外源油菜素內酯對低溫脅迫下水稻幼苗生長及生理特性的影響
2020-09-10 02:41:08周偉江吳旺嬪唐才寶肖志芳陳光輝
西北農業學報 2020年9期
周偉江,吳旺嬪,唐才寶,肖志芳,陳光輝,王 悅
(1.湖南農業大學 農學院,長沙 410128;2.水稻油菜抗病育種湖南省重點實驗室,長沙 410128)
低溫是影響作物生長發育及產量最主要的逆境之一[1]。水稻作為中國主要的糧食作物之一,在早春常遇到“倒春寒”天氣,導致水稻秧苗遭受低溫冷害,從而造成巨大的經濟損失。
油菜素甾醇(Brassinosteroids,BRs)是一類廣泛存在于植物體內的生理活性極強的植物激素[2]。大量研究表明,2,4-表油菜素內酯(2,4-Epibrassinolide,EBR)可以顯著增強植物的抗逆性。李杰等[3]研究發現,噴施EBR能夠緩解低溫脅迫對辣椒幼苗生長的抑制作用,表現為地上部干物質的增加,植株抗低溫能力增強。Hu等[4]研究表明,EBR預先處理黃瓜,能夠提高植株光合速率和光化學效率,進而增強黃瓜幼苗低溫耐受能力。李淑葉等[5]證明,噴施EBR能夠減輕低溫脅迫對棉花幼苗細胞膜的損害,通過提高葉綠素含量、提升PSⅡ反應中心活性和增強PSⅡ受體側電子傳遞能力,進而提高低溫脅迫下棉花幼苗的光合能力。丁丹陽等[6]發現,干旱脅迫下對煙草幼苗噴施EBR不僅能顯著促進幼苗的生長和根系發育,通過提高葉綠素含量增強其光合速率,還能降低O2-產生速率,從而減少對幼苗的損傷,增強其耐旱能力。
目前關于油菜素內酯的研究大多集中于逆境脅迫下植物的光合作用效率[7]、生理特性[8]、種子萌發[9]等方面的影響,而對低溫脅迫下油菜素內酯對水稻幼苗生長及生理特性的研究卻鮮有報道。因此,本研究以‘湘早秈45號’為材料,通過不同濃度EBR噴施處理,測定低溫脅迫下幼苗生長形態指標、丙二醛含量和抗氧化酶活性等,為水稻幼苗低溫耐受機制研究奠定基礎。
1 材料與方法
1.1 試驗材料
供試水稻(OryzasativaL)品種為‘湘早秈45號’,由湖南省水稻研究所提供;EBR購買于美國Sigma 公司。
1.2 試驗方法
EBR先用乙醇溶解,再用蒸餾水定容,配成6個濃度,分別為0、0.001、0.01、0.1、1.0、10.0 μmol/L。挑選粒型整齊一致的水稻種子,經φ=40%次氯酸鈉溶液消毒40 min后,再用滅菌蒸餾水浸種24 h,將浸種后的水稻種子清洗 3~5次,置于37 ℃進行催芽,催芽24 h后播種于留孔的PCR板中,置于盛有蒸餾水的定制培養盒進行培養。用蒸餾水進行緩苗,3 d后用1/2營養液進行培養,待幼苗生長至一葉一心期時,再用全營養液進行培養,營養液參照國際水稻研究所公布的水稻營養液配方,每3 d換一次營養液,培養環境溫度(25±0.5) ℃,濕度(75±5)%。試驗共設置7個處理:CK1,常溫下噴施蒸餾水;CK2,低溫下噴施蒸餾水;T1,低溫下噴施0.001 μmol/L EBR;T2,低溫下噴施0.01 μmol/L EBR ;T3,低溫下噴施0.1 μmol/L EBR ;T4,低溫下噴施1.0 μmol/L EBR ;T5,低溫下噴施10.0 μmol/L EBR。待幼苗生長至兩葉一心期時,于20:00用不同濃度EBR進行葉面噴施,為了增強葉片對EBR的吸收效果,在噴施溶液中加入φ=0.1%的吐溫20,每個PCR板噴施5 mL處理液,連續噴施3 d后放入RLD-1000E-4人工氣候箱(寧波樂電儀器制造有限公司)進行處理,每個處理6盆幼苗,每盆4個PCR板,每板48株幼苗,重復4次。常溫對照溫度為25 ℃/25 ℃(晝/夜),低溫處理溫度為4 ℃/4 ℃(晝/夜),白天光照度為 4 000 lx。光照時數12 h,相對濕度80%。于低溫處理前(0 d)、低溫處理后 3 d取樣并測定相關指標。
1.3 測定指標及方法
1.3.1 生長量 選取長勢一致的幼苗20株,用電子游標卡尺測定距根基部1 cm處莖粗(水稻幼苗莖稈為橢圓形,測定長軸長度),用直尺測量根基部到生長點的株高。蒸餾水將根系沖洗干凈后用吸水紙吸干,用萬分之一電子天平(Mettler Toledo ME 104E,瑞士)測定植株地上部分與地下部分鮮質量。將新鮮樣品進行105 ℃殺青30 min后,60 ℃烘至恒量,用萬分之一天平測定干質量。
壯苗指數=(莖粗/株高 + 根干質量/地上部干質量)×全株干質量。
1.3.2 根系形態分析 選取長勢一致的植株20株,用HT2A-3獨立版型掃描儀(上海中晶科技有限公司,中國)掃描幼苗根系圖片并保存,用Microtek ScanWizard EZ型根系形態分析系統(上海中晶科技有限公司,中國)進行分析,得出根系長度、表面積、體積、平均直徑、連接數、根尖數。
1.3.3 抗氧化系統相關指標 超氧化物歧化酶(superoxide dismutase, SOD)活性測定采用氮藍四唑光化還原法[10];過氧化物酶(peroxidase, POD)活性采用Kochba等[11]的方法測定;過氧化氫酶(catalase, CAT)活性采用Aebi[12]的方法測定;丙二醛(malondialdehyde, MDA)含量采用Hodges等[13]的硫代巴比妥酸法測定;可溶性蛋白(soluble protein, SP)含量采用考馬斯亮藍G-250法[14]測定。
1.3.4 數據分析 使用Microsoft Excel 2007和SPSS 17.0軟件進行數據分析和作圖,用Duncan’s新復極差法進行差異顯著性檢測(α= 0.05)。
2 結果與分析
2.1 EBR對低溫脅迫下水稻幼苗形態的影響
由表1可知,低溫處理前(0 d),T4處理相比對照(CK1、CK2),其地上部分鮮質量、地上部分干質量、地下部分鮮質量、地下部分干質量以及壯苗指數均顯著提高,分別增加13.55%、 23.68%、18.35%、20.99%和28.57%。低溫處理3 d后,水稻幼苗生長會受到抑制,而T4處理的水稻幼苗株高、莖粗、地上部分鮮質量、地上部分干質量、地下部分鮮質量、地下部分干質量以及壯苗指數較CK2分別提高9.65%、9.22%、 17.04%、26.76%、26.32%、29.41%和40.74%,均達到顯著性差異。
2.2 EBR對低溫脅迫下水稻幼苗根系形態的 影響
由表2可知,低溫處理前(0 d),適宜濃度EBR能促進低溫脅迫下水稻幼苗根系的生長,以T2和T4處理效果較好,而T5處理會抑制水稻幼苗根系的生長。低溫處理3 d后,T4處理表現最好,其水稻幼苗根系的長度、表面積、體積、連接數和根尖數均顯著提高,較CK2分別增加 34.29%、25.19%、14.42%、32.02%和42.8%。
2.3 EBR對低溫脅迫下水稻幼苗抗氧化酶活性的影響
由圖1可知,低溫處理前(0 d),水稻幼苗葉片與根系的抗氧化酶活性隨EBR濃度的增大呈先增加后下降的趨勢,并以T4處理效果較好,其幼苗葉片的SOD、POD和CAT活性相比CK2分別顯著增加了47.71%、19.61%和21.44%;幼苗根系較CK2分別提高了38.37%、27%和50%,均達到顯著性差異。低溫處理3 d后,水稻幼苗根系SOD、POD、CAT活性增強,以T4處理表現最好,較CK2顯著提高 47.15%、14.88%和73.92%。

表1 EBR處理及低溫脅迫下水稻幼苗形態特征Table 1 Morphological characteristics of rice seedlings under EBR treatment and chilling stress
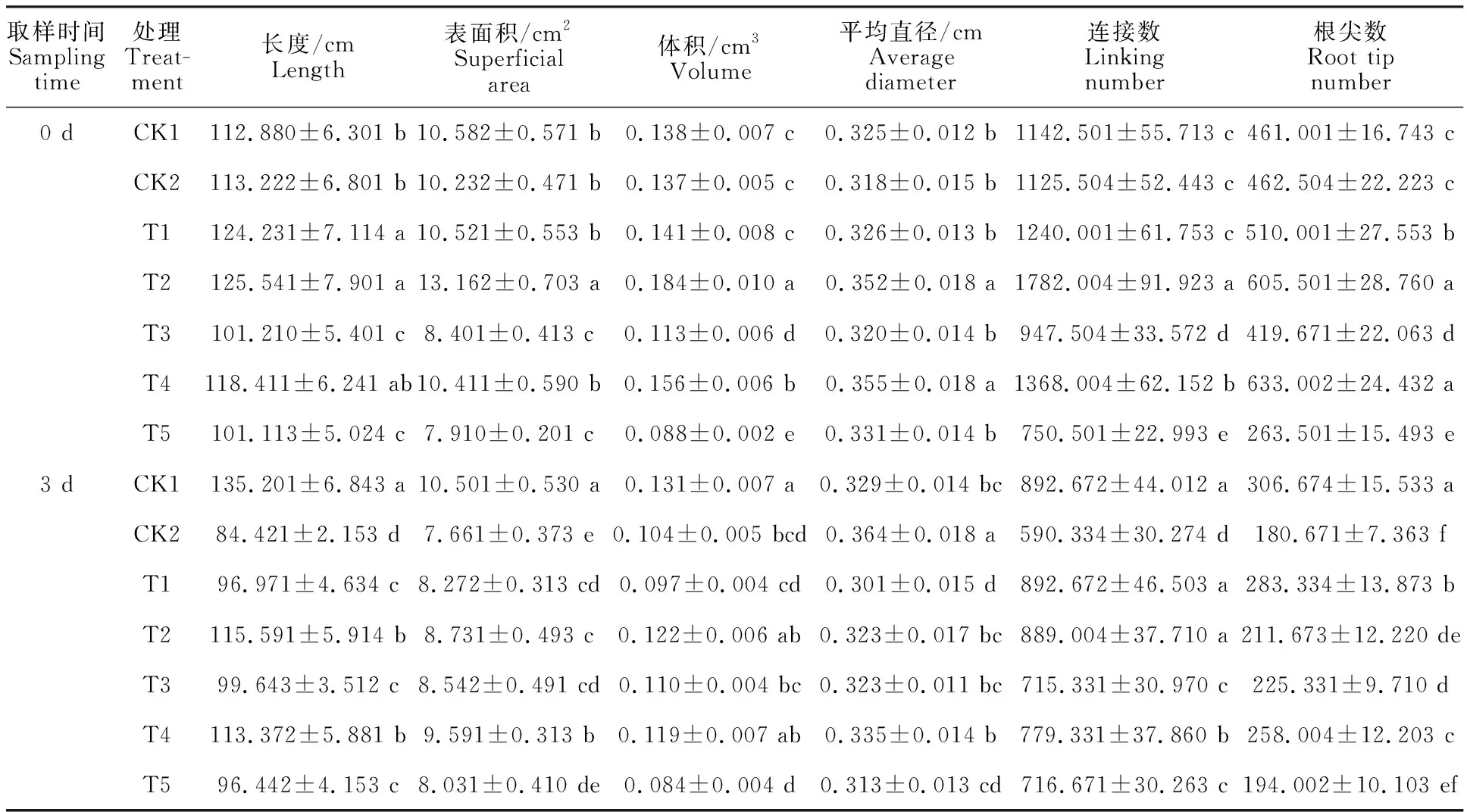
表2 EBR處理及低溫脅迫下水稻幼苗根系形態特征Table 2 Root morphological characteristics of rice seedlings under EBR treatment and chilling stress
2.4 EBR對低溫脅迫下水稻幼苗MDA及SP含量的影響
由圖2可知,低溫處理前(0 d),噴施EBR能顯著降低水稻幼苗葉片和根系的MDA含量,并增加其SP含量。其中,T4處理的水稻幼苗葉片中MDA含量相比CK2顯著降低了25.09%,而SP含量則顯著增加67.73%;根系中MDA含量較CK2顯著降低20.21%,而SP含量則顯著增加163.82%。低溫處理3 d后,T4處理效果最為明顯,其水稻幼苗葉片和根系的MDA含量相比CK2分別顯著降低30.19%和34.03%,而SP含量則分別顯著增加41.71%和56.48%。
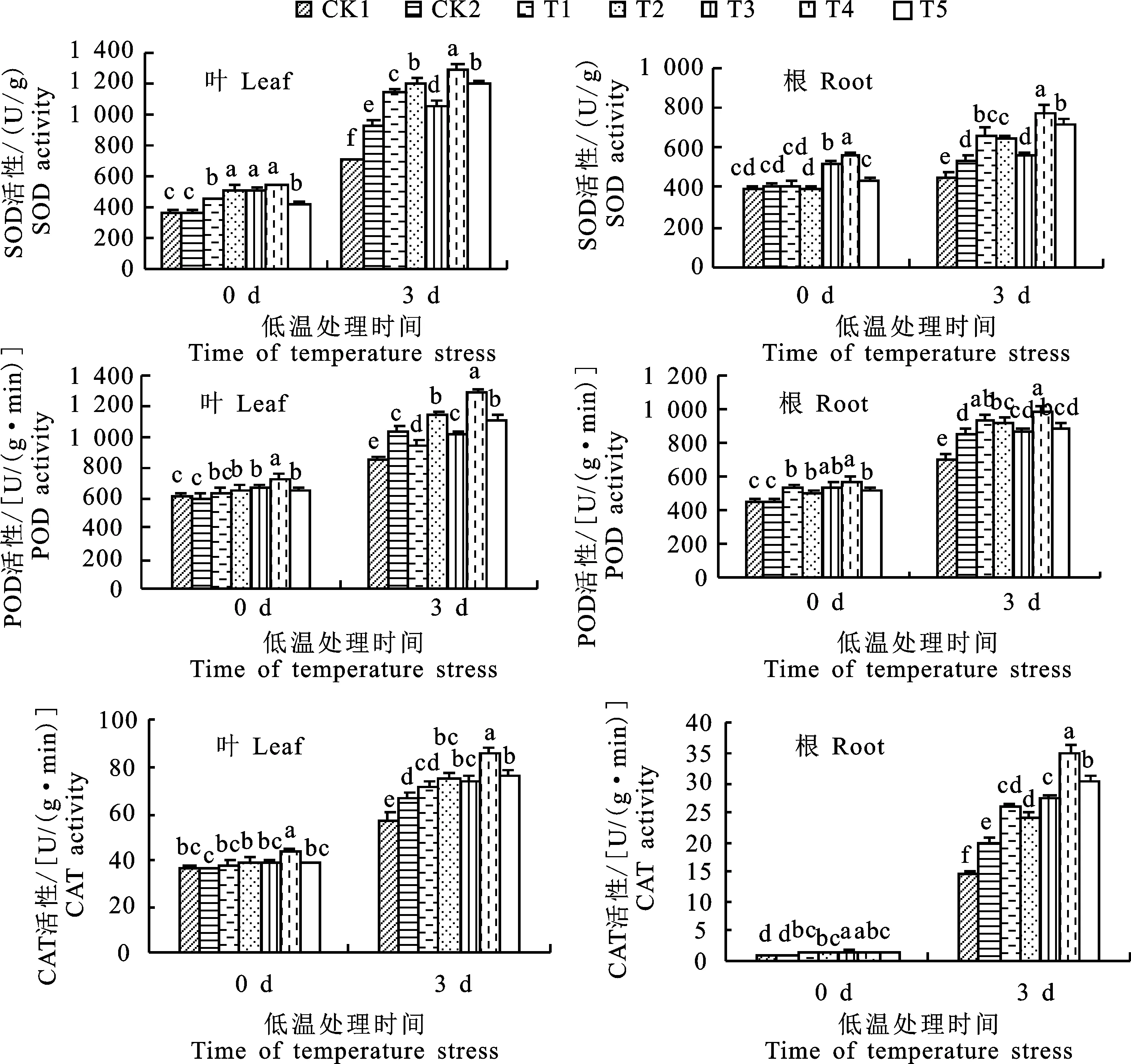
圖中不同小寫字母表示同一時期同一指標處理間差異顯著(P<0.05)。下同

圖2 EBR處理及低溫脅迫下水稻幼苗MDA及SP含量Fig.2 MDA and SP content of rice seedlings under EBR treatment and chilling stress
3 討 論
低溫脅迫主要通過減小幼苗葉片的柵欄組織厚度,造成植株生物量減少,從而影響植株形態建成、物質合成與積累[15]。有研究發現,10 ng/L的BR處理幼苗第二節間,便可以引起該節間顯著伸長,促進細胞分裂,造成節間膨大,甚至開裂[16]。王道平等[17]研究表明,低溫脅迫使水稻幼苗單株鮮質量顯著減少,而EBR對單株鮮質量無明顯影響。而本試驗發現,在低溫脅迫下水稻幼苗生長受到顯著的抑制作用,但噴施1.0 μmol/L EBR具有明顯的緩解作用,表現在幼苗的株高和莖粗均得到顯著提升,幼苗地上部和地下部干鮮質量、壯苗指數和根冠比均較低溫對照有顯著的提高。這與徐曉昀等[7]、閆慧萍等[9]的研究結果一致,這可能與EBR能夠通過誘導細胞分裂和伸長,改善滲透調節能力,從而增強植株抗逆能力有關。
根系是植物吸收水分與礦質養分的主要器官,其形態及構型在極大程度上決定著植株獲取養分的能力[18]。康云艷等[19]研究發現,低氧脅迫下黃瓜幼苗根系生長遭受了明顯的抑制作用,噴施EBR則能有效緩解這種抑制作用。趙雪松等[20]研究證明,1.0 μmol/L EBR處理6 h后,植株體內結合態生長素轉化為游離態生長素的過程受到顯著抑制,致使根尖頂端優勢的喪失,從而促進側根的生長發育。閆小紅等[21]研究表明,在低溫15 ℃下,無論是浸泡法還是添加法,不同濃度EBR對辣椒幼苗根長均表現為抑制作用。本試驗研究表明,1.0 μmol/L EBR處理能極大地促進水稻幼苗根系生長,使根系長度、表面積、體積、平均直徑、連接數及根尖數顯著增加,而在低溫脅迫下水稻幼苗根系生長受抑制,但噴施EBR能極大地緩解這種抑制作用。這可能與EBR能夠在低溫脅迫下,通過誘導轉錄因子BZR1,激活參與細胞壁形成的相關基因,促進根系細胞伸長有關。大量研究也表明,EBR能夠通過與生長素互作,調控快速堿化因子,調節側根發育,促進細胞的縱向生長[22-23]。

綜上所述,低溫脅迫下,適宜濃度的EBR能通過提高水稻幼苗株高、莖粗、地上與地下部分干鮮質量、壯苗指數及根冠比,增加根系長度、表面積、體積、平均直徑、連接數以及根尖數,增強葉片與根系中抗氧化酶(SOD、POD、CAT) 活性,降低MDA含量,加快SP含量積累來緩解低溫對水稻幼苗的傷害。因此,低溫脅迫下噴施1.0 μmol/L EBR溶液能明顯減輕低溫脅迫對水稻幼苗生長的抑制作用,增強其低溫耐受能力。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
軍事文摘(2021年22期)2021-11-26 00:43:51
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
文苑(2018年22期)2018-11-19 02:54:14
