添加硫和錳對長白山森林土壤與腐殖質頑固性有機碳礦化的影響*
2020-09-16 04:54:56王玉哲張鳳麗鄭興波韓士杰鄭俊強
林業科學 2020年8期
王玉哲 張鳳麗 鄭興波 韓士杰 鄭俊強,
(1. 福建農林大學林學院 福州 350002; 2. 中國科學院森林生態與管理重點實驗室 中國科學院沈陽應用生態研究所 沈陽 110016; 3. 河南大學生命科學學院 開封 475004)
土壤碳庫是陸地生態系統中最大的碳庫,其中森林土壤有機碳約占全球土壤總有機碳的70%(Panetal., 2011)。根據周轉時間不同,土壤有機碳可分成不同組分,其中蛋白質、淀粉和碳水化合物等容易被微生物利用和降解,稱為活性碳庫(Bergetal., 2007)。木質素、腐殖質類和多酚化合物等頑固性碳組分的周轉速度較慢,容易在土壤中累積,故稱為頑固性有機碳,鑒于其在全球碳循環中的重要性,對其礦化的關注愈來愈多(Davidsonetal., 2006)。
近年來,工業生產將大量的二氧化硫和氮氧化物排入大氣,之后不同形態的硫和氮隨降水(濕沉降)或顆粒物(干沉降)進入環境,引起一系列生態環境問題(Duanetal., 2016)。我國已成為繼歐美之后的世界第三大酸沉降(包括氮沉降和硫沉降)區(Liuetal., 2013)。硫沉降往往在酸沉降中占據很大比例,會引起土壤酸化,并通過改變微生物的豐度和活性從而影響土壤有機碳礦化(Vileetal., 2003)。此外,硫酸鹽的還原過程也會產生CO2(Chambersetal., 2011)。目前國內關于硫對土壤有機碳礦化影響的研究主要集中在熱帶和亞熱帶森林(吳建平等, 2015; Wuetal., 2016),而對溫帶森林的研究相對較少。
Mn在土壤中含量豐富(Boudissaetal., 2006),自然本底濃度能達到5 mg·g-1(Hernandez-Sorianoetal., 2012)。錳過氧化物酶(MnP)是一種依賴Mn的過氧化物酶,有助于白腐菌將木質素降解為CO2和水(Hofrichter, 2002)。有研究報道凋落物在分解后期的質量損失與其Mn2+濃度呈正相關(Bergetal., 2007; Hofrichter, 2002; Virzo De Santoetal., 2009)。此外,Trum等(2011)對歐洲山毛櫸(Fagussylvatica)林土壤腐殖質層的研究發現,Mn2+對土壤有機碳礦化的影響因腐殖質類型及分解程度而異。在未來全球變化背景下,大氣CO2濃度升高和酸沉降導致的土壤酸化也會引起土壤Mn2+濃度升高(Bowmanetal., 2008; Ohetal., 2004; Watmoughetal., 2007)。然而,土壤Mn2+濃度增加是否能促進土壤頑固性有機碳的分解,還需進一步研究(Trumetal., 2011)。
鑒于此,本研究以長白山自然保護區的闊葉紅松(Pinuskoraiensis)林、白樺(Betulaplatyphylla)和山楊(Populusdavidiana)為主的楊樺林和高山苔原的土壤以及闊葉紅松林和楊樺林的腐殖質為對象,探討硫和錳添加對土壤頑固性有機碳礦化及其溫度敏感性的影響,以期為評估長白山森林碳元素的生物地球化學循環對大氣硫輸入的響應提供科學依據。
1 材料與方法
1.1 供試土壤來源
供試土壤采自長白山自然保護區的闊葉紅松林、楊樺林和高山苔原。該區域屬于典型的溫帶大陸性季風氣候,年均氣溫3.8 ℃,年降水量600 ~ 900 mm,水熱同期,夏季短暫多雨,冬季漫長寒冷。長白山氣象觀測場監測數據表明,冬季(10月至翌年2月)降水中的硫酸根月均含量從2015的3.0 mg·L-1增加到2017年的4.0 mg·L-1,說明長白山森林地區硫沉降有一定程度的增加趨勢。闊葉紅松林位于中國科學院長白山森林生態系統定位站一號標準地(128°05’E,42°24’N),海拔766 m,主要樹種為紅松、蒙古櫟(Quercusmongolica)、水曲柳(Fraxinusmandsurica)和紫椴(Tiliaamurensis),優勢樹種的平均年齡為300年。楊樺林與闊葉紅松林毗鄰,于20世紀40年代林地干擾后形成。闊葉紅松林和楊樺林土壤為砂壤土,屬于暗棕壤,由松散的火山灰巖發育而成,0~5 cm土層的土壤有機碳含量約為13%,闊葉紅松林和楊樺林0~20 cm土層土壤的有效錳含量分別為1.20~1.59和0.81~1.22 mg·kg-1。高山苔原(128°04’E,42°02’N,海拔2 150 m),坡向朝南,有機層厚10~15 cm,透水性好,pH值為5.9。有機層下礦質土壤非永久凍土,主要植被為越桔屬(Vacciniumspp.)矮灌和苔蘚。
1.2 土樣采集
2010年5月,在闊葉紅松林和楊樺林分別設置20個采樣點,在高山苔原設置16個采樣點。在每個采樣點,分別于1 m2的區域用直徑5 cm的取土器采集3個0~10 cm土層土柱,將每個采樣點的3個土柱裝入同一塑料密封袋,放入冰盒中冷藏保存。土壤運回實驗室后,移去根系和石塊等雜質后,過2 mm篩,將每個植被類型所有土壤樣品混合均勻,取部分土壤樣品風干后用于測定基本理化性質,其余土壤置于4 ℃下冷藏保存用于培養試驗。在闊葉紅松林和楊樺林長期監測標準樣地內隨機選擇20個1 m×1 m樣方,移去樣方中心處地表凋落物層后,用10 cm×10 cm的不銹鋼采用框采集腐殖質層樣品(Fisheretal., 2000),過4 mm篩后分成2份,1份自然風干用于全碳和全氮含量測定,另一份置于4 ℃下冷藏保存以備室內培養試驗使用。供試土壤和腐殖質層的基本性質見表1。
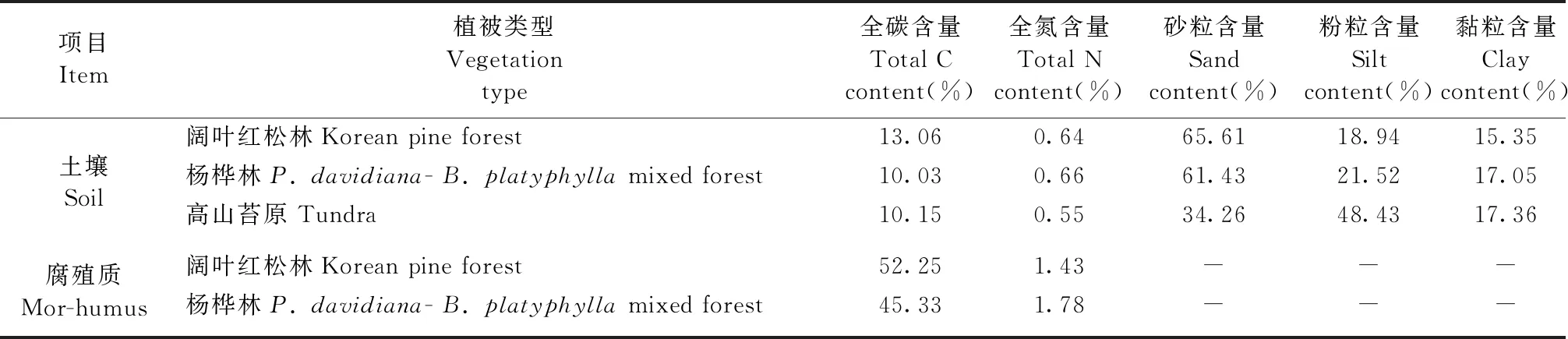
表1 供試腐殖質和0~10 cm土層土壤基本理化性質
1.3 試驗設計
稱取每種植被類型4 ℃冷藏保存的土壤(20 g)和腐殖質(7 g)樣品各50份,置于180 mL的培養瓶中,在黑暗條件下預培養90天。為保持好氧條件,分別將土壤和腐殖質的含水率調至其生長季的野外平均水平,分別為40%和70%。預培養是為了消耗樣品中的活性碳組分(Hoosbeeketal., 2007),在預培養過程中通過定期稱質量來維持樣品含水率。預培養結束后,用微型噴霧器將2 mL MnCl2、NaCl、MnSO4和Na2SO4溶液均勻噴灑在培養瓶中土壤和腐殖質樣品表面,每種溶液處理設10個重復。本研究供試土壤和腐殖質的Mn2+含量為1.12~3.90 mg Mn·g-1有機碳,根據土壤和腐殖質樣品中Mn2+添加量為3.00 mgMn·g-1有機碳的標準(Bergetal., 2007; Virzo De Santo, 2009),計算MnCl2和MnSO4溶液濃度,然后根據MnCl2和MnSO4溶液中Cl-和SO42-的摩爾濃度分別推算出NaCl和Na2SO4溶液濃度。闊葉紅松林、楊樺林和高山苔原土壤樣品添加的MnCl2溶液濃度分別為8.96、6.88和6.97 g·L-1、添加的NaCl溶液濃度分別為16.67、12.80和12.96 g·L-1、添加的MnSO4溶液濃度分別為10.76、8.26和8.36 g·L-1、添加的Na2SO4溶液濃度分別為10.12、7.77和7.86 g·L-1。闊葉紅松林腐殖質樣品添加的MnCl2、NaCl、MnSO4和Na2SO4溶液濃度分別為12.55、23.34、15.06和14.16 g·L-1; 楊樺林腐殖質樣品添加的MnCl2、NaCl、MnSO4和Na2SO4溶液濃度分別為10.89、20.25、13.07和12.29 g·L-1。對照處理中加入等量的雙蒸水。所有溶液的pH值調至7后再加入到土壤和腐殖質樣品中。添加溶液后,立即用Parafilm 封口膜覆蓋培養瓶來避免水分蒸發。將每個處理的10個重復分為兩組(每組5個),一組放至25 ℃培養,另一組在35 ℃下培養。分別于第1、3、6、10、15、21、30天用注射器采集氣體,每次氣體采集前用橡膠塞密封培養瓶 4 h。在第10天往所有培養瓶中加入1 mL的雙蒸水,以便去除水分對碳礦化的限制。用氣相色譜儀(Agilent 7 890 A,美國)測定所采集氣體的CO2濃度,并計算出單位時間CO2釋放量,用以表示有機碳礦化速率。用相鄰2次采樣時間的有機碳礦化速率均值代表這一時段的平均礦化速率,將平均礦化速率乘以培養時間得到該時間段的CO2釋放量,將所有采樣時間段的CO2釋放量相加得到CO2累積釋放量,用以表示土壤和腐殖質頑固性有機碳累積礦化量。采用磷脂脂肪酸生物標記(PLFA)法測定培養結束時(第30天)土壤和腐殖質樣品的磷脂脂肪酸總量,用于表征土壤微生物生物量。
1.4 數據處理
溫度敏感性系數Q10的計算公式如下(Kirschbaum, 1995):
Q10=(R2/R1)10/(T2-T1)。
式中:R2和R1分別為T2和T1培養溫度下土壤和腐殖質頑固性有機碳平均礦化速率。
采用重復測量方差分析方法檢驗錳和硫添加對土壤和腐殖質有機碳礦化速率的影響,用雙因素方差分析測定植被類型和錳、硫添加對土壤和腐殖質頑固性有機碳累積礦化量和Q10的影響,用單因素方差分析檢驗錳和硫添加對同一植被類型土壤或腐殖質頑固性有機碳累積礦化量、Q10和磷脂脂肪酸總量的影響,采用LSD方法進一步對比兩兩處理之間的差異。統計分析在SPSS 22.0(SPSS Inc. Chicago, IL, USA)軟件中完成,顯著性水平設為α=0.05。
2 結果與分析
2.1 土壤和腐殖質頑固性有機碳礦化速率
闊葉紅松林、楊樺林和高山苔原3種土壤頑固性有機碳礦化速率存在顯著差異(P<0.01,表2),而不同植被類型腐殖質頑固性有機碳礦化速率沒有顯著差異(P> 0.05,表2)。雙因素方差分析結果顯示,在MnCl2和NaCl處理之間,以及在 MnSO4和Na2SO4處理之間,土壤和腐殖質頑固性有機碳礦化速率均沒有顯著差異,因此,圖1只呈現MnCl2、MnSO4和對照3種處理土壤和腐殖質頑固性有機碳礦化速率。MnCl2和MnSO4處理之間的土壤頑固性有機碳礦化速率存在顯著差異(P<0.05),MnSO4處理能提高土壤頑固性有機碳礦化速率,而MnCl2對土壤頑固性有機碳礦化速率沒有顯著影響。
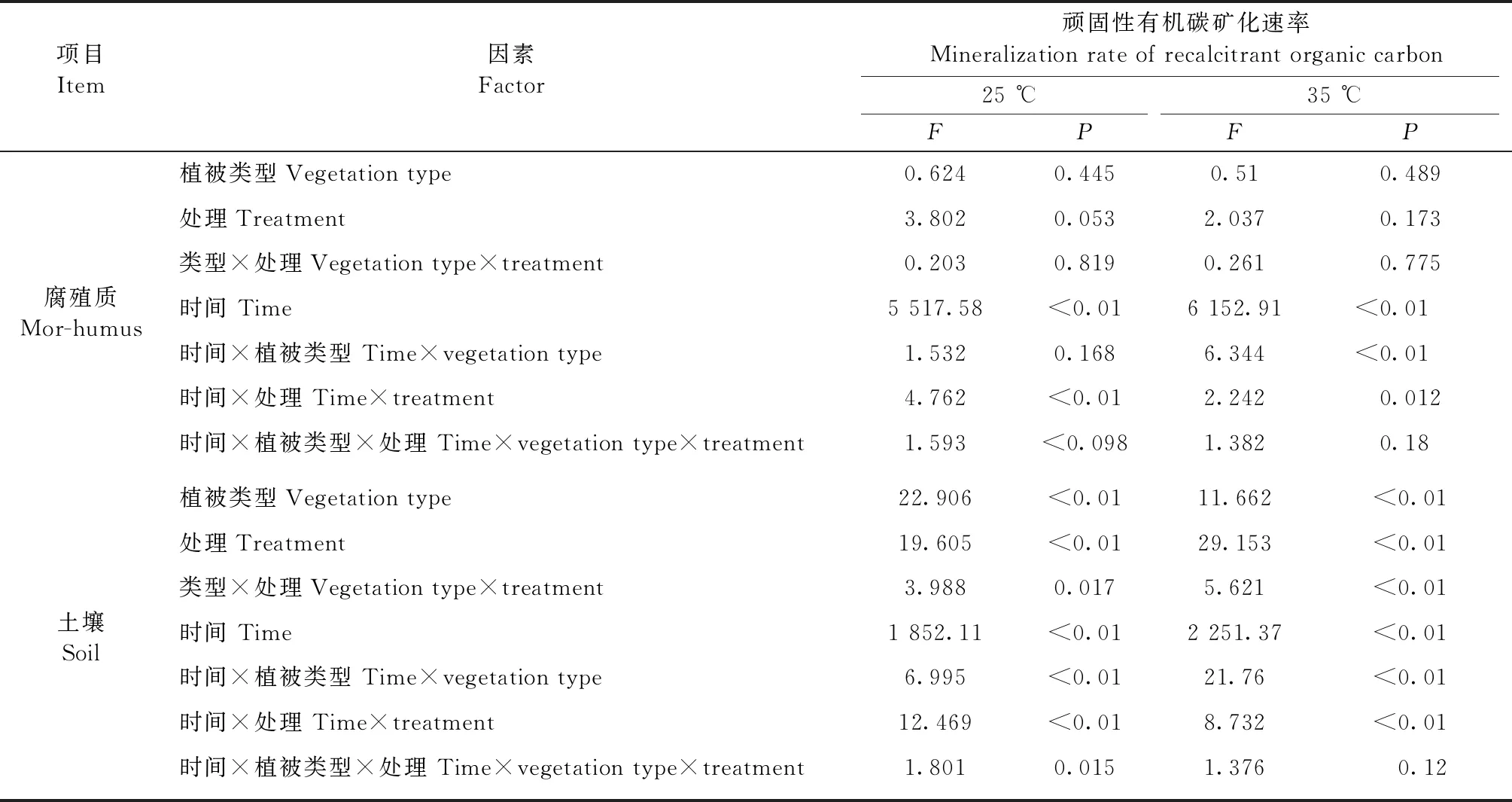
表2 土壤和腐殖質頑固性有機碳礦化速率的重復測量方差分析結果
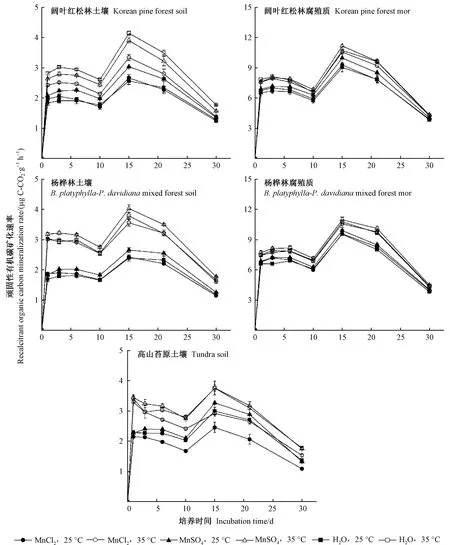
圖1 長白山森林土壤和腐殖質頑固性有機碳礦化速率
2.2 土壤和腐殖質頑固性有機碳累積礦化量
在MnCl2和NaCl處理之間以及在MnSO4和Na2SO4處理之間,土壤和腐殖質頑固性有機碳累積礦化量均沒有顯著差異(P> 0.05,圖2)。在培養溫度為25 ℃時,MnSO4和Na2SO4處理顯著提高了楊樺林的土壤和腐殖質頑固性有機碳累積礦化量(P<0.05,圖2)。在培養溫度為35 ℃時,MnSO4和Na2SO4處理顯著提高楊樺林土壤的頑固性有機碳累積礦化量,相反降低闊葉紅松林土壤的頑固性有機碳累積礦化量(P<0.05,圖2)。
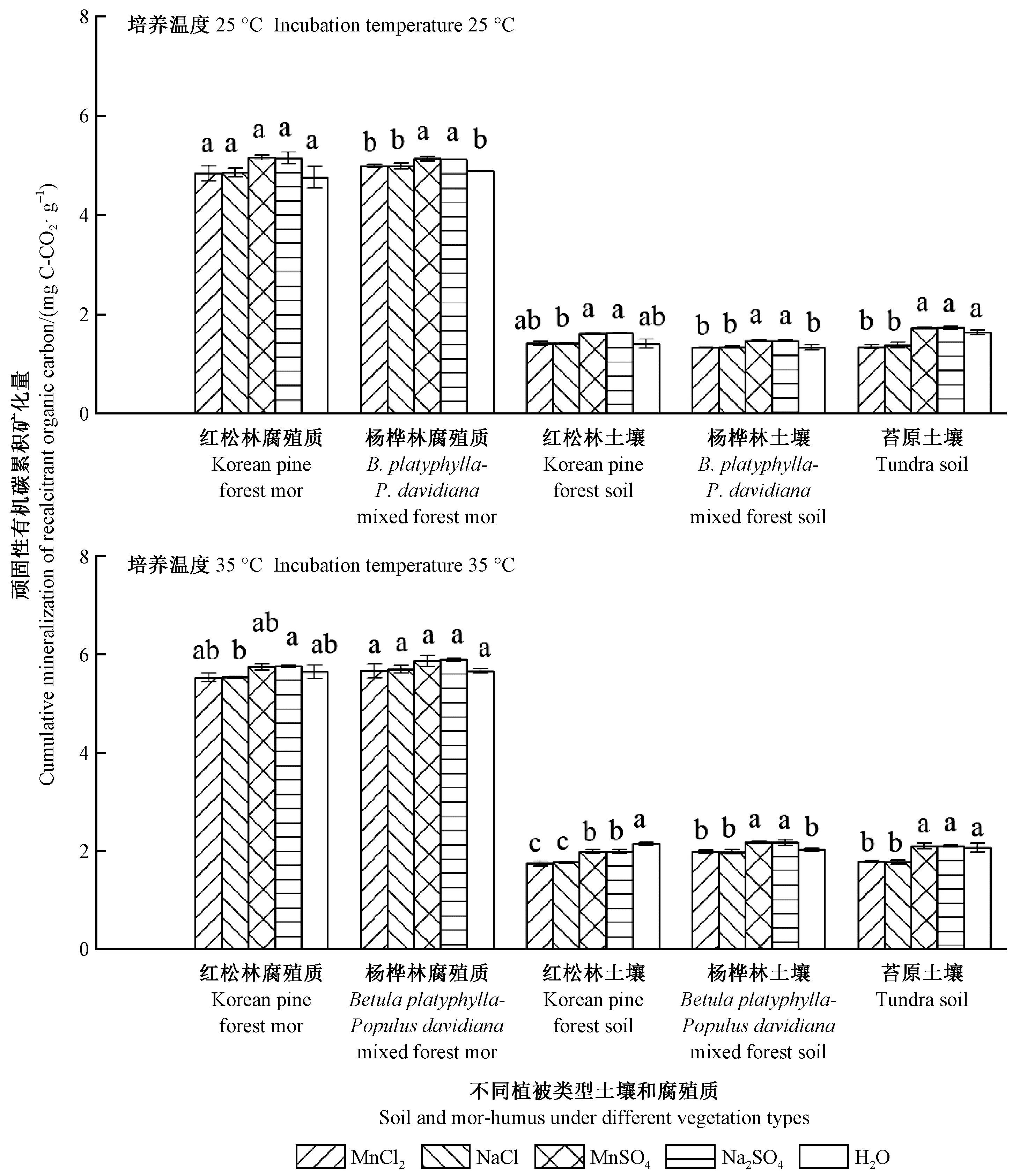
圖2 長白山森林土壤和腐殖質頑固性有機碳累積礦化量
2.3 土壤和腐殖質頑固性有機碳礦化速率的溫度敏感性
圖3表明:土壤和腐殖質頑固性有機碳礦化速率的Q10為1.1~1.6; MnCl2、NaCl、MnSO4和Na2SO4添加對長白山闊葉紅松林和楊樺林的腐殖質頑固性有機碳礦化速率Q10均沒有顯著影響(P> 0.05;MnCl2、NaCl、MnSO4和Na2SO4添加對土壤頑固性有機碳礦化速率Q10的影響因森林類型而異,其中顯著降低闊葉紅松林土壤頑固性有機碳礦化速率Q10(P<0.05),而對楊樺林和高山苔原的土壤頑固性有機碳礦化速率Q10沒有顯著影響; 對于高山苔原土壤,MnCl2添加處理的頑固性有機碳礦化速率Q10顯著高于MnSO4和Na2SO4處理(P<0.05)。
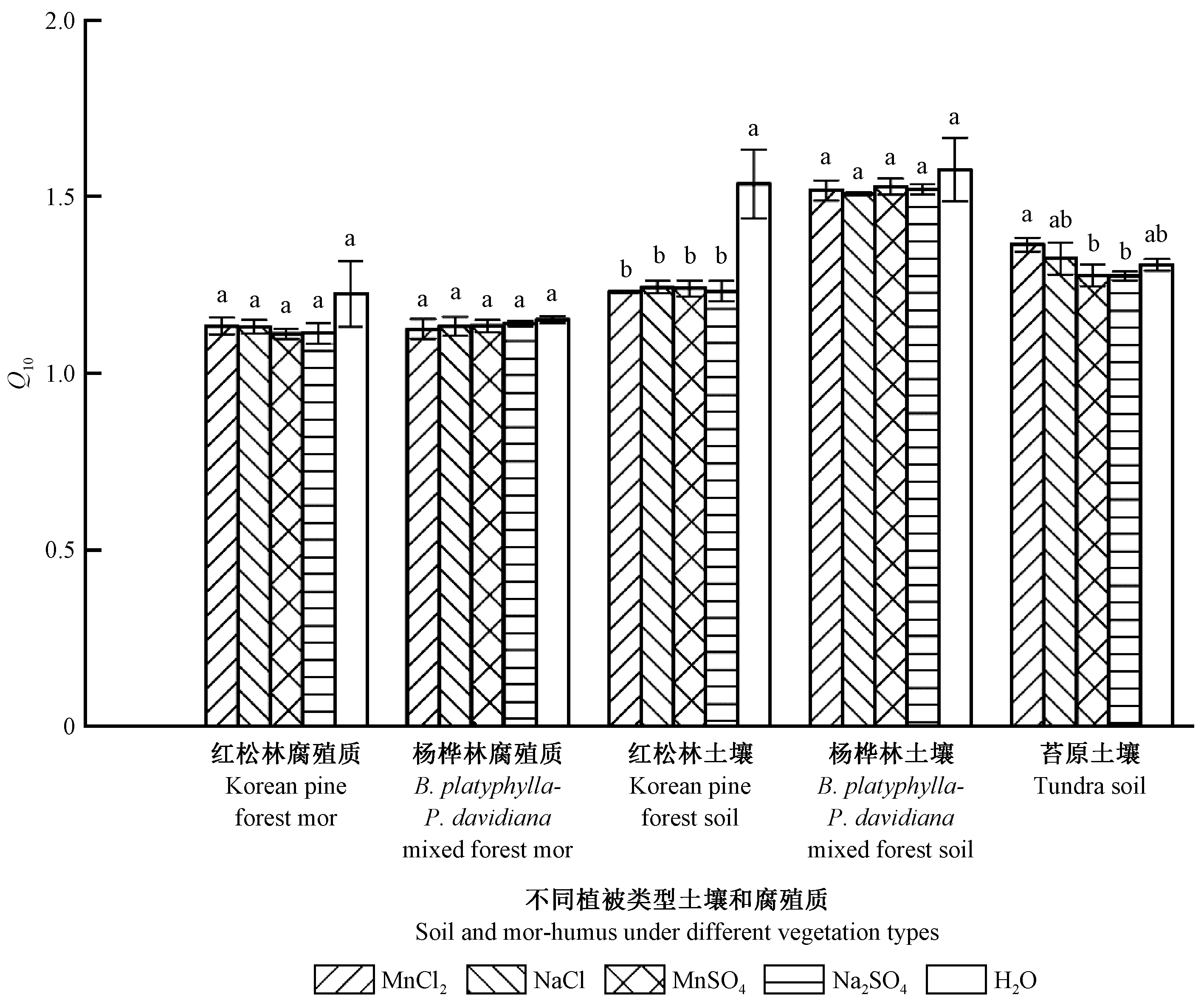
圖3 長白山森林土壤和腐殖質有機碳礦化速率的溫度敏感性(Q10)
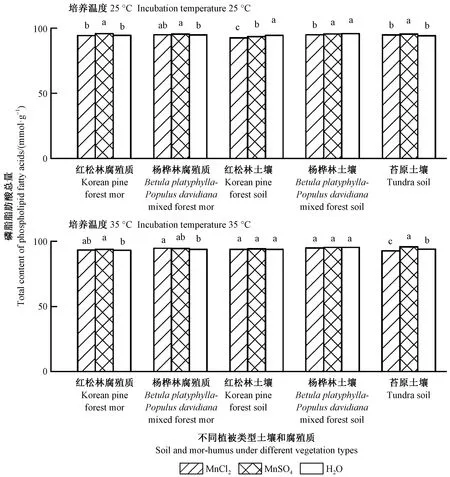
圖4 培養結束時土壤和腐殖質磷脂脂肪酸總量
2.4 土壤和腐殖質磷脂脂肪酸總量
圖4表明: 與對照處理相比,MnSO4添加處理顯著提高闊葉紅松林和楊樺林腐殖質的磷脂脂肪酸總量(P<0.05,培養溫度為35 ℃時楊樺林腐殖質未達顯著性差異); 培養溫度為35 ℃時,MnCl2添加處理的楊樺林腐殖質磷脂脂肪酸總量顯著高于對照處理(P<0.05); 培養溫度為25 ℃時,MnSO4添加處理紅松林土壤磷脂脂肪酸總量顯著低于對照處理(P<0.05),而MnSO4添加處理高山苔原土壤磷脂脂肪酸總量在25和35 ℃培養溫度下均顯著高于對照處理(P<0.05)。
3 討論
3.1 硫添加對土壤和腐殖質頑固性有機碳礦化的影響
本研究發現硫添加提高了長白山森林土壤頑固性有機碳礦化速率,說明硫是長白山森林土壤頑固性有機碳礦化的限制因素。陳晶等(2016)在落葉松(Larixprincipis-rupprechtii)林的研究也發現,土壤呼吸速率與硫沉降水平正相關(P<0.05)。然而,硫輸入易引起土壤酸化,從而影響土壤微生物群落的豐度和活性(Vileetal., 2003)。有學者對亞熱帶森林土壤的研究發現硫添加抑制土壤有機碳礦化(吳建平等, 2015; Wuetal., 2016),這可能是由于南方土壤呈酸性,持續硫添加加劇土壤酸化,從而抑制土壤微生物活性及其調控的有機碳礦化過程。本研究的供試土壤采自溫帶森林,與我國南方地區相比,該地區酸沉降程度相對較低(程念亮等, 2016)。因此,硫添加在一定程度上為土壤微生物提供了養分,提高了土壤和腐殖質層的微生物生物量(圖4),從而促進土壤有機碳礦化。
此外,之前研究發現硫沉降對土壤有機碳礦化的影響還與土壤pH背景值和硫添加水平有關,本研究并未設置不同的硫添加梯度,因此,在何種硫添加水平下將會抑制土壤有機碳礦化,還需今后進一步研究。由于20世紀70年代開始大氣硫沉降在減少,因而其對土壤有機碳礦化的影響易被忽略。然而,在有些地區硫沉降強度還在增加(程念亮等, 2016; Sorimachietal., 2007),因此,未來在土壤碳礦化模型中應將大氣硫輸入作為一個重要參數,以便提高模型的準確性。
3.2 錳添加對土壤和腐殖質頑固性有機碳礦化的影響
MnP能促進土壤和凋落物中木質素和胡敏酸的降解,錳是合成MnP的重要組分(Hatakka, 1994; Hofrichteretal., 1998; Bergetal., 2007; Virzo De Santoetal., 2009)。本研究假設錳添加能促進土壤和腐殖質頑固性有機碳的礦化作用,為此,首先進行了為期90天的預培養,以消耗土壤和腐殖質中的易分解碳組分(Hoosbeeketal., 2007),預培養結束時土壤和腐殖質的碳礦化速率約為初始碳礦化速率的10%(圖1),說明大部分活性碳組分已消耗完,從而認為錳和硫添加處理時測得的礦化速率是來自頑固性有機碳。與預期不同,本研究并未發現添加錳能顯著改變土壤和腐殖質頑固性有機碳的礦化速率。錳添加對森林土壤和腐殖層有機碳礦化的影響與環境理化條件、森林類型以及腐殖質分解程度有關(Trumetal., 2011)。所添加的Mn2+并不能保證參與合成MnP,錳在環境中的形態與pH值和氧化還原條件有關,在本研究供試土壤pH值大于6的環境下,錳的形態也可能是MnO2和Mn2O3等錳氧化物(Lindsay, 1979),這在一定程度上限制錳參與合成MnP。本研究中錳添加對長白山森林土壤和腐殖質有機碳礦化沒有顯著影響,說明錳不是該地區森林土壤和腐殖質層頑固性有機碳礦化的限制因子。
4 結論
本研究首次探討了硫和錳添加對長白山森林土壤和腐殖質頑固性有機碳礦化速率及其溫度敏感性的影響,發現硫添加顯著提高森林礦質土層和腐殖質層頑固性有機碳礦化速率,這是由于硫添加通過提高土壤微生物的生物量而促進了有機碳分解。錳添加對土壤和腐殖層頑固性有機碳礦化沒有顯著影響。硫和錳添加對Q10的影響因森林類型而異,對闊葉紅松林表現為顯著降低,對楊樺林和高山苔原沒有顯著影響。建立土壤有機碳礦化模型時應將硫輸入作為一個重要參數。要準確預測森林土壤碳礦化對硫輸入水平的響應,還需開展硫添加梯度的野外定位研究。
