瀕危植物對節白蠟遺傳多樣性的cpSSR分析
2020-08-28 08:58:00鄭鵬麗程建如周明芹
西南農業學報 2020年7期
鄭鵬麗,程建如,周明芹
(長江大學園藝園林學院,湖北 荊洲 434025)
【研究意義】對節白蠟(FraxinushupehensisChiú.Shang et Su)又名湖北白蠟,為木犀科白蠟屬的落葉喬木,其生長緩慢、壽命長、樹齡可達2000年以上,既是珍貴的用材樹種,又是優美的園林綠化樹種,還是極佳的盆景、根雕素材,被譽為“活化石”、“盆景之王”[1]。對節白蠟野生資源僅分布于鐘祥市與京山縣交界的大洪山余脈及周圍相鄰地區,分布范圍狹窄,種群規模小,野生資源稀少,為中國第二批珍稀瀕危保護植物[2],被列入《中國植物紅皮書》。【前人研究進展】目前, 國內外對對節白蠟研究主要集中在生物學特性[3]、人工栽培技術[4-6]、化學成分及藥理作用與應用[7-8]等方面,遺傳多樣性與野生資源保護研究很少。【本研究切入點】葉綠體微衛星(chloroplast simple sequence repeat,cpSSR)是一種新型的標記技術,它不僅有SSR的優點,還能兼顧到cpDNA獨立進化的特點[9],目前已被廣泛應用到植物種群遺傳多樣性[10]、種群結構分析[11]、種群分類[12]、系統地理分布[13]等方面研究。【擬解決的關鍵問題】本研究以來自京山和鐘祥兩地的10個對節白蠟天然居群共196個個體為材料,采用cpSSR分子標記方法對對節白蠟居群野生資源進行了遺傳多樣性分析,旨在從分子水平上揭示10個居群間對節白蠟的親緣關系,為對節白蠟的生物多樣性保護提供參考,并為cpSSR等分子技術在對節白蠟等瀕危植物上的進一步研究提供科學依據。
1 材料與方法
1.1 植物材料
本研究所用材料分別采自京山和鐘祥兩地的客店、雞鳴寺、大口林場、虎爪山、孫橋、空山洞、雁門口、觀音巖、永興及楊集10個對節白蠟天然居群共196個個體。隨機從距離至少10 m的成樹上采集5 g當年生嫩葉,加適量變色硅膠后封口保存備用,種群編號及樣品數分別為KD(16)、JMS(36)、DKL(6)、HZS(31)、SQ(19)、KSD(17)、YMK(21)、GYY(21)、YX(12)、YJ(25)(表1),樣本的數量反映了相應種群的大小。
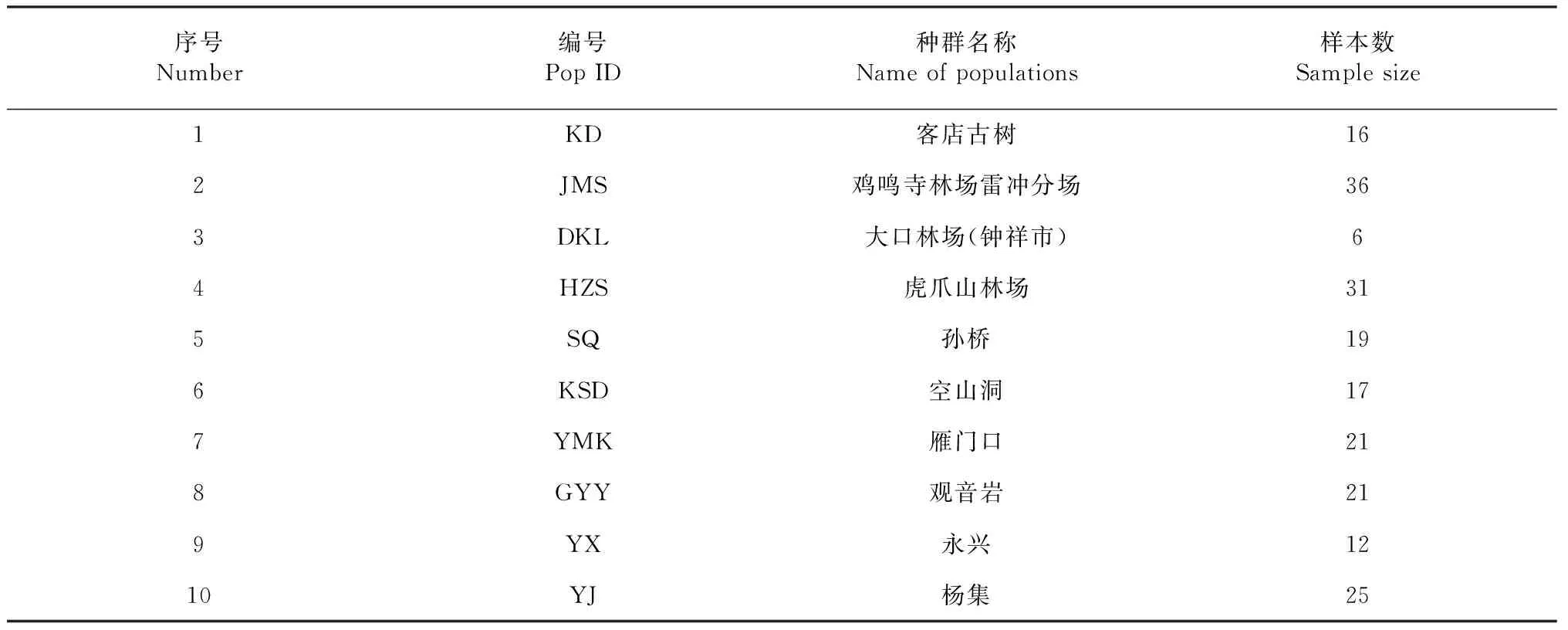
表1 對節白蠟10個居群的樣本及編號
1.2 DNA提取和cpSSR反應體系的建立
稱取0.2 g干燥葉片,采用改良CTAB法[14]提取對節白蠟基因組DNA。cpSSR反應體系為7.5 μl 2×TaqMaster Mix、0.45 μl cpSSR primer(F)、0.45 μl cpSSR primer(R)、1 μl 模版DNA(20 ng/μl)、6.1 μl RNase-Free Water,反應總體系為15 μl。在PCR儀上擴增程序如下:預變性94 ℃ 5 min;變性94 ℃ 30 s,最佳退火溫度72 ℃ 30 s,30個循環;72 ℃延伸7 min;4 ℃保存。
由于cpDNA相對保守,cpSSR突變率較低,cpSSR引物的通用性較高[15]。故本研究從10對雙子葉植物[16]和12對油橄欖[17]的cpSSR引物中篩選出6對能擴增出穩定且有多態性條帶的微衛星引物(ccmp1、ccmp2、ccmp7、ccSSR8、ccSSR12、ccSSR20)用于對對節白蠟10個居群進行擴增,引物由北京天輝遠生物公司合成。
1.3 數據分析
在電泳圖譜中,同一位置的條帶被視作一個位點,根據同一位置上是否有條帶進行統計,有帶的記為“1”,無帶記為“0”。利用軟件 POPGENE 1.31計算居群內及居群間的多態位點百分率(PPL)、等位基因數(Na)、有效等位基因數(Ne)、Nei's基因多樣性指數(h)、香農信息指數(I)、總的基因多樣性(Ht)、物種基因多樣性(Hs)、遺傳分化系數(Gst)及基因流(Nm)、遺傳一致度、遺傳距離。用NTSYS-pc 2.10軟件根據居群間的Nei's遺傳一致度,對對節白蠟10個居群進行UPGMA聚類分析。
2 結果與分析
2.1 cpDNA遺傳多樣性分析
利用POPGENE 1.31對cpSSR標記擴增的結果進行遺傳多樣性參數的統計分析,篩選出的6對cpSSR引物共擴增了40條帶,其中多態性帶有30條,多態位點百分率為75.00 %。在居群水平上,PPL為7.50 %~55.00 %;Na為1.0750~1.5500,平均值為1.2575;Ne為1.0452~1.3124,平均值為1.1712;h值在0.0272~0.1850之間,平均值為0.0974;I值在0.0409~0.2802之間,平均值為0.1440。PPL、Na、Ne、h、I值最高的是雞鳴寺(JMS)居群,最低的居群是觀音巖(GYY)。對節白蠟種在物種水平上,Na、Ne、h、I平均值分別為1.7500、1.2756、0.1720、0.22732(表2)。cpDNA遺傳多樣性分析表明對節白蠟居群遺傳多樣性水平較低。
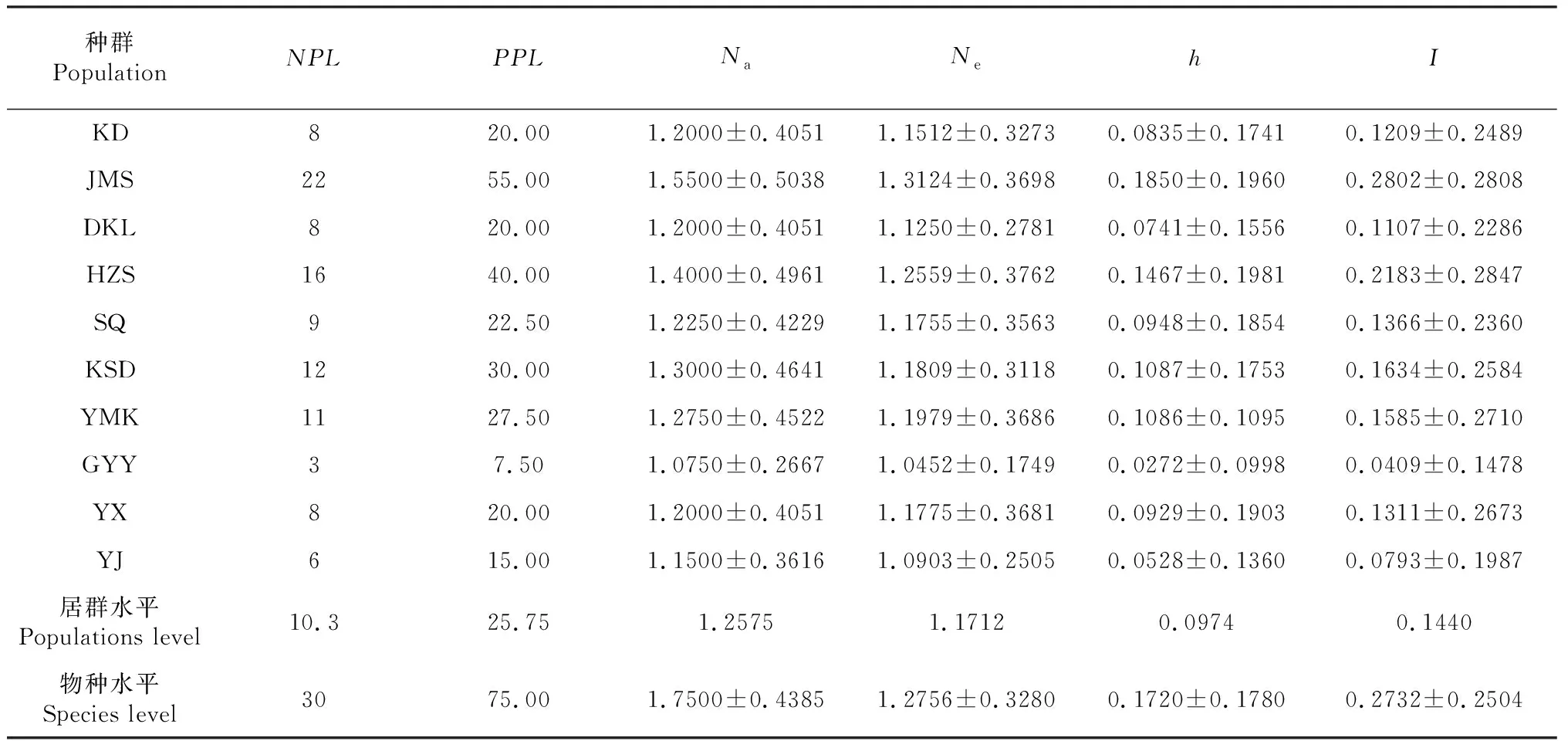
表2 cpSSR檢測對節白蠟10個居群的遺傳多樣性分析
2.2 cpSSR遺傳結構分析
據POPGENE 1.31顯示分析,對節白蠟的Ht為0.1676,Hs為0.0974,Gst為0.4189。用AMOVA1.55軟件包對所得數據進行方差分析,對節白蠟居群間的遺傳多樣性和居群內遺傳多樣性分別占總遺傳多樣性的41.89 %和58.11 %,表明對節白蠟遺傳變異主要存在于居群內。Nm為0.6942,說明居群間基因交流不夠豐富。
2.3 cpSSR遺傳聚類分析
對節白蠟各居群間Nei’s遺傳距離值的變化范圍為0.0329 (KD、GYY)~0.1569(DKL、GYY),平均值為0.0843;遺傳一致度范圍為0.8548(DKL、GYY)~0.9676(KD、GYY),10個居群的遺傳一致度平均值為0.9141(表3)。根據遺傳一致度,用NTSYS-pc 2.10軟件對對節白蠟的10個居群進行UPGMA遺傳關系聚類分析(圖1)。從聚類圖可以看出,10個自然居群聚成4個大類:第I類包括5個居群,分別為客店、觀音巖、虎爪山、楊集和雁門口;第 II類為孫橋和空山洞2個居群;永興居群獨成第III類;雞鳴寺和大口林場2個居群構成第IV類。

表3 基于cpSSR標記的10個居群的遺傳一致度和遺傳距離
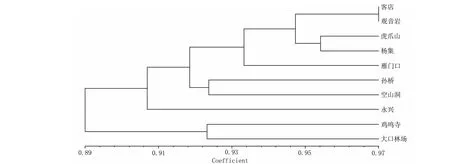
圖1 根據 Nei’s遺傳一致度構建的10個居群UPGMA聚類圖Fig.1 The UPGMA dendrogram of the 10 populations based on genetic identity
3 討 論
近年來,雖一些學者致力于研究瀕危植物多樣性,但這方面的研究仍然相對較少,分子標記等方法的運用將為野生遺傳資源的保護提供更好的研究平臺,在植物遺傳多樣性研究上將發揮更為重要的作用。對節白蠟作為我國特有的瀕危物種,其應用價值極高,野生植物資源受到極大的破壞。因此,了解對節白蠟植物野生居群系統發育,進行遺傳分析及鑒定,從種群基因層面制定出野生植物資源的保護性利用措施極為重要。
由于微衛星兩側的堿基序列具有很強的保守性,因此一個引物可以在另外一個物種上進行穩定擴增,稱之為“種間擴增”。據研究,已開發的SSR引物具有較好通用性[18],可以用于不同物種之間的擴增,如Weising等[19]報道了在煙草中成功擴增的10對cpSSR引物在獼猴桃科、桃金娘科、十字花科、龍舌蘭科、薔薇科、禾本科和豆科中擴增成功。本研究通過篩選已發表的cpSSR通用引物得到一些能穩定擴增的引物,但有部分引物多態性較低,難獲得較高的多態性位點,而多態性高的引物可以得出較好分析結果。這與鄧紹勇[20]對大青屬9個種研究結果相似,表明盡管cpSSR引物的通用性較高,但由于植物親緣關系較遠會導致引物擴增的多態性降低。本研究結果顯示:對節白蠟各居群的PPL平均值為25.75 %,h和I平均值為0.0974和0.1440,其中DKL、YX居群因其分布范圍最小,樣本數最少,群體比較小,致使個體間親緣關系較近,遺傳多樣性水平較低。在物種水平上,PPL值有所提高為75.00 %,而h和I值分別為0.1720和0.2732,均高于居群的平均值,說明對節白蠟在物種水平上具有較高水平的遺傳多樣性。這與桃兒七[21]、榧樹[22]的研究結果一致。
物種的遺傳分化程度可以通過基因流反映出來,基因交換頻繁的物種,其基因流大,居群間的遺傳分化較小[23]。實驗結果顯示:Nm為0.6942(<1),Gst為0.4189(>0.25),表明對節白蠟自然居群間存在著極強的遺傳分化趨勢,這可能是對節白蠟繁育方式為異交,蟲媒傳粉影響了基因流的傳播距離,致使長時間的地理隔離產生,花粉和種子在居群間的傳播較為困難,居群交流頻率較低。此外,對節白蠟居群間的遺傳多樣性和居群內遺傳多樣性分別為41.89 %和58.11 %,遺傳一致度較高(0.8548~0.9676),其原因可能是對節白蠟以前是一個完整的自然居群,由于人類活動的破壞了對節白蠟林,其自然居群數量減少,呈片段化分布,但在大型林場或自然保護區內的大量對節白蠟自然種群得以完整保存,地理距離相距較近的種群仍能有效的進行基因流的交換,使其遺傳多樣性在種群內較高。另一方面,對節白蠟樹較為長壽,種子有深休眠的習性[5],種群天然更新困難,導致其種群間遺傳一致度較高。
對于10個對節白蠟自然居群,提出針對化保護建議:首先,對節白蠟的遺傳變異主要發生在于居群內,因此要做好就地保護,保護其生存環境破不被破壞,保持居群大小和自然更新,并擴大種群規模。其次,對對節白蠟進行遷地保護,從多個居群中收集優質種質資源建立對節白蠟遷地保護林,加強保育遺傳學的研究,更好的保護該物種各水平上的遺傳多樣性。最后,應積極開展其種子繁殖研究和人工繁殖技術和育苗,有效緩解人為過度破壞對天然種群生存的威脅。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
