硝酸根調控植物開花和產量分子機制的研究進展
2020-08-28 16:38:44宮偉余健源張曦單曉昳
生物技術通報 2020年8期
宮偉 余健源 張曦 單曉昳
(1. 北京林業大學生物科學與技術學院,北京 100083;2. 中國科學院遺傳與發育生物學研究所,北京 100101)
選育早熟高產的新品種是作物遺傳育種研究的主攻方向之一。氮素(Nitrogen)既是生物大分子如核酸、脂類和蛋白質等的組成成分,也是植物激素、葉綠素等生長發育必需元素的構成組分。氮素是植物生長發育不可或缺的大量元素,也是調控植物開花時間和種子產量最為重要的營養元素。
在合適的外界環境和內部因子的共同作用下,植物莖頂端的分生組織(Shoot apical meristem,SAM)由營養期轉入生殖期,分化形成花芽,隨后花器官發育、配子體發生、授粉受精,形成種子和果實。作物的生殖生長時期如抽穗、開花以及成熟期與作物產量密切相關。氮素不足迫使作物早衰,產量較低;而在農業生產實踐中大量施用氮肥會導致作物“貪青晚熟”即生殖生長延后,開花成熟推遲。特別是在高緯度地區,生育后期如溫度過低會影響灌漿從而導致作物大量減產。氮素的積累也是促進種子發育和提高作物產量的重要因素,取決于植物的氮素利用效率NUE(Nitrogen use efficiency),主要由氮素攝取效率NUpE(Nitrogen uptake efficiency)和氮素同化效率NUtE(Nitrogen utilization efficiency)共同決定[1]。
硝酸根(NO3-)是植物吸收利用的主要氮素來源,也是植物生長過程中的重要信號分子,在調節植物發育如種子萌發、根系形態建成、植株分支、葉片伸展、成花、種子發育以及植株衰老等過程中發揮重要作用[2]。其中,植物開花和產量與硝酸根轉運、代謝與信號轉導途徑等多方面密切相關。基于此,本文對近年來模式植物擬南芥、水稻和其他主要農作物中硝酸根調控植物開花時間和種子產量的分子機制研究進展進行綜述,重點介紹參與其中的關鍵基因及作用機理,以期為合理利用氮肥、提高氮素利用效率、培育早熟高產的作物新品種提供較為全面的理論參考。
1 硝酸根調控植物開花的分子機制
開花是植物從營養生長到生殖生長過渡的關鍵階段。高等植物開花包括開花誘導、花原基形成和花器官發育3個階段,其進程受到復雜基因網絡的調控。外界環境中不同濃度的硝酸根會對植物開花時間產生不同程度的影響。對模式植物擬南芥的研究表明,硝酸根濃度與開花時間呈U形曲線關系[3]。擬南芥在土培、水培和固體培養基條件下,硝酸根或氮素最適濃度分別為4 mmol/L、1 mmol/L和3-9 mmol/L,此時植株體內硝酸根含量和信號轉導水平最適宜植物開花,因此開花時間最早。濃度高于最適濃度時促進營養生長,延遲開花;而濃度低于最適濃度時,植株營養不足生長遲緩,也會導致開花延遲。目前,已鑒定得到部分參與硝酸根調控開花途徑的關鍵基因,初步解析了植物感知并響應外界不同濃度硝酸根從而調控開花時間的分子機制。但是,所涉及的研究大多沒有覆蓋整個U型曲線,這些調控因子只在某一范圍的硝酸根濃度下發揮作用。
1.1 硝酸根信號通路中參與開花調控的關鍵基因
AtNRT1.1(Nitrate transporter 1.1)定位于根系表皮和皮層細胞,是擬南芥雙親和性硝酸根轉運蛋白及感受器,在根系硝酸根攝取和植株初級硝酸根反應中發揮主導作用[4]。與野生型擬南芥植株相比,AtNRT1.1基因功能缺失突變體chl1-1和chl1-5在長日照或持續光照的條件下均呈現為晚花表型,抽薹后莖生長速度更慢,開第一朵花的時間更晚[5]。AtNRT1.1點突變株系chl1-9喪失了轉運硝酸根的能力,卻仍然保留了正常的初級硝酸根反應。突變體chl1-9開花特征與野生型無顯著性,表明AtNRT1.1調控的擬南芥晚花表型與硝酸根轉運無關,可能與其介導的硝酸根信號轉導途徑有關[6]。水稻基因組中存在3個AtNRT1.1同源蛋白,依據序列相似性高低分別命名為OsNRT1.1A、OsNRT1.1B和OsNRT1.1C[4]。其 中,OsNRT1.1A與AtNRT1.1蛋白序列同源性最高,定位于液泡膜上,主要負責感受細胞內的硝酸根并傳遞信號;而OsNRT1.1B與AtNRT1.1的生物學功能最為接近,定位于細胞質膜上,主要負責轉運硝酸根和感受胞外硝酸根信號[4]。突變體osnrt1.1a呈現出發育遲緩和晚熟的表型,而在水稻和擬南芥中分別超表達OsNRT1.1A均可促進植株早花。特別是在高氮條件下,超表達OsNRT1.1A的植株相較于野生種可提早開花2周以上,從而有效縮短了水稻的成熟時間[7]。因此,硝酸根作為信號分子可能通過不同物種中的NRT1.1s蛋白介導的信號途徑調控植物開花(圖1)。
轉錄因子NLPs(Nin-like proteins)蛋白家族成員是硝酸根信號傳導途徑中的關鍵調控因子。在擬南芥中,硝酸根可能通過PLC(Phospholipase C)/InsP3(Inositol 1,4,5-trisphosphate)途徑誘導胞內鈣離子濃度的大幅度升高,從而激活鈣依賴激酶CPKs(Calcium-dependent kinases)家族成員CPK10、CPK30和CPK32。CPK10/30/32進 而 將AtNLP7磷酸化,使其滯留在細胞核內[8]。經磷酸化修飾后的AtNLP7可結合到多個硝酸根轉運基因(NRT1.1、NRT1.2、NRT2.1、NRT2.2和NAR2.1等)、同 化 基因(NIA1、NIA2和GS2等)以及響應基因(HYH、LBD37和LBD38)的啟動子上,啟動初級硝酸根反應[9]。擬南芥單突變體nlp6、nlp7和雙突變體nlp6nlp7均表現出晚花的表型,而且雙突變體開花比單突變體推遲時間更長[10]。水稻中,OsNLP3和OsNLP4是AtNLP7兩個最為接近的同源蛋白。突變體osnrt1.1a中,OsNLP3和OsNLP4的核定位受到明顯抑制;而在氮缺乏的條件下,在水稻原生質體中共表達OsNRT1.1A和OsNLP3或OsNLP4可促進后兩者的核定位[7]。因此,不同物種中的NRT1.1s蛋白可能通過下游的NLPs轉錄調控因子調控植物開花(圖1)。
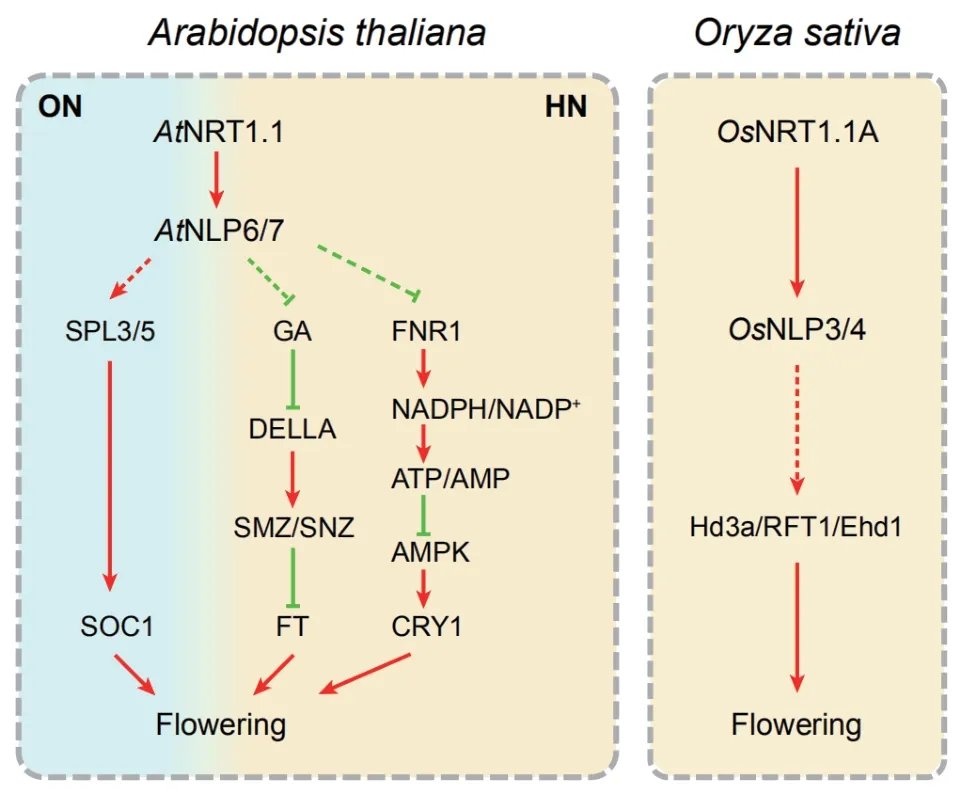
圖1 擬南芥和水稻硝酸根信號調控植物開花的基因網絡示意圖
1.2 硝酸根信號通路與其他開花調控途徑互作基因
對模式植物擬南芥的研究表明,植物開花調控途徑主要包括光周期途徑(Photoperiod pathway)、自主途徑(Autonomous pathway)、春化途徑(Vernalization pathway)和赤霉素途徑(Gibberellin pathway)四大類[11]。CO(Constans)編碼一個具有鋅指結構的轉錄因子,是光周期途徑的核心蛋白[12]。長日照條件下,光敏色素(Phytochrome A,PHYA)、PHYB與隱花色素(Cryptochrome 1,CRY1)、CRY2互作,調節生物鐘基因GI(Gigante)的表達,進而誘導CO的積累,促進擬南芥開花。FLC(Flowering locus C)編碼一個MADS-box家族的轉錄因子,是自主途徑和春化途徑中關鍵開花抑制因子[13]。春化作用負調控FLC的轉錄和FLC蛋白的表達水平,進而誘導擬南芥開花。其中,VRN1(Vernalization 1)、VRN2和VIN3(Vernalization insensitive 3)等 通 過修飾組蛋白維持FLC在發育后期的低水平表達[14]。自主途徑中,FCA(Flowering control locus A)、FY、FVE和FLD(Flowering locus D)等是FLC的抑制因子,通過調節其mRNA水平影響開花[15]。赤霉素(Gibberellin,GA)是調節開花的主要激素之一[16]。成花誘導前,SAM組織可檢測到活性GA的積累,而阻斷GA合成或者信號轉導過程均會影響擬南芥開花過程。下游因子FT(Flowering locus T)和SOC1(Suppressor of constans 1)整合上述多種調控途徑,進而作用于花分生組織決定基因AP1(Apetala 1)、CAL(Cauliflower)和LFY(Leafy)等,促進植株開花[17]。
擬南芥中硝酸根信號途徑調控開花的基因網絡相對獨立于光周期、自主、春化和赤霉素等經典開花調控途徑,但又與其中的關鍵基因互作[18](圖1)。CRY1是硝酸根途徑與光周期途徑互作的關鍵因子。硝酸根通過調控FNR1(Ferredoxin-NADP+-oxidoreductase 1)的 表 達 改 變NADPH/NADP+和ATP/AMP的比率,從而影響腺苷酸活化蛋白激酶(Adenosine monophosphate-activated protein kinase,AMPK)的活性[19]。AMPK可調節細胞核內CRY1蛋白豐度和晝夜節律基因以及光周期途徑關鍵基因GI和CO的轉錄水平,從而影響植株開花[19]。高濃度硝酸根以GA信號依賴的方式抑制擬南芥開花。高濃度硝酸根環境中,活性GA的合成減弱,GA信號途徑的底物蛋白DELLA積累,能夠上調其下游轉錄因子CGA1(Cytokinin-responsive gata factor 1)和GNC(Gata transcription factor 21)的表達[6]。AP2類轉錄因子SMZ(Schlafmutze)和SNZ(Schnarchzapfen)的表達水平隨之提高,SMZ和SNZ蛋白通過與FT啟動子結合,抑制FT表達,從而延遲開花[6]。各途徑整合因子SOC1是硝酸根調控開花過程的重要元件。低濃度硝酸根環境下,SAM中SOC1的表達水平較低,引起晚花表型。硝酸根信號途徑下游轉錄因子AtNLP7和AtNLP6也在SAM中表達,可能通過結合到SOC1兩個上游調控基因SPL3(Squamosa promoter binding protein-like 3)和SPL5的啟動子上,影響SOC1表達[10]。
水稻是短日照植物,但是其開花過程的光周期調控機制與擬南芥較為相似。水稻中的Hd3a(Heading date 3a)是與擬南芥中FT功能一致的開花促進因子,其表達在短日照下被誘導,促進植株開花[20]。RFT1(Rice flowering locus T1)與Hd3a同源性最高,是水稻長日照條件下的開花促進因子[21]。Ehd1(Early heading date 1)編碼B型反應調控因子,是水稻中特有的開花調控基因。無論在短日照還是長日照條件下,Ehd1均是開花促進因子,分別在Hd3a和RFT1上游誘導二者的表達進而促進開花[22]。上文提到的水稻中感受胞內硝酸根信號的OsNRT1.1A晚花突變體中,Hd3a、RFT1和Ehd1的表達水平均顯著下調,而早熟的OsNRT1.1A超表達植株中上述3個基因的表達水平均明顯上調[7]。上述研究表明,水稻中硝酸根信號途徑與光周期途徑協同作用調控開花抽穗(圖1)。
2 硝酸根調控植物產量的分子機制
硝酸根的攝取和轉運,即根系對硝酸根的攝取、硝酸根在根和莖中的長距離運輸以及硝酸根再分配等過程,主要通過各類硝酸根轉運蛋白完成。硝酸根的同化則由一系列氮素同化酶共同完成,包括硝酸還原酶NIA(Nitrate reductase)、亞硝酸還原酶NIR(Nitrite reductase)、谷氨酰胺合成酶(Glut-amine synthase,GS)和谷氨酸合成酶(Glutamate synthase,GOGAT)等[1]。近年來,利用基因工程策略進行的轉基因研究表明,調控硝酸根轉運、同化和信號轉導相關基因的表達可以影響植物的生長和產量(圖2)。
2.1 硝酸根轉運相關基因與植物產量調控
擬南芥中共有53個NPF(Nitrate transporter 1 /Peptide transporter)家 族 成 員 和7個NRT2家族 成 員。其 中,2個NRT1類 轉 運 蛋 白NPF6.3(NRT1.1)、NPF4.6(NRT1.2)和4個NRT2類轉運蛋 白NRT2.1、NRT2.2、NRT2.4和NRT2.5負 責 從外界吸收硝酸根[23]。3個NRT1類轉運蛋白NPF7.3(NRT1.5)、NPF7.2(NRT1.8)和NPF2.9(NRT1.9)以及NPF2.3在硝酸根長距離轉運過程中發揮作用[23]。4個NRT1類轉運蛋白NPF2.12(NRT1.6)、NPF2.13(NRT1.7)、NPF1.1(NRT1.11)和NPF1.2(NRT1.12)和3個NRT2類 轉 運 蛋 白NRT2.4、NRT2.5和NRT2.7以及NPF5.5在硝酸根循環再利用過程中發揮重要作用[23]。目前,雖有部分研究證明擬南芥中NPF家族和NRT2家族成員可影響氮素利用效率,但未見其調控結實產量的報道。
水稻中有多于80個NPF(NRT1/PTR)家族成 員 和4個NRT2家 族 成 員[23]。雖 然 僅 有 部 分硝酸根轉運蛋白的功能得到解析,但仍有多個成員的功能顯示與水稻結實產量密切相關。其中,NRT2.1、NRT2.2、NRT2.4、NPF6.3(NRT1.1A)、NPF6.5(NRT1.1B)、NPF8.9(NRT1)和NPF2.4定位于根系,負責從外界吸收硝酸根和傳遞硝酸根信號[23]。OsNRT1.1A的突變不但導致水稻開花期延長,同時降低了氮素利用效率和水稻產量;而過表達OsNRT1.1A在不同水稻品種及高氮低氮條件下均可顯著提高水稻生物量和產量[7]。結合前文可知,OsNRT1.1A兼具促進植株早熟高產的雙重效應。在秈稻與粳稻間,OsNRT1.1B蛋白一個氨基酸的區別導致二者氮素利用效率和產量產生顯著差異[24]。秈稻具有更高的硝酸鹽吸收及轉運活性和硝酸根同化能力,在低氮和正常施氮條件下產量均顯著高于粳稻。根系微生物組分析進一步表明,OsNRT1.1B在水稻根系微生物構成中發揮關鍵作用。與粳稻相比,秈稻根系富集的微生物類群更加多樣化,微生物功能與氮素代謝更加相關,這可能是導致其氮素利用效率高于粳稻的重要原因之一[25]。NPF2.2、NRT2.3a、NRT2.3b、NPF6.5和NPF2.4參與硝酸根從根部到莖部的轉運[23]。OsNRT2.3b主要定位于韌皮部細胞的細胞質膜,在其胞質側具有一個通過pH感受機制調節硝酸根轉運活性的結構域[26]。超表達OsNRT2.3b可提高植物pH平衡緩沖能力和對多種礦質元素的吸收能力。因此在不同氮素條件下,OsNRT2.3b超表達水稻植株的NUE和產量均大幅提高[26]。OsNPF7.3定位于液泡膜,在硝酸根循環再利用過程中發揮作用。超表達OsNPF7.3不但可以促進水稻的生長,還可以促進生殖生長后期氮素由葉片向籽粒的轉運從而提高作物的產量[27]。OsNAR2.1是NRT2家 族 成 員OsNRT2.1、OsNRT2.2和OsNRT2.3a等的互作因子,調控不同氮素條件下硝酸根的攝取[28]。轉基因水稻中,以OsNAR2.1基因啟動子分別調控OsNRT2.1和OsNAR2.1的表達均可大幅提高植株的NUE和產量[29-30]。

圖2 植物中硝酸根調控種子產量的相關基因總結
其他主要農作物中,玉米中有51個NPF和3個NRT2家族成 員;大豆中有96個NPF和3個NRT2家族成員;小麥中有7個NPF和5個NRT2家族成員;高粱中有67個NPF和4個NRT2家族成 員;谷 子 中 有67個NPF和5個NRT2家 族 成員[31-32]。對于上述硝酸根轉運蛋白的研究多集中在基因家族系統進化分析方面,生物學功能研究尚不深入,調控產量相關機制更不多見。高濃度硝酸根環境下,超表達小麥中高親和硝酸根轉運蛋白TaNRT2.1或TaNRT2.1及其輔助蛋白TaNAR2.1后,轉基因擬南芥角果干重和整體生物量均較高[33]。小麥中另外一個NRT2家族成員TaNRT2.5也需要與TaNAR2.1互作,在根系硝酸根吸收和花期硝酸根循環再利用過程中發揮轉運功能。超表達TaNRT2.5-3B的小麥植株生物量和籽粒產量均大幅提高,而其RNAi植株產量顯著低于對照植株[34]。
2.2 硝酸根同化相關基因與植物產量調控
植物從外界環境中攝取硝酸根后,首先在胞質中被NIA轉化為亞硝酸根(NO2-),隨 后 轉運到葉綠體或質體中被NIR1還原為銨(NH4+)。擬南 芥 中 有2個NIA基 因(AtNIA1和AtNIA2)和1個NIR基因(AtNIR),而水稻中有3個NIA基因(OsNIA1和OsNIA2等)和兩個NIR基因(OsNIR1和OsNIR2)[23]。對NIA和NIR基因突變體及超表達植株的研究表明,上述基因的表達水平影響植株的氮素攝取和利用效率,但是對植株的生長和產量沒有顯著作用[35]。
銨由GS和GOGAT進一步同化,其中GS將銨轉化為谷氨酰胺,GOGAT將其進一步轉化為谷氨酸。在高等植物中,存在兩條銨同化途徑,一條定位于胞質中,受GS1和NADH-GOGAT調控,主要參與根部銨根代謝和葉片衰老進程中氮素的重新利用;另一條定位于質體中,受GS2和Fd-GOGAT調控,主要負責同化質體中硝酸根還原形成的銨和光呼吸產生的銨根[1]。
GS1在擬南芥和水稻中分別有5個成員(GLN1;1、GLN1;2、GLN1;3、GLN1;4和GLN1;5)和3個成員(OsGS1;1、OsGS1;2和OsGS1;3);GS2在擬南芥和水稻中均只有一個成員(AtGS2和OsGS2)[23]。相 比GS2,GS1在 種 子 發 育 調 控 中起到更為重要的作用。不同氮素條件下,擬南芥gln1;1/gln1;2/gln1;3三突變體葉中氨基酸和銨根的含量更高,而氮素再利用效率以及多個產量指標如種子總產量、單個種子重量和收獲指數等均顯著低于野生型植株[36]。在煙草中超表達擬南芥的GLN1;4基因,可提高植株在低氮條件下的GS活性、Glu含量以及種子產量[37]。水稻GS1基因突變體也有不同程度的產量降低。OsGS1;1在成熟葉片和韌皮部中表達豐度最高,其突變體成熟期的株高明顯低于野生型,而且籽粒灌漿受到抑制導致穗粒體積較小[38]。OsGS1;2以銨根依賴的方式在根部皮層細胞表達,其突變體分蘗數和穗數與野生型相比明顯減少[39]。過表達OsGS1;1或OsGS1;2的水稻植株雖然GS活性升高,但是增產效果并不顯著,僅有個別研究表明在特定的生長環境(人工生長箱)中超表達OsGS1;2可提高水稻在高氮條件下的產量[40]。
擬南芥中,Fd-GOGATs分別有2個同源基因GLU1和GLU2編碼,前者在葉片中豐度較高,而后者主要在根中表達;NADH-GOGAT僅有GLT一個基因編碼[23]。與擬南芥相反,水稻中Fd-GOGAT有一個基因編碼,而NADH-GOGAT有兩個基因編碼(OsNADH-GOGAT1和OsNADH-GOGAT2)[23]。OsNADH-GOGAT2與OsGS1;1組織定位類似,二者協同作用負責將衰老葉片中的氮素經韌皮部轉運到穗中。在大田中,OsNADH-GOGAT2基因的突變導致水稻成熟期每穗小穗數與產量均大幅減少[41]。OsNADH-GOGAT1與OsGS1;2組織定位類似,二者協同作用負責根系中銨根的代謝。OsNADH-GOGAT1基因的突變不但影響水稻苗期根系的發育,同樣影響其在大田中的產量。與對照植株相比,OsNADHGOGAT1突變體每株穗數、每穗小穗數、千粒重等產量指標均小幅下調[42]。在其自身啟動子的調控下過表達OsNADH-GOGAT,轉基因水稻植株谷物重量提升幅度最高可達到80%[43]。
其他主要農作物中,硝酸根同化相關基因功能研究主要集中在GS1和GS2家族成員上。玉米中ZmGln1-3基因定位于葉肉細胞,是GS1家族中調控產量的主要成員[44]。其突變體籽粒數目明顯減少,而超表達植株籽粒數目顯著增加[44]。小麥中,TaGS2基因的4個單元型2Ab、2Ba、2Bb和2Da等與氮素利用和多種農藝性狀密切相關[45]。導入其中一個優良單元型TaGS2-2Ab轉基因小麥,不但氮素利用效率和生物量均明顯高于對照植株,而且穗數、每穗粒數和千粒重等籽粒產量指標均有不同程度的提升[46]。高粱中,GS1家族成員之一的SbGln1.2在葉片中表達活性較高[47]。超表達SbGln1.2的轉基因植株在冬季溫室中生長,產量高于對照植株;而在春季實驗中則無顯著差異。這說明光照等其他環境因素可能會影響GS1的活性,進而調控植物的生長發育[47]。另外,將大豆中GS1家族成員GS15和Gmglnb1分別在百脈根和紫花苜宿中異源表達,均未發現對產量有顯著影響[48-49]。
2.3 轉錄因子與植物產量調控
越來越多的研究表明除NLPs外,許多其它類型的轉錄因子參與到硝酸根信號通路中連接外界氮素信號和細胞核內響應基因的表達。但是,僅有少數類型的轉錄因子與植物產量的調控關系得以闡明。
Dof(DNA binding with one finger)蛋白是植物特有的一類轉錄因子。其N末端保守的單鋅指Dof結構域是既與DNA結合又和蛋白相互作用的雙重功能域;C末端序列多變,是特異轉錄調控結構域。擬南芥、水稻和玉米等多個物種中的Dof轉錄因子家族成員均在植物增產中發揮重要調控作用。水稻中的Dof轉錄因子OsRDD1(Dof daily fluctuations 1)可通過促進包括氮素在內的各類無機離子的轉運和吸收提高產量[50]。反義OsRDD1轉基因水稻籽粒大小和千粒重均小于對照植株[51],而超表達OsRDD1水稻的氮素響應能力和每穗粒數、單株粒數及單株總粒重等產量指標均有不同幅度的提高[51]。玉米中Dof轉錄因子ZmDof1可平衡植株中N/C代謝。利用組織特異性啟動子rbcS1調控ZmDof1在小麥中的表達,轉基因植株的氮素利用效率、生物量和產量均有顯著增加[52]。同時,在擬南芥和水稻中分別超表達ZmDof1均可提高植株在低氮條件下的氮素利用效率和營養生長[53-54]。此外,研究人員還將擬南芥的AtDof1.7基因轉入煙草中進行過量表達。低氮條件下,轉基因植株中總蛋白、葉綠素、蔗糖和葡萄糖含量以及參與碳氮代謝的多種酶類的活性明顯高于野生型植株,種子產量也得到提高[37]。
NAC(NAM,ATAF and CUC)蛋白也是植物特有的一類轉錄因子,其N端具有一個約由150個氨基酸組成的保守NAC結構域,C端具有一個高度變異的轉錄激活區。TaNAC2-5A是小麥中一個受硝酸根誘導的NAC調控因子[55]。超表達TaNAC2-5A不但可以增強小麥根系吸收氮素的能力,還可以促進氮素向籽粒的轉運進而提高籽粒產量[55]。進一步分析表明,TaNAC2-5A可直接與硝酸根轉運蛋白TaNRT2.5-3B、TaNRT2.1-6B、TaNPF7.1-6D和谷氨酰胺合成酶TaGS2-2A等編碼基因的啟動子結合,并調控相應基因的表達[34,55]。因此,TaNAC2-5A可直接通過調控硝酸根的吸收和同化促進小麥產量的提高。
NF-Y是一類結合CCAAT-box的重要轉錄因子,包括NF-YA、NF-YB和NF-YC三個亞基。氮饑餓或磷饑餓的條件可誘導小麥中多個TaNF-YA基因的表達[56]。過表達其中的一個成員TaNF-YA-B1后,轉基因小麥側根分枝、硝酸根和磷酸根轉運蛋白表達水平以及氮素吸收能力顯著增強[56]。同時,不論在低氮還是低磷條件下,轉基因小麥的穗數和籽粒產量與對照相比均有提高[56]。因此,TaNF-YA-B1可有助于投入較少的肥料達到高產的目標。
研究人員最近還鑒定了一個負調控小麥硝酸根吸收和籽粒產量的轉錄因子TabZIP60。與正調控轉錄因子不同,硝酸根可抑制TabZIP60的表達[57]。利用RNAi 技術下調TabZIP60的表達后,NADHGOGAT酶活性增強,轉基因小麥的側根分枝、硝酸根吸收和穗數和籽粒產量均不同程度的增加;而超表達植株的表型與之相反[57]。ChIP-qPCR和EMSA實驗表明,TabZIP60可結合到TaNADH-GOGAT-3B基因啟動子中的ABRE結構域并負調控其表達[57]。進一步探究表明,將TaNADH-GOGAT-3B在超表達TabZIP60-6D的轉基因株系中過表達,可恢復其穗數和產量減少的性狀[57]。因此,TabZIP60影響小麥NUE和產量主要是通過其對TaNADH-GOGAT表達的負調控實現的。
3 總結與展望
綜上所述,植物通過感知外界環境硝酸根濃度變化、傳遞硝酸根信號以及有效地吸收分配硝酸根到不同的組織器官中等多種方式調節開花時間和種子產量。其中每一個步驟都受到外界因子和內在遺傳調控網絡的復雜調控。目前,已經對模式植物擬南芥和水稻以及部分農作物中促進早熟或高產的硝酸根感受器、轉運蛋白、代謝調控基因和信號轉導因子等的作用機理進行了闡釋(表1)。但是,在深入挖掘分子機制和應用于生產實踐等方面仍然存在多個挑戰。
許多氮素轉運和代謝蛋白具有不同的表達模式及組織、細胞或亞細胞定位特征。精確動態地調控這些基因的時空表達對合理發揮其生物學功能至關重要。目前,大多數轉基因研究中均采用CaMV 35S和Ubiquitin等組成型啟動子驅動目的基因表達,進而提高NUE和植物產量。鑒于氮素轉運和代謝蛋白的復雜性,在后續研究工作中可選擇組織特異性啟動子或誘導型啟動子調控基因表達,進而更為精準有效地改善NUE。例如,通過低氮誘導的啟動子調控硝酸根轉運蛋白的表達進而增強植株在低氮環境中的硝酸根吸收能力;通過衰老誘導啟動子調控硝酸根再利用蛋白的表達從而促進成熟期硝酸根從葉片向種子中的轉運;通過根部或光誘導特異性啟動子調控根系或葉片中硝酸根轉運和代謝基因的表達等。
增強硝酸根的吸收和轉運,在提高產量的同時可能會推遲開花而延長成熟時間。因此,具有早熟高產雙重效應的調控因子還不多見。對擬南芥AtNRT1.1和水稻OsNRT1.1A的研究表明,多種作物中的NRT1.1s可作為調節氮素利用和促進早熟高產的重要候選基因(圖3)。另外,水稻中OsNRT1.1A和OsNRT1.1B存在較大的功能異化,也提示我們同一作物中NRT1.1s的不同拷貝可在植物環境適應性中發揮重要調控作用。除NRT1.1s外,轉錄組學、蛋白組學、代謝組學和表型組學等多組學聯用將為在今后工作中大力挖掘更多的雙重效應調控因子提供有力手段。
硝酸根作為信號分子調控植物生長,植物激素主要生物學功能之一也是調節植物發育。長期以來,人們對硝酸根和植物激素信號通路的解析大多分開進行,從而導致對營養元素和激素互作協同調控植物發育機制的理解比較有限。除了上文提到的開花調控外,根系形態建成和植株分枝性狀與硝酸根利用和產量性狀密切相關。為了適應土壤中硝酸根非均勻供應條件,植物根系表現出較強的可塑性反應,合理利用養分,維持適當生長狀態和生理功能。同時,生長素、乙烯和脫落酸等3類激素在根系形態建成過程中發揮調節作用。分枝不但是植物株型的重要決定因子,也是與作物產量直接相關的生物性狀。生長素、細胞分裂素和獨腳金內酯等三類激素互作,共同調控植株分枝過程。因此,深入探究硝酸根和激素信號協同調控植物生長的分子機制,并進一步挖掘其中的核心作用元件,將是未來重要研究方向之一。
總之,揭示硝酸根相關重要基因的分子調控機制,并通過基因工程的手段進行遺傳改良是改善植物開花及產量性狀的可行方法。目前,除繼續利用單雙子葉模式植物進行基因功能解析外,仍亟需將模式植物的研究成果延伸到其他農作物品種中,尋找同源基因或者探究類似的調控機制,為合理利用氮肥、提高氮素利用效率和培育早熟高產作物新品種發揮更大的應用潛力。
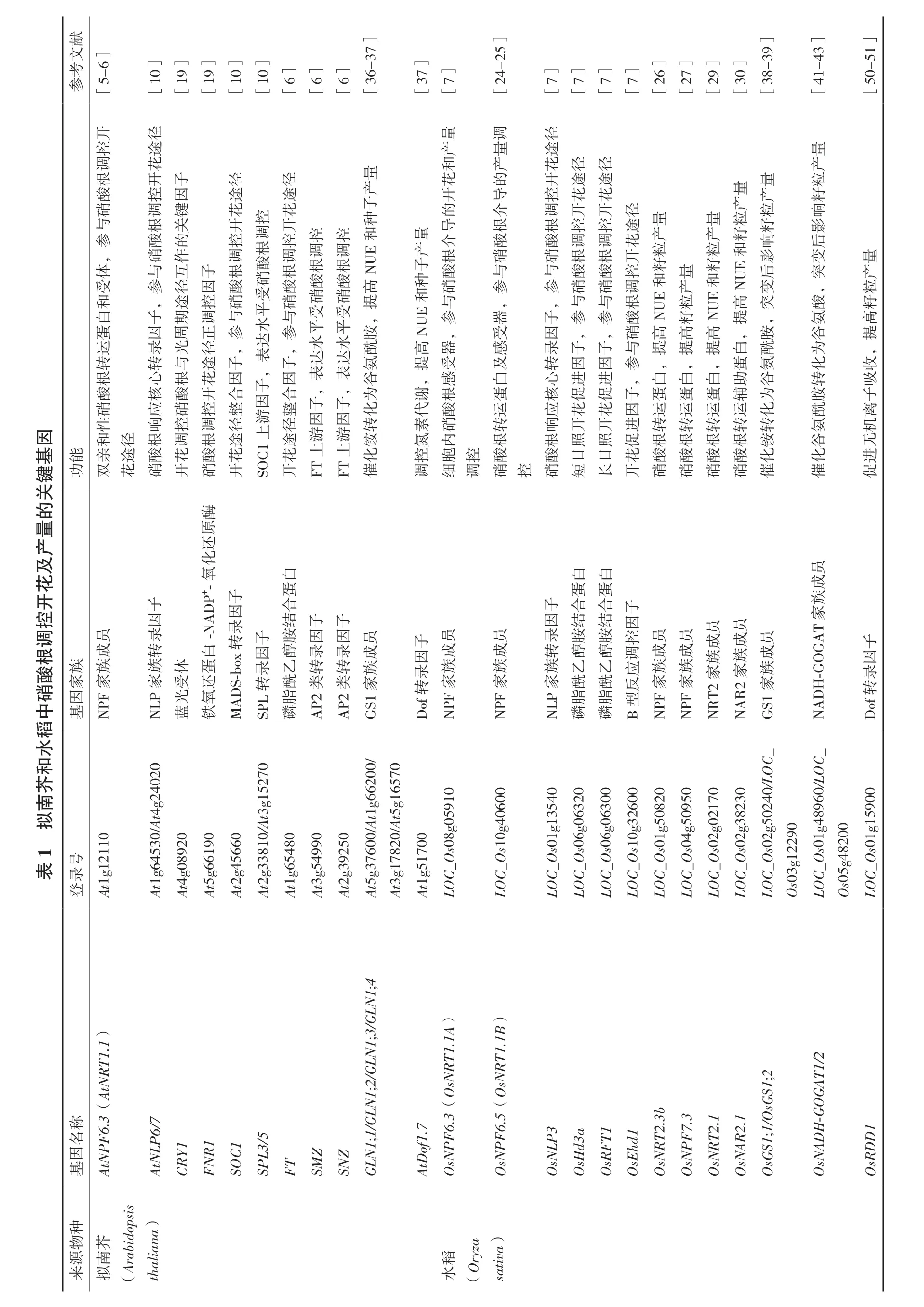
獻文考參[5-6][10][19][19][10][10][6][6][6][36-37][37][7][24-25][7][7][7][7][26][27][29][30][38-39][41-43][50-51]開控徑途量產調量徑途量因基鍵調花根開子徑徑量酸控因途途產硝調鍵花花子與根關開控開種參酸的控調控控控,硝作調根調調調體與互子根酸根根根NUE和受參徑因酸硝酸酸酸高和,途控硝受硝硝硝提白子期調與平與受受蛋因周正參水參平平,胺運錄光徑,達,水水酰轉轉與途子表子達達氨根心根花因,因表表酸核酸開合子合,,谷為硝應硝控整因整子子化性響控調徑游徑因因轉和徑根調根途途游游能親途酸花酸花花銨化功雙花硝開硝開SOC1上開FT上FT上催和產花徑徑花的開途途開導控花花量量產的介調開開徑產粒量導根根控控途量量粒產籽籽介酸酸調調花產產粒粒響子根硝硝根根開籽籽影種酸與與酸酸控量后NUE和硝參參硝硝調產NUE和變與,,與與參器子參參根粒高突酸NUE和籽NUE和提,,受因,,硝高高高,胺高器感錄子子與提提提白酰提受及轉因因,感白心進進參,,,蛋氨,白白白助谷謝根蛋核促促子蛋蛋蛋輔為代酸運應花花因運運運運化素硝轉響開開氮內根根照照進轉轉轉轉轉促根根根根銨控胞控酸酸日日花酸酸酸酸化調細調硝控硝短長開硝硝硝硝催產粒籽響影后變突,酸氨谷為化轉胺酰氨谷化催量產粒籽高提,收吸子離機無進促關的量產及花開控調根酸硝中員成族族家因基NPF家酶原還化白子子蛋因因合錄-NADP+-氧錄結子子轉子胺因因員族白因醇錄錄成NLP家體蛋受還錄乙轉酰轉族光氧脂藍鐵MADS-box轉SPL轉磷AP2類AP2類GS1家子員因成錄族Dof轉NPF家員成族NPF家白白子蛋蛋因合合結結子錄因員員員員轉胺胺控成成成成員族醇醇調族族族族成NLP家乙乙酰酰應族脂脂反磷磷B型NPF家NPF家NRT2家NAR2家GS1家員成族NADH-GOGAT家子因錄Dof轉稻水和芥南1 擬表登At1g12110號錄At1g64530/At4g24020 At4g08920 At5g66190 At2g45660 At2g33810/At3g15270 At1g65480 At3g54990 At2g39250 At5g37600/At1g66200/At3g17820/At5g16570 At1g51700 LOC_Os08g05910 LOC_Os10g40600 LOC_Os01g13540 LOC_Os06g06320 LOC_Os06g06300 LOC_Os10g32600 LOC_Os01g50820 LOC_Os04g50950 LOC_Os02g02170 LOC_Os02g38230 LOC_Os02g50240/LOC_Os03g12290 LOC_Os01g48960/LOC_Os05g48200 LOC_Os01g15900基AtNPF6.3(AtNRT1.1)稱名因AtNLP6/7 CRY1 FNR1 SOC1 SPL3/5 FT SMZ SNZ GLN1;1/GLN1;2/GLN1;3/GLN1;4 AtDof1.7 OsNPF6.3(OsNRT1.1A)OsNPF6.5(OsNRT1.1B)OsNLP3 OsHd3a OsRFT1 OsEhd1 OsNRT2.3b OsNPF7.3 OsNRT2.1 OsNAR2.1 OsGS1;1/OsGS1;2 OsNADH-GOGAT1/2 OsRDD1種物芥源南來擬(Arabidopsis thaliana)稻水(Oryza sativa)
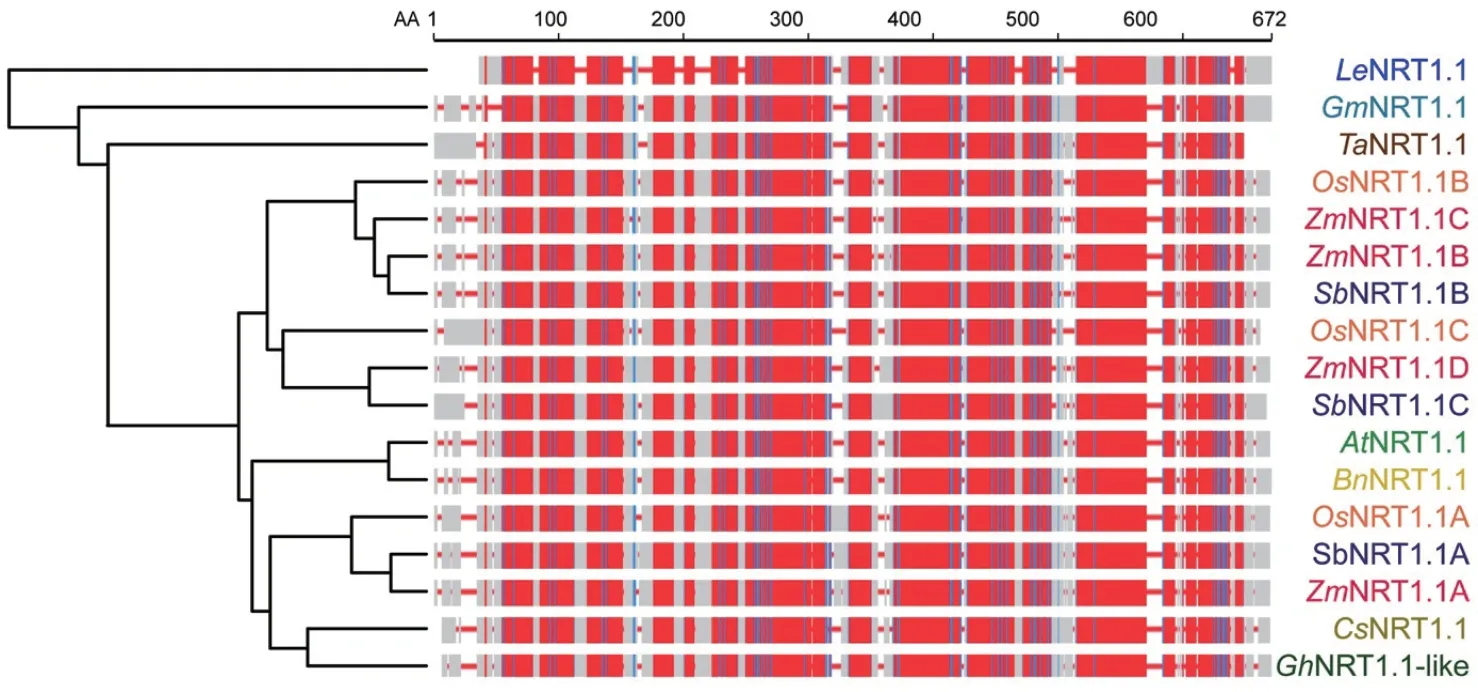
圖3 多種植物中NRT1.1s進化樹分析和序列比對圖
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
