蒙古沙冬青葉片解剖特征與生態因子的關系
2020-08-20 09:07:04馬紅英李小偉楊君瓏黃文廣王旭康錦蕓
生態環境學報 2020年5期
關鍵詞:生態
馬紅英,李小偉*,楊君瓏,黃文廣,王旭,康錦蕓
1. 寧夏大學農學院,寧夏 銀川 750021;2. 寧夏草原工作站,寧夏 銀川 750021
葉片是植物進行光合作用和蒸騰作用以及呼吸作用等生理代謝活動的重要器官(張詠梅等,2019),與周圍環境關系最為密切,是植物體暴露在環境中面積最大的器官,對外界環境反應最為敏感且具有較大的可塑性,也是植物進行生命活動的主要部位,受生境變化影響顯著(康薩如拉等,2013;吳麗君等,2015)。葉片可以在不同的環境中形成各種適應類型,主要體現在葉片形態學和解剖學上(Zhang et al.,2016;Bosabalidis et al.,2002)。葉片結構對環境敏感、適應性強,且葉片結構的變化是植物適應環境變化的重要機制(He et al.,2018)。國內外學者從葉片解剖結構與生態因子之間的關系進行探討分析(田苗,2016;郭婧宇等,2017;Hoque et al.,2018),研究各種植物與環境的關系,進而得出環境因子對它的影響,以此探尋植物適宜生長發育的環境。因此,通過分析植物葉片的解剖結構與生態因子的關系能夠反映環境對其的影響以及植物對環境的適應性。
沙冬青屬(Ammopiptauthus Cheng f.)是豆科蝶形花亞科中的一屬。沙冬青屬有2種,一種是蒙古沙冬青(Ammopiptanthus mongolicus),是豆科常綠闊葉灌木樹種(蘇志豪等,2018),分布于蒙古國南部以及中國甘肅中東部、內蒙古西部和寧夏中北部地區,另一種是新疆沙冬青(Ammopiptanthus nanus)國家二級保護植物,地方一級保護植物,僅見于新疆喀什地區南部昆侖山與帕米爾交界處(馬松梅等,2012;Gao et al.,2015)。蒙古沙冬青是一種超旱生的珍稀瀕危植物,在中國將其列為國家三級保護植物(段義忠等,2018),也是中國西北荒漠地區唯一的常綠闊葉灌木。蒙古沙冬青抗逆性強,對荒漠區及鹽堿地植被恢復、維持西北荒漠生態系統穩定具有重要的生態價值;另外,蒙古沙冬青也是抗凍蛋白提取的主要材料,在牲畜飼料、化學和藥用等方面具有巨大的經濟價值(Chao et al.,2015;Chen et al.,2016)。蒙古沙冬青生長于西北干旱的荒漠地區,在生長發育過程中會受到風沙、寒冷、干旱、高溫、鹽堿各種不利生態因子的影響(Kosová et al.,2018),且隨著極端氣候出現,環境惡化,生境破碎化加重,棲息地遭到破壞,種群數量不斷縮小,面臨瀕危的現狀(馬松梅等,2012),因此,備受學者關注。目前,眾多研究人員對蒙古沙冬青的研究主要集中在遺傳多樣性與育種(Liu et al.,2013;沈奇等,2019)、無性繁殖(Wei et al.,2011)、基因分析(唐寬剛等,2018;Chen et al.,2011)方面,而對于自然條件下,某一生態主導因子作用下其解剖結構變化的研究還未見報道。
本研究選取 26個樣地的蒙古沙冬青植物葉片作為主要研究對象,借助石蠟切片技術觀察葉片解剖結構,再結合氣候因子與土壤指標,利用冗余分析(RDA)進一步探討其葉片解剖特征與生態因子之間的關系,試圖從多個方面、多個指標評價蒙古沙冬青葉片解剖特征在大尺度上的變化,確定一個或多個與蒙古沙冬青葉片解剖結構有關的生態因子,并確定影響蒙古沙冬青葉片解剖結構的主導因子,主要回答以下幾個問題:(1)研究蒙古沙冬青葉片解剖結構特征;(2)闡明導致蒙古沙冬青葉片結構差異的主導環境因素,探究蒙古沙冬青適應環境的策略,以其為瀕危植物蒙古沙冬青的保護提供理論依據。
1 材料方法
1.1 研究區域概況
本研究區域位于東經 104.79°—107.15°、北緯36.28°—41.77°之間,分別是寧夏與內蒙古西部地區(圖 1),屬典型的大陸性半濕潤半干旱氣候,降雨從南向北依次遞減,且多集中于 7—9月,年降雨量100—400 mm,蒸發量2000 mm以上,地勢南高北低,海拔1014—1883 m,年均溫9.5 ℃,年均輻射 198712 kJ·m?2·d?1,熱量充足;土壤類型從南向北依次為黑壚土、黃綿土、灰鈣土和砂質土。
1.2 樣本的采集及測定
1.2.1 葉片樣本的采集
于2018年7—8月,在研究區內設置26個樣地,每個樣地設置3個10 m×10 m樣方,每個樣方內選取3個生長良好、發育成熟、有代表性的植株,選取其同一部位的葉片(3—5片),裝入帶有FAA固定液的離心管,帶回實驗室。采回的蒙古沙冬青葉片經過取材﹑固定、脫水、透明、透蠟、包埋、切片、展片、貼片、烤片、染色、封藏、烤片等步驟制成永久制片,將制片置于Olympus顯微鏡下觀察。采用Motic Image Advanced 2.0軟件測量并進行顯微照相。
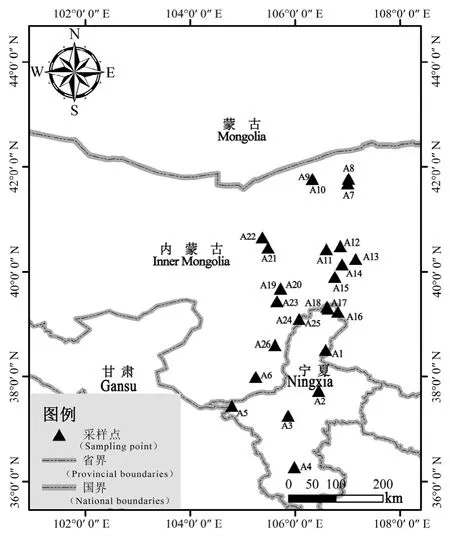
圖1 蒙古沙冬青采樣點詳細分布圖Fig. 1 Detailed map of the sampling points of Ammopiptanthus mongolicus
選擇葉片厚度(Thickness of leaf,TL)、上表皮細胞厚度(Thickness of upper epidermis cell,TUE)、下表皮細胞厚度(Thickness of lower epidermis cell,TLE)、上柵欄組織厚度(Thickness of upper palisade tissue,TUP)、下柵欄組織厚度(Thickness of lower palisade tissue,TLP)、主脈厚度(Thickness of main vein,TMV)共6項葉片結構指標進行測定。
1.2.2 土壤樣本的采集
在設置的每個蒙古沙冬青樣方內同時采集土樣,按五點取樣法取樣,用土鉆在每個樣地內取 0—40 cm土層土壤,剔除其中的植物根系及石塊等非土樣組成部分,帶回實驗室自然風干。將土壤樣品混合均勻并按照四分法取土樣研磨,過2 mm土壤篩后用于土壤養分指標的測定。
土壤有機質(SOC)含量采用重鉻酸鉀外加熱法測定,土壤全氮(TN)含量采用全自動凱氏定氮儀測定,土壤全磷(TP)含量采用 HClO4-H2SO4消煮鉬銻抗比色法測定,土壤含水率(MOIS)采用烘干法測定,土壤pH(pH)采用電位法測定。
1.3 氣候數據的獲取
選取5個氣候因子,即年均降雨量(MAP)、太陽輻射(SRAD)、蒸汽壓(VARP)、年均溫(MAT)、生長季降雨量(GSP)。每一個樣地都使用GPS確定海拔(ALT)、經度(LON)和緯度(LAT)即空間因子。依據世界氣候數據網站(http://www.woridclim.org/),基于調查地點的經度、緯度與海拔數據,分別獲取26個樣地的上述5個氣候因子指標。
1.4 統計分析
用SPSS 21.0對蒙古沙冬青植物葉片解剖結構與生態因子的相關性進行分析。用Canoco 5.0軟件對蒙古沙冬青葉片解剖結構和生態因子進行冗余分析(RDA排序)和繪圖。
2 結果
2.1 葉片解剖結構特征
本研究測量了蒙古沙冬青葉片厚度、上、下表皮細胞厚度、上、下柵欄組織厚度與主脈厚度(圖2),結果顯示,蒙古沙冬青葉片具有超旱生結構,葉片小而厚,且解剖結構為等面葉。由表1可以看出,蒙古沙冬青葉片厚度平均值為385.01 μm,最大值為 (516.02±25.35) μm,最小值為 (278.44±22.13) μm;表皮由單層細胞組成,形態特征基本一致,近長方形或橢圓形,上表皮細胞較細長,排列較下表皮細胞疏松,下表皮細胞近圓狀或不規則狀,排列較緊密。葉片上下表皮細胞厚度平均值分別為:(19.39±1.46) μm 和 (22.90±2.00) μm,上表皮細胞厚度最大值為 (28.77±3.25) μm,最小值為(13.33±1.26) μm,下表皮細胞厚度最大值為(39.81±3.98) μm,最小值為 (15.89±1.85) μm,表明下表皮細胞厚于上表皮細胞,并且下表皮細胞厚度變化幅度高于上表皮細胞厚度變化。葉肉組織分化為上柵欄組織與下柵欄組織,上柵欄組織排列較緊密,其厚度平均值為 (128.47±5.02) μm,最大值為(165.21±8.50) μm,最小值為 (95.78±8.60) μm;下柵欄組織排列較疏松,其厚度平均值為(161.03±6.35) μm,最大值為 (199.28±19.62) μm,最小值為 (116.28±12.35),這表明蒙古沙冬青葉片下柵欄組織厚于上柵欄組織,并且變化幅度也高于上柵欄組織。蒙古沙冬青葉片主脈較突出,主脈厚度平均值為 (277.25±9.23) μm,最大值為 (396.16±29.26) μm,最小值為 (185.94±25.02) μm。
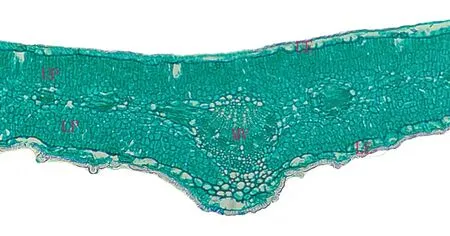
圖2 蒙古沙冬青葉片解剖結構Fig. 2 The anatomical structure of Ammopiptanthus mongolicus leaves
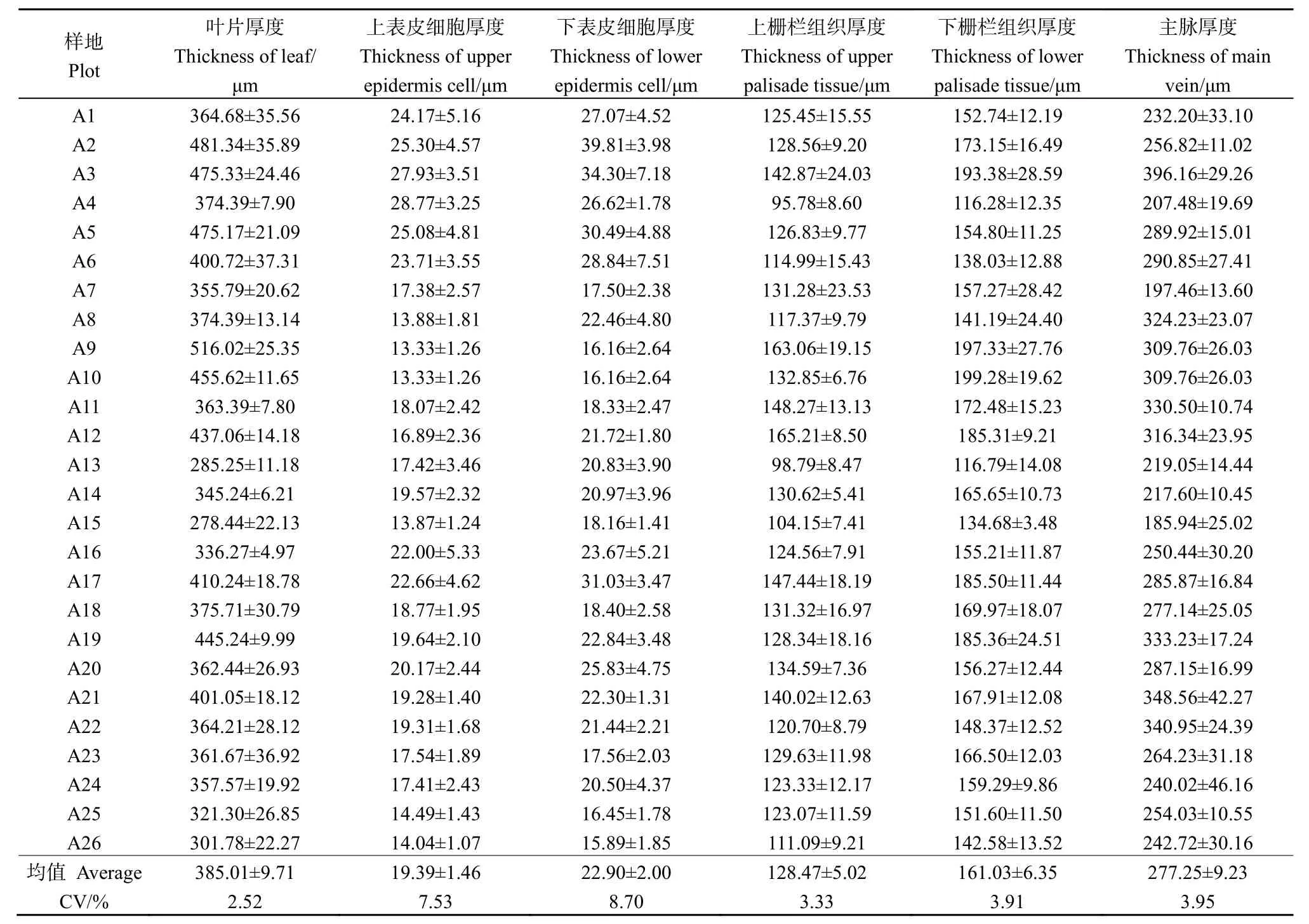
表1 蒙古沙冬青葉片解剖結構指標Table 1 Anatomical structure index of Ammopiptanthus mongolicus leaves
2.2 葉片解剖結構與生態因子的相關性
對葉片解剖結構與生態因子之間的相關性分析結果(表 2)表明,上、下表皮細胞厚度分別與緯度呈極顯著負相關(r= ?0.780,P<0.01)、(r=?0.634,P<0.01);上表皮細胞厚度與年均降雨量呈極顯著正相關(r=0.671,P<0.01),下表皮細胞厚度與年均降雨量呈顯著正相關(r=0.492,P<0.05);上、下表皮細胞厚度也分別與生長季降雨量呈極顯著正相關(r=0.681,P<0.01)、(r=0.518,P<0.01);葉片厚度、上表皮細胞厚度分別與太陽輻射呈顯著負相關(r= ?0.395,P<0.05)、(r= ?0.423,P<0.05);上、下表皮細胞厚度分別與蒸汽壓呈極顯著正相關(r=0.774,P<0.01)、(r=0.686,P<0.01);上表皮細胞厚度與土壤全氮呈極顯著正相關(r=0.506,P<0.01),下表皮細胞厚度與土壤全氮呈顯著正相關(r=0.485,P<0.05)。上柵欄組織厚度分別與年均降雨量、生長季降雨量呈顯著負相關(r= ?0.419,P<0.05)、(r= ?0.412,P<0.05),而下柵欄組織厚度、主脈厚度與生態因子均無顯著相關關系。葉片解剖結構與海拔、經度、年均溫、有機質、土壤水分、土壤全磷、pH無顯著相關關系。
2.3 蒙古沙冬青葉片解剖與生態因子之間的冗余分析
對 26個蒙古沙冬青樣地葉片解剖與生態因子進行冗余分析(圖 3、表 3),結果表明:蒸汽壓(F=8.0,P=0.002)、緯度(F=7.2,P=0.002)、生長季降雨量(F=5.0,P=0.01)、年均降雨量(F=4.7,P=0.012)、土壤全氮(F=2.9,P=0.04)、太陽輻射(F=2.7,P=0.048)對葉片解剖結構具有顯著影響。而年均溫(F=1.9,P=0.144)、經度(F=1.7,P=0.168)、有機質(F=0.6,P=0.562)、土壤 pH(F=0.3,P=0.858)、土壤水分(F=0.3,P=0.862)、土壤全磷(F=0.3,P=0.824)、海拔(F<0.1,P=0.972)對葉片解剖結構影響不顯著。
方差分解結果顯示(圖4),氣候因子、土壤因子和空間因子共同解釋了蒙古沙冬青葉片解剖75.6%的方差,未解釋率為24.4%。其中氣候因子、土壤因子和空間因子對蒙古沙冬青葉片解剖的單獨解釋率分別為 21.1%、17.3%和 10%,氣候因子的影響最大,土壤因子次之,空間因子最小。對 3組環境進行雙因素分析,氣候因子和土壤因子、空間因子和氣候因子、空間因子和土壤因子的共同解釋率分別為65.6%、58.3%和54.5%,三者之間的共同解釋率為27.2%。
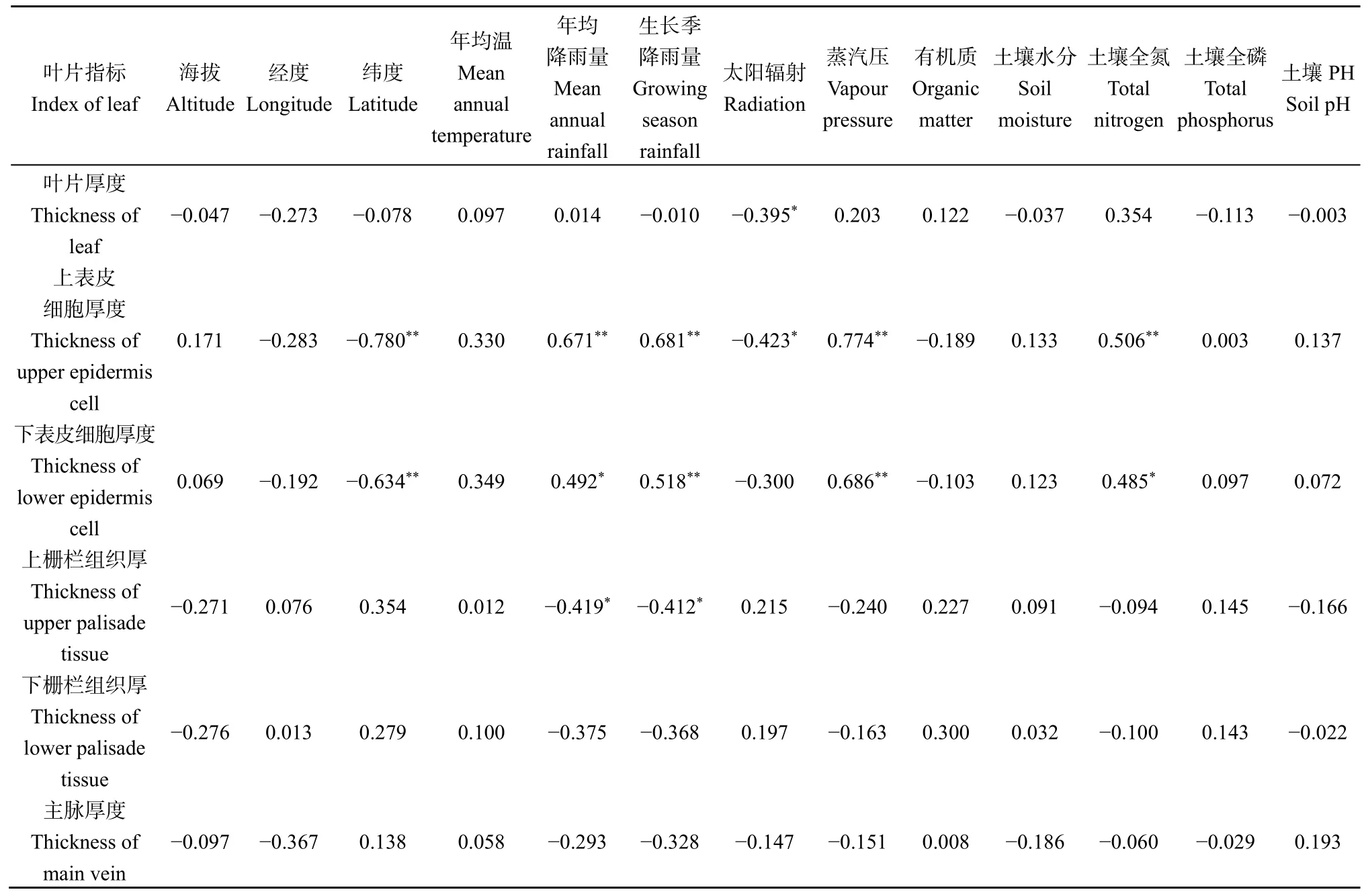
表2 葉片解剖與生態因子的相關性Table 2 Correlation between leaf anatomy and ecological factors
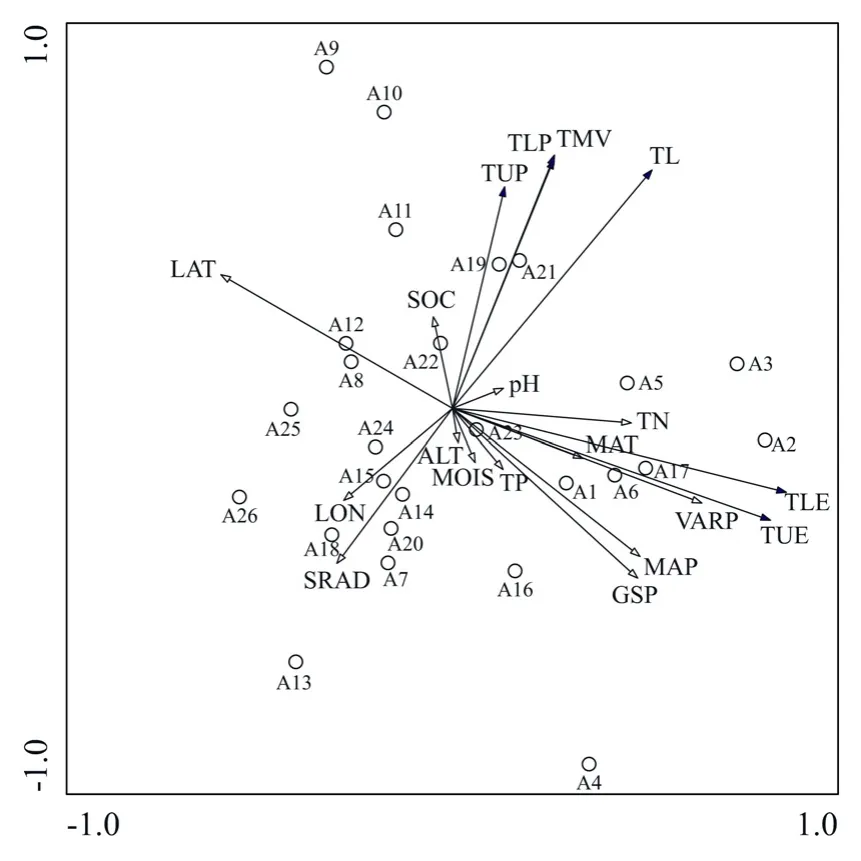
圖3 蒙古沙冬青樣地、葉片解剖與生態因子的RDA排序Fig. 3 Sample plot、leaf anatomy and ecological factor ranking of Ammopiptanthus mongolicus by RDA
3 討論
3.1 蒙古沙冬青葉片解剖特征
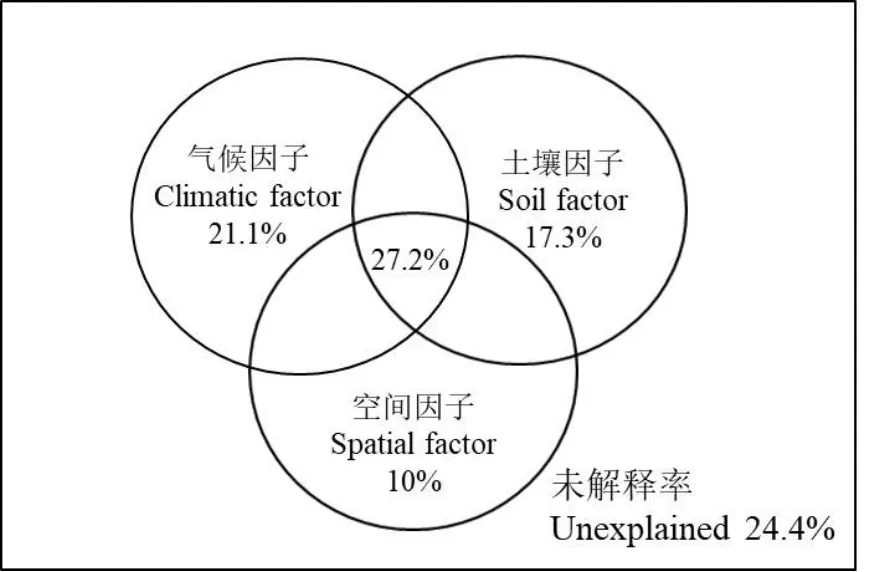
圖4 氣候因子、土壤因子和空間因子對蒙古沙冬青葉片解剖的方差分解圖Fig. 4 Variance decomposition diagram of Ammopiptanthus mongolicus leaf anatomy by climatic, soil and spatial factors
干旱是影響植物生長發育的重要因素之一,不同種植物受自身遺傳特性與外界環境的共同影響,對干旱形成了不同的生態適應對策。為了適應高溫、干旱等逆境環境,植物葉片解剖結構均具有相應的變化特征(孟林等,2011)。有研究表明,長期生長在干旱環境中的植物葉片通常表現出葉片厚度增大、角質層增厚、柵欄組織厚度增加、海綿組織厚度減少或退化等(李蒙蒙等,2016)。也有研究表明,植物適應干旱環境的第一個特點是盡量縮小葉面積以減少蒸騰量,但是不同的植物適應干旱環境所表現出來的特征有所不同(康薩如拉等,2013;吳麗君等,2015)。本研究發現:(1)蒙古沙冬青的葉片橫切面為典型的等面葉,屬于厚葉植物;(2)葉片的上、下表皮均由一層細胞組成,細胞肥大形狀規則,排列整齊緊密無細胞間隙,可以減少葉片的蒸散量;(3)葉肉細胞緊密排列全部柵欄化,貼近上下表皮分布,上柵欄組織厚度較小,細胞垂直排列且較緊密規則,下柵欄組織厚度較大,排列較疏松,海綿組織退化,可以減少水分蒸騰流失;(4)葉脈為平行脈,由維管束與機械組織構成,發達的主脈呈橢圓狀且較為突出,增強了對水分的輸導能力。以上這些特征表明生長于干旱荒漠地區的蒙古沙冬青葉片具有典型的超旱生結構(李蒙蒙等,2016;劉美等,2017;李苗苗,2008),且這與石麗麗等(2018)對蒙古沙冬青和新疆沙冬青的比較研究結果一致。
3.2 蒙古沙冬青葉片解剖特征與環境因子的關系
植物葉片的結構與功能可較好的反映植物對各種環境的適應性(England et al.,2006;Tian et al.,2016)。盡管葉片基本上由表皮、氣孔和葉肉組成,但由于系統發育關系和對特定環境的適應,不同種類葉片在面積、厚度和形狀上表現出明顯的差異(Royer et al.,2008)。研究表明,決定大尺度植物形態結構差異的因素主要為溫度和降雨等生態因子(胡建瑩等,2008);而在小尺度上,植物形態結構差異主要由微環境導致,如地形條件(緯度)、土壤理化性質及其他生態因素(邰建輝等,2011)。植物的形態特征與環境因子的關系,尤其是水熱因子的關系逐漸從定性描述轉變為定量描述(莫億偉等,2011)。環境對葉片的影響主要體現在葉片厚度、上表皮細胞厚度、下表皮細胞厚度、柵欄組織厚度、主脈厚度等方面(王維睿等,2015)。本研究對葉片解剖特征與生態因子的相關性分析表明:生態因子對下柵欄組織和主脈無顯著影響,以往的研究表明光合作用主要發生在完全發育的柵欄組織中(Mendes et al.,2001),而下柵欄組織是由海綿組織發育而來,并未完全發育成熟,所以環境因素對它的影響不顯著(周宜君等,2001);主脈則主要是進行養分與水分的運輸(張文霞等,2018),與環境因子的關系較弱。生態因子主要對蒙古沙冬青上、下表皮影響較大,對葉片上柵欄組織也有一定影響。具體而言,在本研究區域中隨著緯度的升高,環境特征趨向于溫暖,上、下表皮厚度減小,有利于葉片內部的氣體交換,使得外界氣體可迅速向內部擴散(Verboven et al.,2015),而且利于葉片內部的柵欄組織進行光合作用(He et al.,2018)。另外,上表皮細胞還與緯度、年均降雨量、生長季降雨量、蒸汽壓、土壤全氮呈極顯著正相關關系(P<0.01),蒙古沙冬青從北向南分布主要是降雨和溫度的變化,降雨量呈增加趨勢,但海拔逐漸升高,溫度下降,為適應寒冷的環境,上表皮細胞隨之增厚,上表皮細胞增厚保水保護能力增強,且足夠的水分也保證了光合作用,而且也起到抵御寒冷的逆境環境(郭學民等,2015;郝文芳等,2019);與上表皮相比,下表皮位于葉片下方,受環境因子影響較小,因此,環境更多的影響上表皮;土壤理化指標中,只有全氮對葉片解剖特征有較大的影響,蒙古沙冬青是豆科植物,根系發達且可結瘤固氮(郝文芳等,2019),而且賈曉紅等(2006)有研究表明,在干旱和半干旱的草地中,大量灌木生長的地方會加劇灌叢下的養分聚積,從而形成了“肥島”,促進這些灌木在群落中的定居,因此,土壤中的氮素對葉片解剖特征有較大的影響。

表3 蒙古沙冬青葉片解剖與生態因子的RDA排序Table 3 RDA sequencing of leaf anatomy and ecological factors in Ammopiptanthus mongolicus
總體而言,年均降雨量與生長季降雨量對蒙古沙冬青葉片解剖特征有較大的影響,這是由于蒙古沙冬青生長于干旱的荒漠地區,水分是其生長發育的限制性因素(石麗麗等,2018)。本研究結果表明,降雨對上、下表皮細胞和上柵欄組織影響較大,葉片表皮細胞具有保護功能與儲水作用(張香凝等,2011),降雨主要通過影響表皮細胞厚度使其適應不同的水分環境。總之,利用 RDA分析蒙古沙冬青葉片解剖特征與生態因子相關的數據,以及方差分解累計解釋量表明,氣候因子是影響蒙古沙冬青葉片解剖特征的主要生態因子,生態因子中的蒸汽壓、緯度、生長季降雨量、年均降雨量、土壤全氮、太陽輻射的影響較大,表明各環境因子對葉片解剖特征的影響不是單一的,它們共同作用,影響蒙古沙冬青葉片的解剖特征(段義忠等,2018);其中熱量的影響最大,降雨的影響次之,這與田苗(2016)的研究結果基本一致。
4 結論
(1)蒙古沙冬青葉片橫切面為典型的等面葉,表皮肥大,葉肉全部柵欄化,主脈突出,維管束發達,具有旱生結構。
(2)對蒙古沙冬青葉片解剖特征影響最大的是氣候因子,其次為土壤因子,空間因子的影響最小;其中氣候因子中的蒸汽壓、降雨是主要影響因子,土壤因子中的全氮是主要影響因子,空間因子中的緯度是主要影響因子。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
