形成有活性的共受體復合物,從而啟動擬南芥中的抗菌免疫。鞭毛蛋白可誘導SUMO蛋白與FLS2結合以觸發激酶BIK1釋放,也誘導脫SUMO化(deSUMOylating)酶Desi3a降解并增強FLS2 SUMO化修飾從而促進BIK1分解并觸發細胞內免疫信號。FLS2 SUMO化的破壞可以消除免疫反應,從而導致擬南芥對細菌性病原體的易感性[42]。
第二,PHD蛋白可以與活化的激酶C受體1(receptor for activated kinase C-1,RACK1)互作,活化的RACK1的SUMO化修飾導致RACK1B與AP2/ERF家族轉錄因子RAP2.6之間的相互作用增強[43]。OsRap2.6與RACK1A相互作用來促進水稻對稻瘟病的先天免疫[44]。
第三,PHD蛋白可以與AMP類蛋白激酶SnRK1(sucrose-non-fermenting 1)互作,SnRK1是AMP激活的蛋白激酶(AMP activated protein kinase,AMPK)。SnRK1蛋白激酶根據細胞外條件平衡細胞能量水平,是植物耐逆性的關鍵。SnRK1復合物在多個亞基上被SIZ1作為E3連接酶(含PHD結構域)SUMO化,SnRK1可以觸發其自身的SUMO化和降解,建立一個負反饋回路,該回路減弱了SnRK1信號傳導并防止了應激反應的有害過度激活[45]。
在感知病原體后植物固有的免疫受體激活各種信號傳導途徑,從而觸發宿主防御。PAMP觸發的防御信號傳導需要絲裂原激活的蛋白激酶途徑,該途徑可通過磷酸化調節轉錄因子的活性。這些轉錄因子也是SUMO結合的目標。SUMO偶聯決定了染色質修飾酶的募集和活性,從而控制基因表達。抗性蛋白信號傳導和SUMO結合都在轉錄復合體匯聚。例如,TIR-NB-LRR 蛋白SNC 1與組蛋白脫乙酰基酶HAD 19和轉錄共阻遏物Topless-related 1相互作用共同調控擬南芥對丁香假單胞菌抗病性,后兩者都是SUMO的靶標。SUMO偶聯可以將轉錄激活因子轉化為阻遏物,從而在沒有病原體的情況下阻止防御誘導[46,47]。
(2)PHD蛋白參與ETI反應:PHD蛋白與TIR-NBS-LRR、CC-NBS-LRR型等抗病蛋白互作在植物免疫調控中發揮作用。根據相關文獻報道,具體有以下三種互作方式參與調控。
第一,以蛋白與蛋白互作方式參與調控。擬南芥的SIZ1編碼一個SUMO E3連接酶,調控生物和非生物脅迫響應,SIZ1蛋白中PHD結構域對于其發揮功能至關重要,PHD能夠識別三甲基化組蛋白H3K4me3,對于組蛋白的識別以及轉錄抑制都很重要[48]。擬南芥防御調節者EDM2(enhanced downymildew 2)的PHD結構域可以識別三重修飾的組蛋白H3肽,通過影響免疫受體基因RPP7(resistance to peronospora parasitica)(RPP7編碼CC-NB-LRR蛋白)中另一個聚腺苷酸化位點處組蛋白H3(H3K9me2)的二甲基賴氨酸9的水平來動態調節RPP7表達水平,從而影響植株抗病能力[49]。擬南芥抗霜霉病基因RPP7編碼CC-NB-LRR蛋白,其抗性調節途徑與NDR1和SA 無關,但是RPP7轉錄水平控制需要EDM2。EDM2蛋白在結構上具有轉錄調節因子的典型特征,包括PHD結構域[50]。擬南芥EDM2除了與RPP7基因互作在抗霜霉病免疫調節中起作用外,還具有促進花型轉化的作用。EDM2在細胞核中可以與蛋白激酶WNK8相互作用調控植物葉和花發育[51]。在抗病過程中,EDM2是否與WNK8相互作用還不清楚。PHD 蛋白13(PHF13)是個與染色質有關的蛋白,PHF13是H3K4me2-3分子閱讀器和轉錄共調節子,可調節染色質結構和有絲分裂染色體濃縮,對于正確的細胞分裂很重要[52]。植物PHD結構域蛋白與病毒運動蛋白直接互作在抗病反應中發揮重要作用[53]。
第二,以蛋白-DNA互作方式(作為轉錄因子)參與調控。編碼致病相關蛋白2(pathogen defense-related,PR2)的歐芹pr2基因的轉錄可被真菌或細菌激發子激發調控。Pr2啟動子內有一個125 bp的區域,該區域包含真菌誘導子介導的表達所需的重要順式調控元件,而且包含的11 bp DNA基序(CTAATTGTTTA)可與歐芹和擬南芥核蛋白提取物中均存在的因子特異性結合。該11 bp DNA基序是PHD蛋白體內潛在的靶位點。研究表明,PHD-DNA相互作用在調節pr2基因表達中發揮了重要作用[54]。
第三,以蛋白-RNA互作的方式參與調控。模式植物擬南芥中APUM 5通過CMV RNA的直接結合特異性調節CMV侵染[55]。水稻稻瘟病抗性基因簇Pigm編碼多個CC(coiled-coil)類型的NLRs,具有廣譜抗病功能。與PigmR互作的蛋白PIBPs(PigmR-interacting and blast resistance proteins,PIBPs)是含有RRM(RNA-recognition motif)結構域的轉錄因子,能與PigmR基因特異性互作。PIBP 1正調控PigmR,激活下游免疫基因的表達[56]。
4 展望
近年來,人們對植物中PHD蛋白的結構、生物學功能、作用機制等進行了研究,結果發現其不僅參與調控植物的多種生命過程,還參與植物對干旱、鹽、溫度和各種病害等非生物和生物脅迫應答。這些研究為提高作物產量以及適應各種脅迫提供了嶄新的思路,具有良好的應用前景。但是關于PHD蛋白的研究仍存在一定的不足。第一,PHD蛋白的結構多種多樣,除含有保守的PHD結構域外,還有其它多種不同類型的結構域,這些是其功能多樣性的基礎。但除了對PHD結構域有所研究外,對于其他各個結構域的功能有待進一步深入研究。第二,PHD蛋白被證實能夠與DNA和RNA結合,在組蛋白密碼解讀等水平發揮調控作用,這也暗示了PHD基因調控機制的多樣性。PHD蛋白作用機制的研究多集中在蛋白-蛋白調控水平,對其在DNA、RNA結合以及組蛋白密碼解讀等水平介導的調控網絡的研究較少,對相關作用機理需要進一步試驗驗證,對這些方面展開深入的研究具有十分重要的意義。第三,對PHD蛋白的功能研究大多集中在植物在應對一些非生物脅迫如干旱、鹽堿等逆境,而對于病害等生物脅迫相關的研究還很少。在今后的研究中,進一步探索PHD蛋白的調控機制可為作物抗病抗逆性狀的改良提供更為詳細的理論指導。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
