天蠶素抗菌肽B在魚類疾病控制中的應用
2020-09-07 08:25:06羅陽瑞代奉林黃紅云樊汶樵姜玉松李曉英
中國獸醫雜志 2020年3期
關鍵詞:結構
羅陽瑞,王 晗,代奉林,黃紅云,樊汶樵,姜玉松,李曉英
(重慶文理學院林學與生命科學學院 重慶文理學院水生動物疫病防控研究所,重慶 永川 402168)
抗菌肽是分子質量小、兩親性、帶正電的非特異性物質[1-2],多為α螺旋結構,天蠶素是最早被發現的,目前研究最清晰,效果最顯著的一類抗菌肽。早在1972年,瑞典科學家Boman等[3]從美國希古比天蠶蛹(Hyalophoracecropia)中獲得抗菌肽,并將其命名為Cecropin;隨后,Boman等測定了天蠶素抗菌肽A (Cecropin A) 和天蠶素抗菌肽B (Cecropin B)的一級結構[4]。目前,天蠶素抗菌肽家族已被分為A、B、C、D、E共5個成員,其中Cecropin B具有最高的抗菌活性[5-6]。
魚類是較低等的脊椎動物,缺乏完善的免疫系統來抵抗微生物的侵襲,其皮膚、黏膜存在的免疫活性因子在主動防御中發揮著重要作用[7]。抗菌肽是魚類自身先天免疫的重要組成成分,研究抗菌肽在魚類中的應用,不僅為預防魚類病害開辟新途徑,還能拓展抗菌肽在漁業生產中潛在的應用領域。天蠶素抗菌肽B抗菌譜廣,對多種細菌性疾病有很強抗性[8],對不同種類的病毒也有抵御作用,使其在水產行業有很好的應用前景。
1 Cecropin B的結構與作用機理
Cecropin B是最早從希古比天蠶中分離得到,由35個L-氨基酸殘基組成的抗菌肽,其序列為KWKIFKKIEKVGRNIRNGIIKAGPAVAVLGEAKAL[9],分子量約4 kDa。Hu等[10]認為天蠶素抗菌肽B具有兩親性的α螺旋結構,N端形成親水性的α螺旋結構,C端形成疏水性的α螺旋結構且酰胺化[11]。Srisailam等[12]用NMR(核磁共振)測定Cecropin B1的結構(圖1),發現其N端堿性很強,帶正電荷,親水,富含賴氨酸(Lys)和精氨酸(Arg),N端氨基酸序列比C端對于其活性具有更重要的作用;如N端第2位上的色氨酸,因色氨酸上有苯環,會影響其活性;天蠶素抗菌肽B一般不含半胱氨酸(Cys),所以不能形成分子內的二硫鍵[11-13]。
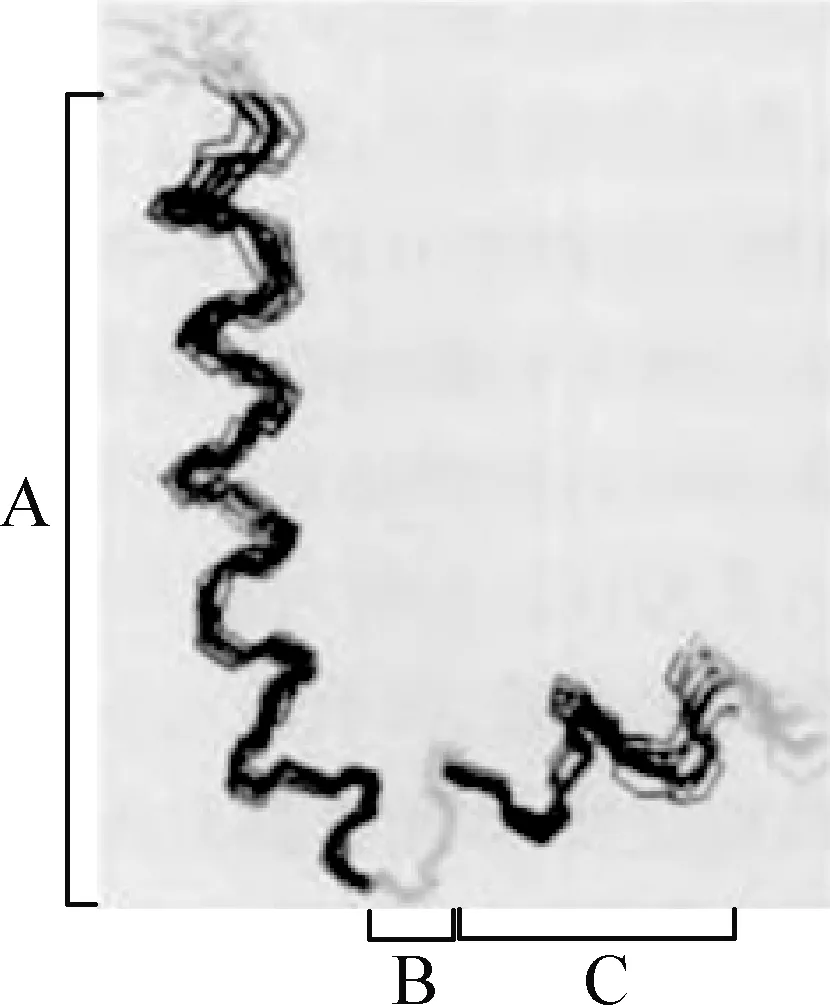
圖1 Cecropin B1的結構示意圖[12]A:N-末端α-螺旋; B:鉸鏈區; C:C-末端α-螺旋
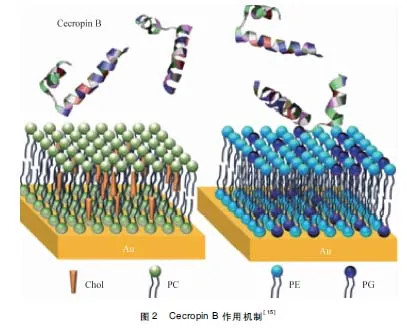
Cecropin B帶正電,與細菌接觸時通過靜電作用,吸附于細菌表面,先破壞細菌細胞壁結構,使大量的Cecropin B選擇性的堆積在細胞膜脂質表面,達到一定濃度時,因為其C端疏水性強,插入細菌胞膜,造成膜缺陷或形成“孔隙”,改變細胞通透性,使細菌內容物大量外流,最終導致細菌死亡[14-16]。其作用機制如中插彩版圖2所示。
2 Cecropin B在細菌病中的應用
引起魚類疾病的致病菌繁多,主要歸屬于氣單胞菌屬(Aeromonas)、假單胞菌屬(Pseudomonas)、維氏氣單胞菌(A.veronii)、殺鮭氣單胞菌(A.salmonicida)、鰻弧菌(Vibrioanguillarum)等。氣單胞菌會引起淡水魚細菌性敗血病、細菌性腸炎、癤瘡等,假單胞菌屬主要危害鯉魚、草魚等,能引起草魚100%的死亡。Sarmasik A等[17]報道重組的Cecropin B和Cecropin P1單細胞克隆入大鱗大麻哈魚(Oncorhynchustshawytscha)胚胎細胞中,表達出的雜合天蠶素抗菌肽對嗜水氣單胞菌(A.hydrophila)、熒光假單胞菌(P.fluorescens)等病原菌有很強的抑菌效果;盧強等[18]將綠色熒光蛋白(GFP)基因與Cecropin B突變體ABP-S1基因進行融合表達,發現對銅綠假單胞菌(P.Aeruginosa)、嗜水氣單胞菌等多種病原菌有抑菌效果;王自蕊等[19]向湘云鯽的鰭部注射0.5 mL嗜水氣單胞菌活菌液后,發現添加Cecropin B組比對照組的存活率更高。林鑫等[20]在感染維氏氣單胞菌的魚類中添加Cecropin B,10 d后,錦鯉的累積死亡率顯著降低;Kjuul等[21]研究發現,Cecropin B對魚的殺鮭氣單胞菌、鰻弧菌、鮭弧菌等多種病原菌都有抑制作用,還發現隨著氯化鈉鹽濃度的升高,Cecropin B的活性會明顯降低;Sallum等[22]用鰻弧菌、嗜鹽弧菌(Vibriohalophilus)和耶爾森菌(Yersinia)感染青鳉魚時,發現Cecropin B的表達量會持續升高,增強青鳉魚的抵抗力,且這3種誘導產生的抗性與它們超微結構的可逆變化相關。Lin X等[23]將天蠶素抗菌肽加入到雜交羅非魚日常飼料中,研究發現,與未添加相比,添加組在生長性能、非特異性免疫能力及對維氏氣單胞菌的抗感染能力方面均有提高。Elaswad A[24]將Cecropin B的基因轉移到鯰魚魚種中,用多毛魚虱(Ichthyophthiriusmultifiliis)這種寄生蟲去感染后,發現轉基因鯰魚存活率比普通魚高;Bugg W[25]將Cecropin B的基因轉移到斑點叉尾鮰(Ictaluruspunctatus)形成轉基因魚,用柱狀黃桿菌(Flavobacteriumcolumnare)和愛德華菌(Edwardsiellaictaluri)去感染,發現轉基因魚存活率都要高于普通魚。一系列的報道表明,Cecropin B在魚類抗細菌感染中發揮著重要的作用。
3 Cecropin B在病毒病中的應用
病毒性疾病會嚴重危害魚類的生長,甚至導致死亡。目前關于Cecropin B對魚類病毒性疾病的研究較少。Lo J H等[26]發現合成肽Cecropin B和模擬Cecropin B結構的模擬肽CF17,能明顯抑制感染性造血壞死病毒(IHNV)、病毒性出血性敗血病病毒(VHSV)、蛇頭棒病毒(SHRV)和感染性胰腺壞死病毒(IPNV)等重要魚類病毒性病原對機體的侵襲。2014年,Chiou P P等[27]人工合成Cecropin B基因并構建CF-17轉基因載體后,植入虹鱒魚精子,隨后用IHNV感染轉基因與非轉基因的虹鱒,發現CF-17轉基因親本魚對IHNV的感染具有顯著的抗性;Chia等[28]發現Cecropin B對致海洋魚類在幼年期大規模死亡的神經壞死病毒(NNV)有較強的抑制作用。
4 展望
Cecropin B抗菌譜廣,對魚類多種細菌性病原體有不同程度的抑制或殺傷作用;鑒于天蠶素抗菌肽結構中沒有二硫鍵,重組表達中不涉及二硫鍵的重構,且鮮有Cecropin B產生耐藥性的報道,證實了Cecropin B成為新型抗生素替代品的潛力。然而從希古比天蠶蛹或其他動物中提取的Cecropin B含量很少,目前一般采取人工合成的方式獲取該抗菌肽;但人工合成抗菌肽價格高昂還容易降解,影響了Cecropin B在生產中的廣泛應用。在具體的研究中,針對重組Cecropin B外源蛋白表達量低、穩定性差等問題,研究者多將2個及以上的目的基因進行串聯表達,取得了較好的進展[29]。隨著基因重組和表達技術的不斷進步,大量獲得外源性Cecropin B逐漸成為現實,Cecropin B必將在實際生產中得到更為廣泛的應用。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
