甘藍型油菜HD-ZIP基因家族的鑒定和表達分析
2020-09-10 02:41:02昝領兄李開祥賈永鵬杜德志
西北農業學報 2020年9期
關鍵詞:分析
昝領兄,李開祥,賈永鵬,杜德志
(青海大學 農林科學院,青海省春油菜遺傳改良重點實驗室,國家油菜改良中心青海分中心,農業農村部春油菜科學觀測實驗站,青海省春油菜工程技術研究中心,西寧 810016)
油菜是主要的油料作物之一,由于各地環境條件有所差異,對油菜品種的特性要求越來越高,在產量高的基礎上,還應具有優質、抗倒伏和抗逆性等其他特點。其中甘藍型油菜作為主要的油菜栽培品種,因具有適應性廣,產量高,品質優良等許多特性,使其種植面積最為廣泛。因此揭示甘藍型油菜抗逆性分子機理的研究對農業生產至關重要。
同源異型亮氨酸拉鏈蛋白(homeodomain-leucine ZIPper, HD-ZIP)是高等植物所特有的一類轉錄因子,其中包括C末端由61個氨基酸構成的高度保守的DNA結合結構域(Homeodomain, HD)和跟隨其后的亮氨酸拉鏈結構域(Leucine ZIPper, LZ)[1-2]。通過LZ結構域,HD-ZIP蛋白才能折疊成為同源或異源二聚體,并且LZ結構域可以實現和其他蛋白質的相互作用,使該蛋白發揮不同的功能。此外,有研究表明含有HD-ZIP結構域的轉基因植物在受到不同脅迫的環境條件下能夠表現出優良的的形態特征[3-4]。根據該基因家族成員的序列相似性和結構域的不同可將HD-ZIP基因家族成員分為四大類,即HD-ZIP Ⅰ,Ⅱ,Ⅲ和Ⅳ[5-6]。這四大類都同時包含保守的HD和LZ結構域,促進了HD-ZIP蛋白的二聚化,但是這四大亞族之間在結構上也存在差異,即HD-ZIP Ⅰ和HD-ZIP Ⅱ可以識別DNA序列CAAT(A/T)ATTG,而HD-ZIP Ⅲ和HD-ZIP Ⅳ可識別的DNA序列為GTAAT(G/C)ATTAC和TAAATG(C/T)A[7-8]。此外,HD-ZIP Ⅱ還另外包含2個五組保守的氨基酸,半胱氨酸(Cys) 、脯氨酸(Pro) 、絲氨酸(Ser) 、半胱氨酸(Cy)和谷氨酸(Glu) ,被稱為CPSCG域。 HD-ZIP Ⅲ和HD-ZIP Ⅳ共享一個高度結合的脂質/甾醇結合區域,稱為START域[9-10]。
由于這四類亞家族成員在結構上存在差異,使得它們在各自的功能方面也有所不同。HD-ZIP Ⅰ蛋白質在植物對于多種環境條件的適應方面發揮重要作用,比如干旱,脫落酸(ABA)脅迫,鹽脅迫和極端溫度等。HB1屬于HD-ZIP Ⅰ家族,在蒺藜苜蓿的初生根和側根分生組織中被表達,研究發現它可參與對鹽和滲透脅迫的反應[11]。在玉米HD-ZIP Ⅰ家族的Zmhdz10基因通過ABA依賴的信號傳導途徑調節植物的干旱和耐鹽性脅迫[12]。HD-ZIP Ⅱ亞家族成員基因主要在避光或光照信號,植物發育和非生物脅迫方面發揮著重要功能[13]。例如,其家族的ATHB2、HAT2、HAT3和HAT4的表達主要受光敏色素調節,因為近紅外光與遠紅光之間的比例會迅速誘導避光響應[14-15]。據報道,HD-ZIP Ⅲ可以作為一種調節因子參與植物細胞的橫向伸長,木質部分化,生長素運輸和側器官分化[16-17]。HD-ZIP Ⅳ對植物表皮發育過程起著至關重要的作用[18];有研究發現這類蛋白質被認為參與毛狀體形成,脂質代謝和角質層生物合成[19]。玉米OCL4(外細胞層4)抑制玉米形成毛狀體并影響花藥分裂或分化過程[20]。
綜上所述,HD-ZIP蛋白在植物特異的發育方面表現出重要的作用,因此了解該家族成員的序列特征及相關生物信息學分析對于進一步挖掘并利用它們在各種植物中的功能機制必不可少。目前,HD-ZIP蛋白家族的基因功能在許多植物中都已經進行了分析和研究。如擬南芥[21]、玉米[22]、水稻[23]、楊樹[24]、黃瓜[25]、大豆[26]等。但在油菜中未見相關報道。因此,本研究利用已公布的甘藍型油菜基因組信息,通過對HD-ZIP家族基因的進行鑒定和序列分析,可為甘藍型油菜的抗性研究及新品種培育奠定基礎。
1 材料與方法
1.1 HD-ZIP基因的序列檢索與鑒定
根據對HD-ZIP蛋白的相關研究,通過Pfam32.0(http://pfam.xfam.org/)數據庫分別下載同源框的保守HD結構域和亮氨酸拉鏈LZ結構域的隱馬爾可夫模型(HMM)PF00046和PF02183[27]。并使用Hmmsearch搜索工具在甘藍型油菜基因組(http://www.genoscope.cns.fr/brassicanapus/)中尋找具有同源序列的基因[28],以在線軟件NCBI-CDD(http://www.ncbi.nlm.nih.gov/Structure)對獲得的蛋白序列進行結構域鑒定,選擇同時含有HD和LZ結構域的蛋白作為甘藍型油菜HD-ZIP蛋白基因家族。從甘藍型油菜基因序列數據庫中檢索這些基因編碼序列的注釋信息,最后使用在線工具ExPASY(http://web.expasy.org/protparam)對獲得序列進行氨基酸數目(aa),等電點(PI)和分子質量(u)等物理參數的統計分析。
1.2 HD-ZIP家族基因染色體定位和基因結構分析
在甘藍型油菜基因組數據庫中搜索HD-ZIP家族基因在染色體上的具體位置和每條染色體總長度,利用在線工具MapGene2Chromosome v2(http://mg2c.iask.in/mg2c_v2.0)繪制該家族基因的染色體定位圖[29]。結合基因家族的gtf3文件,使用在線分析軟件GSDS(http://gsds.cbi.pku.edu.cn/)繪制基因結構圖,分析基因的內含子和外顯子分布情況。
1.3 HD-ZIP家族基因進化樹分析和motif 預測
利用Clustal[30]工具進行氨基酸序列比對。通過MEGA7.0[31]軟件,以鄰接法(NJ,Neighbor-Joining)構建系統發育進化樹,使用poisson模式,將Bootstrap參數設置為1 000,其余參數設置為默認。并利用在線軟件MEME[32](http://meme-suite.org/tools/meme)鑒定該家族基因蛋白保守基序,除了基序數量設置為20個,其余參數設置為默認。Motif分析結果使用TBtools軟件下載保存[33]。
1.4 HD-ZIP家族基因之間的共線性分析
利用軟件McScanX[34]對甘藍型油菜的HD-ZIP家族基因進行共線性分析,采用Circos繪制關系圖。
1.5 甘藍型油菜HD-ZIP家族基因的表達分析
為了進一步分析和挖掘HD-ZIP轉錄因子在甘藍型油菜不同組織中的表達模式,從NCBI數據庫中下載已發表的轉錄組數據(在鹽脅迫處理下,甘藍型油菜HD-ZIP家族基因在根和葉片中的表達情況)。通過fasterq-dump軟件將SRA數據轉化為fastq數據,同時利用fastqc進行數據質控和過濾處理,然后利用STAR軟件和甘藍型油菜參考基因組進行比對,接著使用RSEM軟件進行表達量定量分析,最后使用R軟件中的pheatmap軟件包繪制表達熱圖。
2 結果與分析
2.1 甘藍型 HD-ZIP基因的鑒定及其染色體 定位
為了在甘藍型油菜全基因組中鑒定所有的HD-ZIP基因家族成員和其在染色體上的分布情況,通過Pfam數據庫中的PF00046 和PF02183在甘藍型油菜基因組數據庫(E-value=le-5)中對HD-ZIP家族基因進行初步的篩選,并且使用在線軟件NCBI-CDD和ExPASY進行結構域鑒定,保留同時含有HD和LZ結構域的蛋白,最終在甘藍型油菜基因組上獲得了74個HD-ZIP基因成員(表1)。結果顯示:74個HD-ZIP 轉錄因子的大小為162~691 aa,其中編碼區序列和對應的氨基酸長度最短的為489 bp和162aa(BnaA08g04720D),最長的為2 076 bp和691aa(BnaC05g19660D),平均氨基酸個數為279個。蛋白分子質量(MW)為18 275.57 u(BnaA 08g04720D)~76 164.87 u(BnaC05g19660D),等電點(PI)為4.54(BnaA09g42630D)~10.06(BnaA 09g18250D)。
染色體定位結果顯示:這74個基因在甘藍型油菜19條染色體中勻有分布,此外還包括8條隨機序列(圖1),其中A染色體組上共有30個,除了A01上僅有1個,其余9條染色體上有2~4個不等,C染色體上共有25個,每條染色體有 2~4個,其余19個分布在8條隨機染色體上。綜上所述基因在染色體組上的分布不均勻。
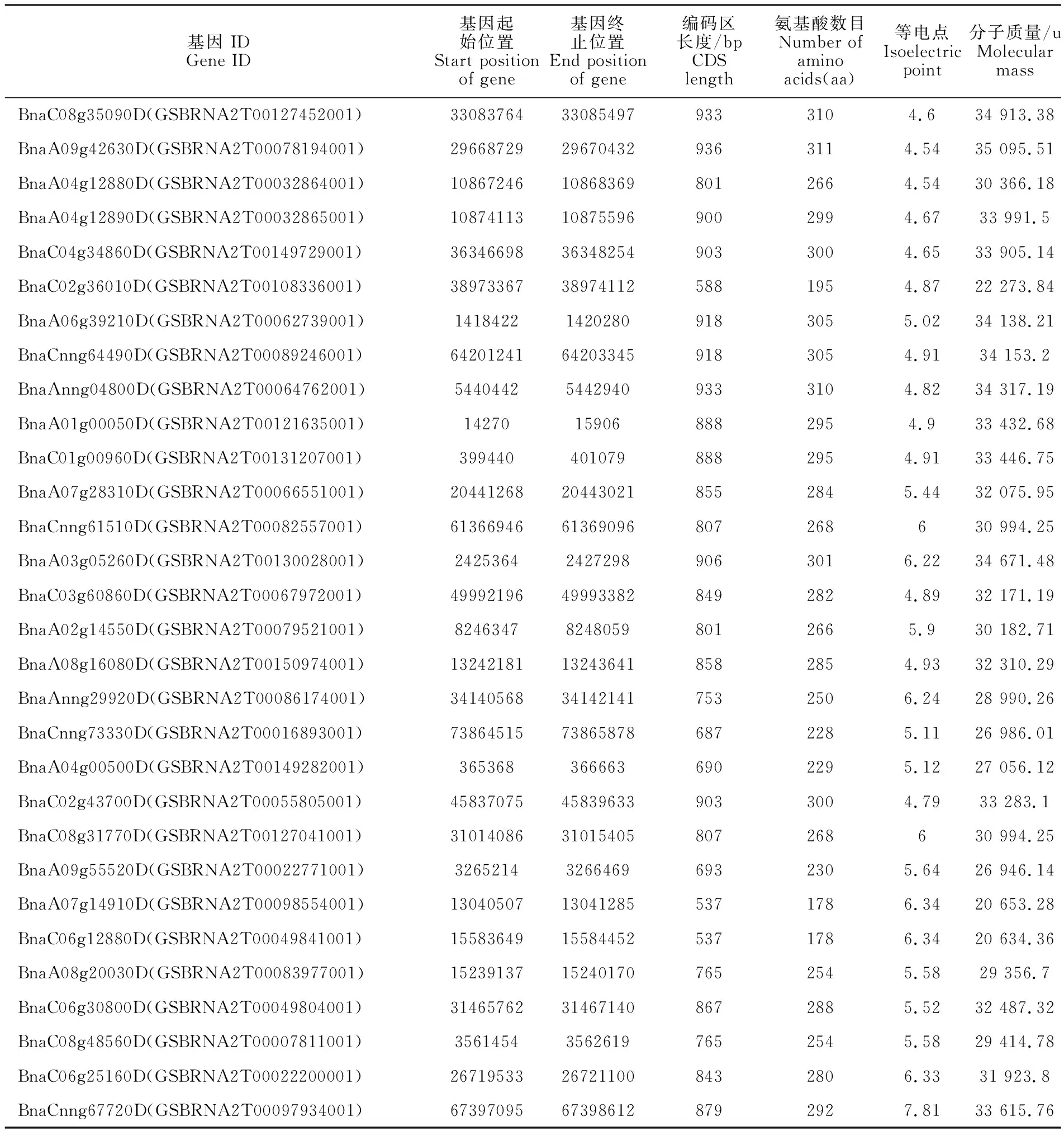
表1 甘藍型油菜HD-ZIP基因家族的特征Table 1 Characteristics of HD-ZIP gene family in Brassica napus
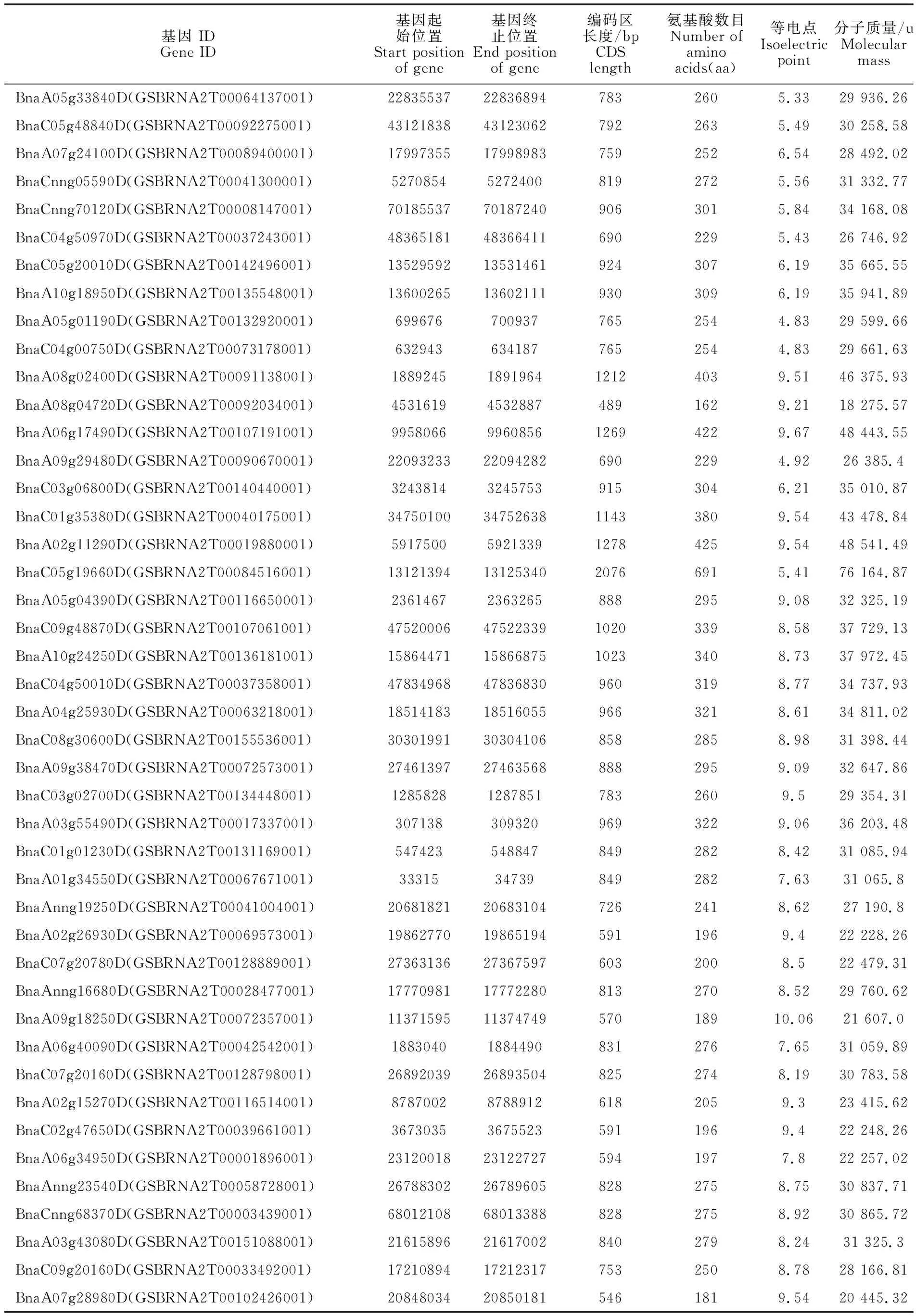
(續表1 Continued table 1)
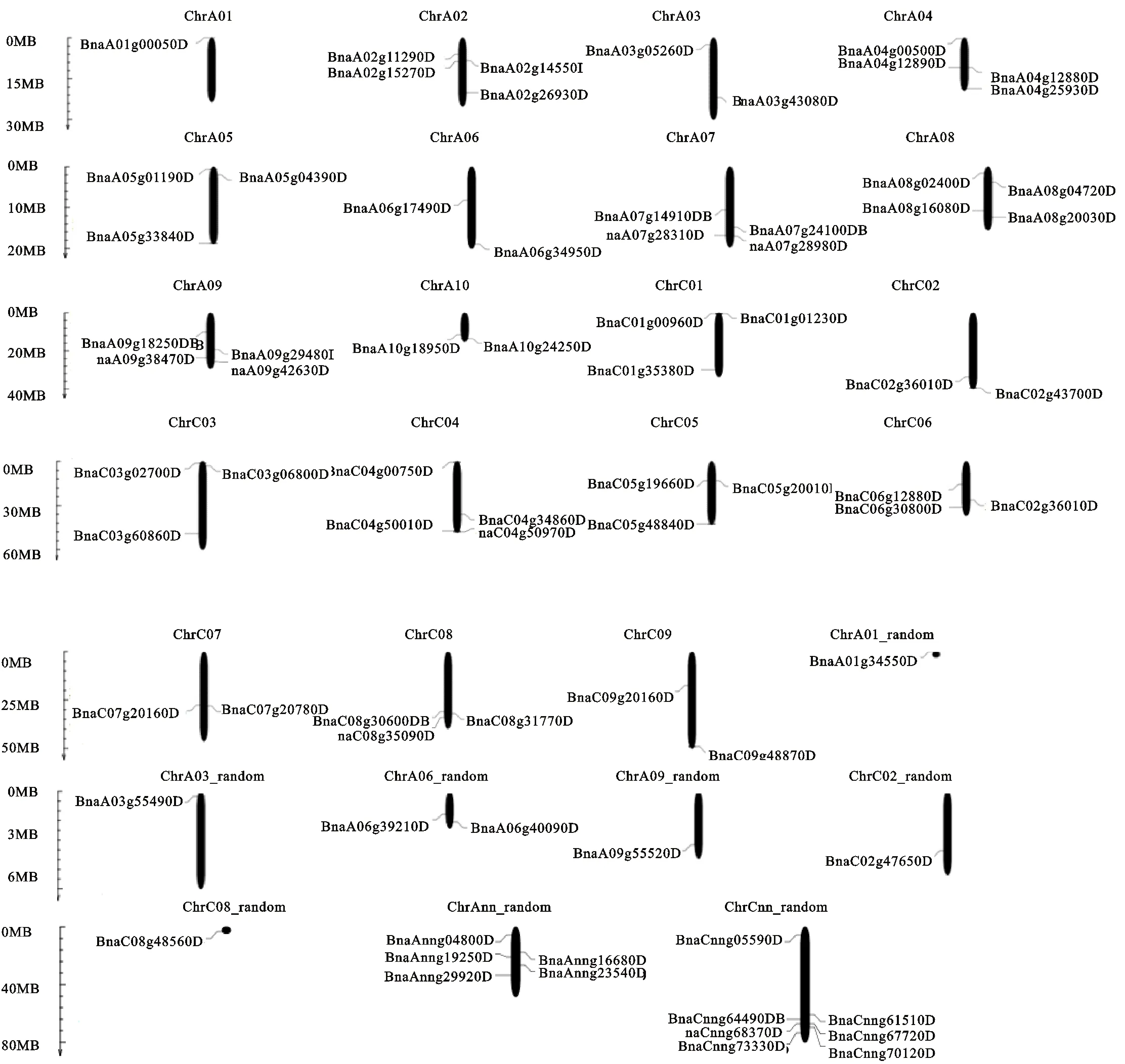
圖1 HD-ZIP家族基因的染色體定位Fig.1 Mapping of HD-ZIP family gens in B.napus chromosme
2.2 甘藍型油菜HD-ZIP家族基因系統進化樹分析
為明確甘藍型油菜中 HD-ZIP 轉錄因子的分類情況,本研究利用MAGA 7軟件對74個甘藍型油菜 HD-ZIP 轉錄因子進行系統發育進化樹的構建(圖2)。結果顯示:74個HD-ZIP轉錄因子可被分為4大亞族,分別為HD-ZIP Ⅰ、HD-ZIP Ⅱ、HD-ZIP Ⅲ 和 HD-ZIP Ⅳ,其中有39個HD-ZIP轉錄因子被聚類在HD-ZIP Ⅰ,是最多的一類,占HD-ZIP轉錄因子總家族的52.7%;HD-ZIP Ⅱ和HD-ZIP Ⅳ分別占HD-ZIP轉錄因子總家族的9.5%和35.1%;HD-ZIP Ⅲ所包含的HD-ZIP轉錄因子最少,僅有2個:分別是BnaC05g19660D和BnaA09g29480D。從進化樹可看出HD-ZIP Ⅰ和HD-ZIP Ⅱ處在同一分枝,其親緣關系可能也比較相近。
2.3 甘藍型油菜HD-ZIP家族基因結構和motif預測
使用在線軟GSDS繪制HD-ZIP基因家族的基因結構圖(圖3),分析內含子和外顯子分布情況。HD-ZIP家族基因的DNA序列和CDS分析表明:74個甘藍型油菜HD-ZIP 的內含子個數從 1到10不等,在相同的亞族中,內含子數量差異不大,具有相似的結構。其中HD-ZIP Ⅱ的內含子數量集中為1~2,HD-ZIP Ⅳ的基因內含子個數為2~4個,基因結構比較簡單;HD-ZIP Ⅰ和HD-ZIP Ⅲ的內含子個數從1~10不等;HD-ZIP家族基因的內含子在不同亞族間基因結構存在較大差異,可能是因為HD-ZIP Ⅰ和HD-ZIP Ⅲ在進化過程中內含子大量插入導致的。

圖2 甘藍型油菜HD-ZIP轉錄因子的系統進化發生樹Fig.2 Unrooted phylogenetic tree of HD-ZIP transcription factors in B.napus
通過MEME在線工具對甘藍型油菜HD-ZIP轉錄因子的保守基序進行分析(圖4)。結果顯示:HD-ZIP家族的74個蛋白都含有motif 1和motif 2,這2個結構共同組成了較為保守的HD-ZIP結構域。在基因結構圖和保守基序預測圖中可發現同源基因間盡管在結構上存在較大的差異,但預測到的保守基序數目基本一致,比如BnaC05g19660D含有10個內含子,BnaA09g 29480D只含有3個內含子,但這2個同源基因預測到的 motif數目基本一致。但也有相反的情況,如BnaC06g12880D預測到了3個motif ,BnaC02g43700D中存在6個motif,但這2個基因都只含有2個內含子。所以同一亞族中的同源基因間基本具有相同的基序,暗示其具有的功能也可能類似。
2.4 甘藍型油菜HD-ZIP家族共線性分析
對甘藍型油菜的74個HD-ZIP家族基因進行串聯重復性分析,挖掘HD-ZIP家族基因在甘藍型油菜A和C基因組中的重復情況。分析結果如圖5所示:鑒定到的74個HD-ZIP基因中大部分基因在甘藍型油菜的A染色體組和C染色體組中存在重復,具有共線性關系,其中有34個基因在A和C染色體組之間具有共線性關系。同時也發現,有11個基因在A染色體組內存在共線性,5個基因在C染色體組內存在共線性。進一步說明,在異源四倍體的甘藍型油菜基因組中,這些基因不僅在A和C染色體組組內存在同源關系,而且染色體組之間存在較多的同源性,并且存在共線性關系的基因在系統進化樹中也都被聚為一類,這也表明在基因組進化過程中HD-ZIP蛋白家族基因在甘藍型油菜中相對保守。

圖3 甘藍型油菜HD-ZIP家族基因結構Fig.3 Gene structure of HD-ZIP family in B.napus
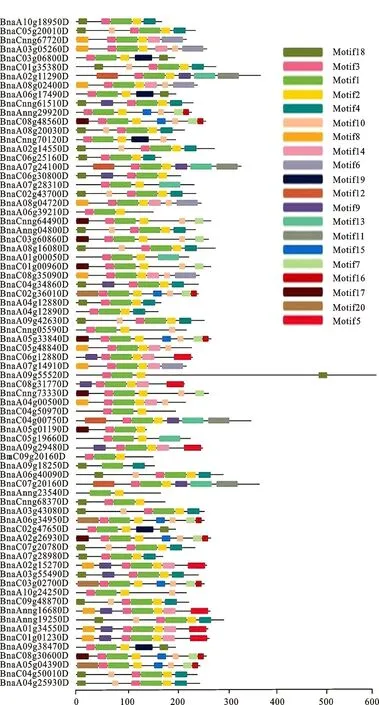
圖4 甘藍型油菜HD-ZIP轉錄因子進化樹和保守基序Fig.4 Phylogetic tree and gene motifs of HD-ZIP family transcription in B.napus
2.5 甘藍型油菜HD-ZIP家族基因的表達分析
為進一步了解HD-ZIP家族基因在脅迫處理下的表達變化。結合甘藍型油菜在鹽脅迫處理下的根和葉片轉錄組數據。結果分析如圖6所示,從圖中可知,HD-ZIP家族的大部分基因在根和葉片中都發生不同程度的表達,BnaC08g35090D和BnaA09g42630D在葉片中的表達量最高,BnaA01g0050D和BnaA05g33840D在根中有最高的表達量。根中表達的基因數目多于葉片中,并且BnaA03g052600D、BnaC03g06800D等13個基因在根中的表達量要明顯高于葉片,受到鹽脅迫后,根和葉片中的大部分基因表達量會增加,并隨著脅迫處理的時間的增加,基因的表達量又會受到一定的下調,如BnaCnng73330D和BnaA04g00500D等基因在1 h和12 h對照下表達量很低,但在鹽脅迫1 h和12 h后表達量增加,并且脅迫1 h時表達量很高,但在12 h時表達量又相對較低,說明這些基因可以響應鹽脅迫。只有少數基因的表達量未發生變化或者下調,如BnaC09g20160D、BnaA05g17490D、BnaA02g11290D、BnaA05g33840D。根和土壤直接接觸,受脅迫后,根中的基因不斷的調節變化應對外界的脅迫,然后再將脅迫的信息傳遞到葉片中,進行相互協調。

圖5 甘藍型油菜HD-ZIP基因的共線性分析Fig.5 Collinear analysis of HD-ZIP gene in Brassica napus genome
3 討 論
HD-ZIP家族基因在功能上非常保守并且參與高等植物的多種生物學過程,目前大多數植物的HD-ZIP 轉錄因子已被鑒定和分析。Zhao等[22]在玉米中共鑒定到55個HD-ZIP家族基因,Yue等[35]在小麥中鑒定到46個HD-ZIP家族基因。但在油菜中還未見相關分析報道,為更加清楚地了解甘藍型油菜中HD-ZIP轉錄因子的功能,本試驗利用生物信息學方法對甘藍型油菜基因組中的HD-ZIP家族基因進行鑒定,并對其基本理化性質、染色體定位、系統進化樹,保守基序及基因結構等進行了統計分析。結果顯示:在甘藍型油菜中共鑒定到74個HD-ZIP家族的轉錄因子,這些基因在甘藍型油菜19條染色體上均有分布;系統進化樹將這些基因分為4個亞族;74個基因都含有motif 1和motif 2保守基序,共同組成HD-ZIP的保守結構域,這些結果與Zhao等[22]的分析結果基本一致,說明HD-ZIP家族基因在不同物種間比較保守。
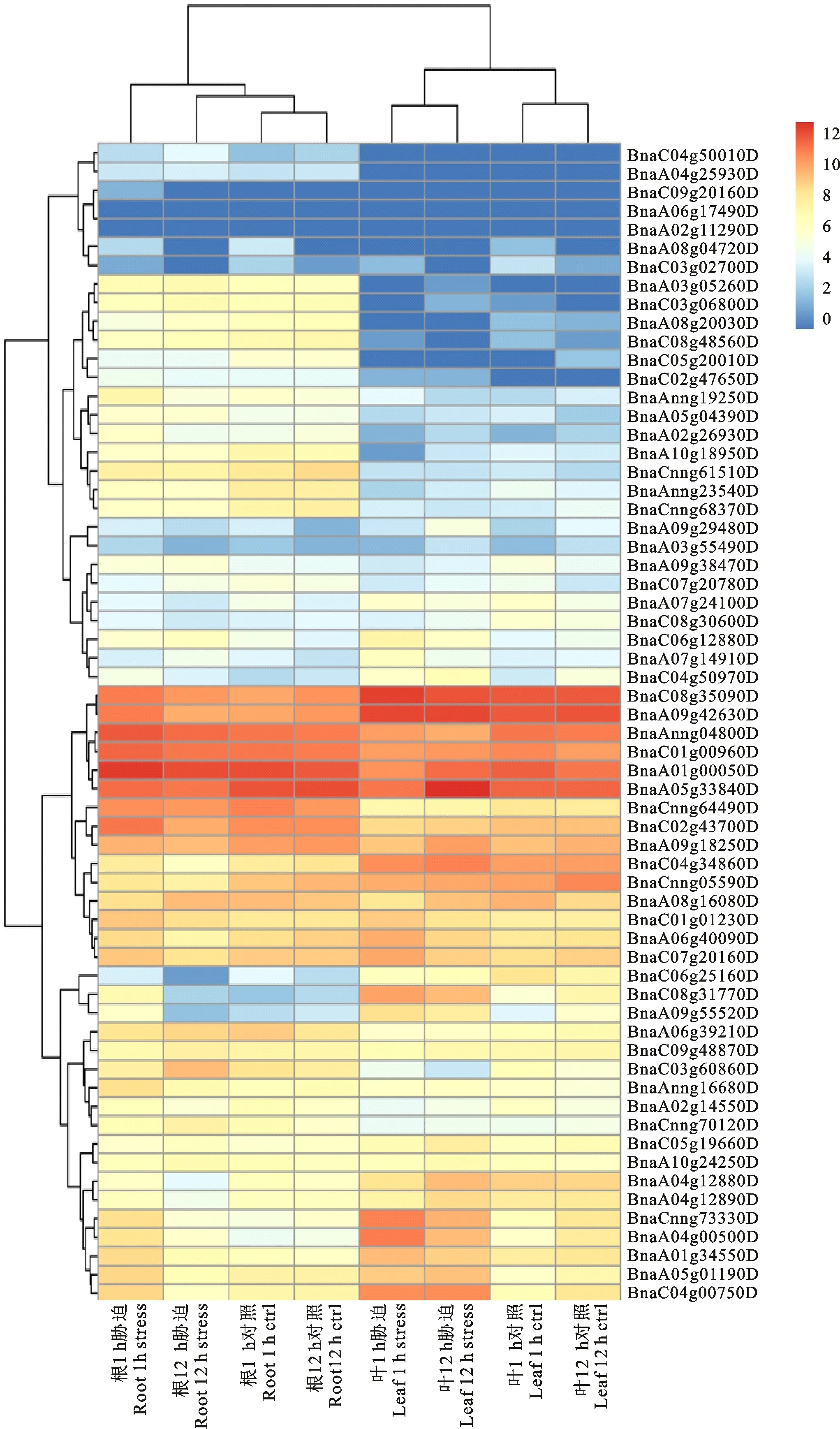
圖6 甘藍型油菜HD-ZIP家族基因的表達分析Fig.6 Expression analysis of HD-ZIP family gene in Brassica napus L.
甘藍型油菜是由白菜和甘藍自然雜交以及后期染色體加倍而產生的異源四倍體,基因組中應該存在多拷貝基因,但是通過共線性分析發現有的基因在甘藍型油菜染色體組中不存在共線性關系,這可能是蕓薹屬植物進化時在加倍復制事件中的不對稱或者丟失所引起的。但是大部分HD-ZIP家族成員基因在甘藍型型油菜的A和C染色體組間的共線性多于在A和C染色體組內的。并且存在共線性關系的基因在系統發育進化樹中也都可被聚為同一類,說明在基因組進化過程中這些基因相對保守,這可能也與這些基因在結構上存在保守的結構域有關。
HD-ZIP轉錄因子是一類較大的基因家族,在生物和非生物脅迫以及植物生長發育過程中具有重要的作用。在鹽脅迫下對甘藍型油菜不同組織HD-ZIP家族基因的表達模式進行分析發現,大部分基因在受脅迫時表達量會升高。Liu等[36]在擬南芥中發現BnaA04g00500D(HB12)基因對非生物脅迫(鹽脅迫)響應,BnaC04g00750D(HB7)基因可以響應干旱脅迫,這些結論與本試驗的分析結果一致,也進一步說明HD-ZIP家族基因在植物發育的調節和對非生物脅迫的響應中起著重要作用。同時也發現有的基因雖然為同源基因,但是在根和葉片中的表達量卻相差很大,比如BnaC06g12880D、BnaA07g14910D和BnaC08g35090D、BnaA09g42630D,前2個基因在甘藍型油菜的根和葉中表達水平低,后2個基因在根和葉中的表達量較高,但是在進化樹分析中發現這兩組基因都屬于HD-ZIP Ⅰ亞家族,這有可能是一些基因在進化過程中功能上出現了 歧化。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
