印加孔雀草對土壤細菌群落多樣性的影響
2020-08-20 09:07:02鄆玲玲張瑞海宋振付衛東王然王忠輝張國良
生態環境學報 2020年5期
鄆玲玲,張瑞海,宋振,付衛東,王然,王忠輝,張國良
中國農業科學院農業環境與可持續發展研究所,北京 100081
外來入侵雜草對生物多樣性、生態環境以及人類生產生活造成了嚴重的影響(Montserrat et al.,2011;萬方浩等,2002;閆小玲等,2014)。在入侵植物與土著植物的種間競爭中,土壤微生物可能起到了重要的“橋梁”的作用,外來植物可能通過改變入侵地土壤微生物群落結構與功能促進自身生長或阻礙共生植物的生長和更新(Yu et al.,2005;Li et al.,2006)。大量研究表明,入侵植物可以通過改變土壤微生物功能類群來改變養分循環及其他環境條件,進而形成有利其競爭的微生態環境,以利于快速形成自身群落,最終實現入侵(Katharine et al.,2006;彭鑫怡等,2019;Niu et al.,2007;宋振等,2016)。入侵植物紫莖澤蘭(Eupatorium adenophorum)根系可從根際土壤中選擇性富集梭狀芽胞桿菌(Clostridium)和腸桿菌(Enterobacter),這些根際微生物的變化使土壤營養成分更有利于紫莖澤蘭生長(Chen et al.,2019);菊科植物黃頂菊(Flaveria bidentis)入侵導致土壤中鏈霉菌(Streptomyces)、中華桿菌(Saprospiraceae)等細菌的豐度增加,而芽孢桿菌(Bacillus)、Solibacterales等細菌的豐度減少,這些微生態環境的改變可能會影響本地植物的生長發育,從而有利于黃頂菊的入侵、競爭與擴張(Song et al.,2017);飛機草(Chromolaena odorata)可以富集入侵地土壤中的土傳致病真菌半裸鐮刀菌(Fusarium semitectum),使其能夠抑制當地植物的生長,促進自身的入侵過程(Vaisakh et al.,2012)。另外,植物之間的競爭作用也會改變土壤細菌群落結構(Garbeva et al.,2004),如閆素麗等(2011)研究發現,黃頂菊與高丹草(Sorghum bicolor×S.sudanense)、向日葵(Helianthus annuus)、紫花苜蓿(Medicago sativa)和黑麥草(Lolium perenne)4種替代植物混合種植可以改變黃頂菊入侵地的微生物群落結構,使其接近于替代植物的土壤微生物類群,這可能是替代植物對黃頂菊具有較強競爭優勢的原因所在;李光義等(2013)研究發現不同搭配方式的白三葉(Trifolium repens)、雜交甜高粱(Sorghum dochna)、雜交狼尾草(Pennisetum alopecuroides)與入侵雜草勝紅薊(Ageratum conyzoides)混種對土壤細菌多樣性產生了不同程度的影響。
印加孔雀草(TagetesminutaL.)又名臭羅杰,屬菊科(Asteraceae),萬壽菊屬(Tagetes),一年生草本植物,原產自南美洲南部,后引種到歐洲、亞洲、非洲、馬達加斯加島、印度、澳大利亞以及夏威夷等地區。在我國境內,2006年首次臺灣地區發現印加孔雀草(Wang et al.,2006),2011年在北京郊區發現了印加孔雀草野生群落(張勁林等,2014),并呈現種群入侵擴張趨勢,近幾年陸續在河北、山東、山西、西藏等多地發生大面積危害(張瑞海等,2019)。印加孔雀草對生境的適應能力強,耐干旱、耐鹽堿、耐瘠薄(Hulina,2008);具有強大的繁殖能力,種子小、產量高、且易傳播(張勁林等,2014);并分泌化感物質抑制其伴生植物的生長(Meissner et al.,2013)。目前國內對其研究較少,研究內容主要集中在形態學、發生危害等方面,張瑞海等(2019)分析證明了印加孔雀草為具有危害性和侵入性的物種,土艷麗等(2019)研究表明了惡性入侵植物印加孔雀草能顯著增加農作物藏青稞的死亡率;而國外對印加孔雀草的研究主要集中在其精油的化學成分及利用等方面(Martín et al.,2005)。
目前國內外對于印加孔雀草入侵對根際土壤微生物群落的影響研究尚未見報道。印加孔雀草入侵怎樣影響土壤細菌群落結構?而與其共生植物存在競爭時又是否會造成土壤微生物群落結構的變化?基于以上問題,本研究采用第二代高通量測序技術,以印加孔雀草為研究對象,探討其入侵及與本地植物競爭條件下土壤微生物群落的響應,比較分析印加孔雀草對土壤細菌群落多樣性的影響及其與環境因子間的關系,以期從微生態學的角度探究印加孔雀草的入侵機制,為其綜合防控及生態系統的修復提供理論依據。
1 材料與方法
1.1 試驗材料采集
試驗所需印加孔雀草種子、入侵發生地原位土壤皆采集于北京市昌平區興壽鎮桃林村(40°13′47.98″N、116°25′26.63″E),該地位于北京市西北部,屬暖溫帶,為半濕潤大陸性季風氣候,具有明顯的干季和雨季,四季分明,是印加孔雀草入侵較為嚴重的地區之一。所采集的原位土壤為適合印加孔雀草生存但未入侵過的褐土,萬壽菊種子購買于市面種子公司。
1.2 試驗設置及取樣
試驗區位于中國農業科學院農業環境與可持續發展研究所順義農業環境綜合試驗基地(北京市順義區 40°14′55.42″N,116°39′23.10″),設置 4 個處理(花盆大小為:23 cm×18.4 cm):未種植任何植物的裸土(不種植任何植物,記為CK)、本地植物萬壽菊單優群落(只種植萬壽菊,記為 Te)、入侵植物印加孔雀草與本地植物萬壽菊混種群落(印加孔雀草與萬壽菊1:1種植,記為TmTe)、入侵植物印加孔雀草單優群落(只種植印加孔雀草,記為Tm),每個處理3次重復。整個試驗持續50 d,覆蓋植物發芽、營養生長等過程,使其形成相對穩定的根際微環境,期間保證適當的水分供應。試驗結束后,將每個花盆輕輕去除上層土(3 cm左右),然后采用抖土法取得植物根際土壤(Hofman et al.,1989)。采集到的每個處理的土壤樣品充分混勻后,分別采用四分法取得一部分土壤樣品,裝進自封袋,立即帶回實驗室放入?80 ℃冰箱中,用于高通量測序;其他部分也分別裝進自封袋,帶回實驗室,置于陰涼處自然晾干,用于測土壤 pH值、全氮(TN)、全磷(TP)、全鉀(TK)和有機質(OM)等指標。而后隨機選取各處理中(CK除外)長勢一致具有代表性的5株植株(混種處理中每種植物各選取5株),將植株洗凈,用于植物生物量的測定。
1.3 植物生物量及土壤理化性質的測定
將植株洗凈,用濾紙吸去多余水分,120 ℃殺青、80 ℃烘干至恒質量,測定植物干質量。檢測前處理后的土壤樣品理化性質,包括土壤pH值(電極法),有機質(重鉻酸鉀容量法)、全氮(半微量開氏法)、全磷(碳酸氫鈉鉬藍法)以及全鉀(火焰光度法)含量,測定方法參照鮑士旦(2005)方法。
1.4 土壤DNA的提取及細菌16S rDNA的PCR擴增
采用土壤基因組 DNA提取試劑盒(北京,Solarbio公司)提取土壤樣品總基因組DNA,利用1%瓊脂糖凝膠電泳及超微量紫外分光光度法檢測DNA純度。以純化后的DNA為模板,以細菌16S rDNA V3-V4區通用引物(V3F:5'-TACGGRAGGCA GCAG-3′,V4R:5′-AGGGTATCTAATCCT-3′)進行PCR 擴增(Peiffer et al.,2013;Bortolini et al.,2016)。采用TransGen AP221-02 PCR反應體系:TransStart Fastpfu DNA Polymerase,20 μL,DNA 模板 10 ng,上、下游引物(5 μmol·L?1)各 0.4 μL,dNTP(2.5 mmol·L?1) 2 μL , FastPfu Polymerase 0.4 μL ,5×FastPfu buffer 4μL,補充 ddH2O 至 20 μL。PCR擴增程序為:95 ℃預變性2 min;95 ℃變性30 s、55 ℃退火30 s,72 ℃延伸45 s,30個循環;最后于72 ℃延伸10 min,4 ℃保存。每個樣品3個重復,將同一樣品的PCR產物混合后用質量體積比為2%的瓊脂糖凝膠進行電泳檢測。
1.5 Miseq文庫構建及測序
樣品質檢合格之后,構建測序文庫,其步驟分為:(1)連接“Y”字形接頭;(2)使用磁珠篩選去除接頭自連片段;(3)利用PCR擴增進行文庫模板的富集;(4)氫氧化鈉變性,產生單鏈DNA片段。利用Illumina公司的Miseq PE300平臺進行測序(由北京奧維森基因科技有限公司提供),步驟如下:(1)DNA片段的一端與引物堿基互補,固定在芯片上;(2)另一端隨機與附近的另外一個引物互補,也被固定住,形成“橋(bridge)”;(3)PCR擴增,產生DNA簇;(4)DNA擴增子線性化成為單鏈;(5)加入改造過的DNA聚合酶和帶有4種熒光標記的dNTP,每次循環只合成 1個堿基;(6)用激光掃描反應板表面,讀取每條模板序列第一輪反應所聚合上去的核苷酸種類;(7)將“熒光基團”和“終止基團”化學切割,恢復3′端粘性,繼續聚合第2個核苷酸;(8)統計每輪收集到的熒光信號結果,獲知模板DNA片段的序列。
1.6 數據分析
Miseq測序得到的PE reads首先根據overlap關系進行拼接,同時對序列質量進行質控和過濾。由于不同樣本間的 reads數不同,因此在分析多樣性和群落組成前,先依據最低 reads數進行標準化。區分樣品后進行OTU(operational taxonomic unit)聚類分析和物種分類學分析,基于 OTU進行物種多樣性指數分析。基于分類學信息,在各個分類水平上進行群落結構的統計分析。在上述分析的基礎上,進行一系列群落結構和系統發育等的統計學和可視化分析。使用FLASH軟件(FLASH v 1.2.7)對下機數據進行拼接,得到OTUs數據(Raw Tags),利用Trimmomatic(v 0.33)對Raw Tags質控過濾,得到Clean Tags,所得Clean Tags數據上傳至在美吉云平臺 I-Sanger(https://www.i-sanger.com/),進行數據分析。基于 97%相似度下,利用 Usearch(vsesion 7.0)軟件在Silva數據庫進行OTU物種分類學分析和聚類分析。4組樣本利用Adonis置換多因素方差分析和 ANOSIM 相似性分析,判斷分組的可信度和可行性。
Ace、Chao指數為土壤細菌群落豐富度指標,皆用以估計群落中OTU數目,Chao指數采用chao1算法(Chao,1984),Ace指數算法與其不同(Chao et al.,1993);Shannon、Simpson 指數為用來估算樣品中細菌群落多樣性指標,Shannon值越大群落多樣性越高(Shannon,1948a,1948b),Simpson指數則是由 Edward Hugh Simpson(1949)提出,Simpson值越大群落多樣性越低。利用 mothur(version v.1.30.1)對 Ace、Chao、Shannon 和 Simpson指數進行分析Alpha多樣性,用T檢驗分析Alpha多樣性指數差異,然后用R作圖;利用柱形圖表示細菌群落組成;各樣本差異物種組成通過One-way ANOVA單因素方差分析,LEfSe軟件首先使用nonparametric factorial Kruskal-Wallis (KW) sum-rank test(非參數因子克魯斯卡爾—沃利斯秩和驗檢)檢測具有顯著豐度差異特征,并找到與豐度有顯著性差異的類群,然后采用線性判別分析(LDA)來估算每個組分(物種)豐度對差異效果影響的大小;環境因子與菌群的關聯分析,檢測pH、OM、TN、TP、TK等5個指標,利用單因素方差分析及posthoc檢驗篩選不同樣點根際土壤菌群差異物種,通過計算環境因子與根際土壤菌群之間的相關性系數(Spearman等級相關系數、Pearson相關系數等),然后利用Pheatmap Package軟件,將獲得的數值矩陣通過Heatmap圖展示出平均相對豐度前10的屬與環境因子的相關性。
2 結果
2.1 植物生物量及土壤理化性質
各處理印加孔雀草與萬壽菊的生物量及土壤理化性質指標分析結果如表1所示。結果顯示,無論單種(Tm、Te)處理還是混種(TmTe)處理,入侵植物印加孔雀草的生物量都顯著高于萬壽菊。土壤理化性質結果顯示,相比于CK,3個處理(Tm、Te、TmTe)的土壤理化性質都發生了不同程度的變化。土壤pH均呈堿性,且有印加孔雀草入侵過的土壤pH顯著增加,各處理之間差異顯著(P<0.05);種植植物的3個處理中,TmTe的土壤全氮(TN)質量分數高于Tm(P=0.033)與Te(P=0.722),均值為0.116%;Tm的土壤全鉀(TK)質量分數顯著高于 TmTe(P=0.047)與 Te(P=0.028),為 1.473%;土壤全磷(TP)、有機質質量分數在各處理之間差異不顯著。
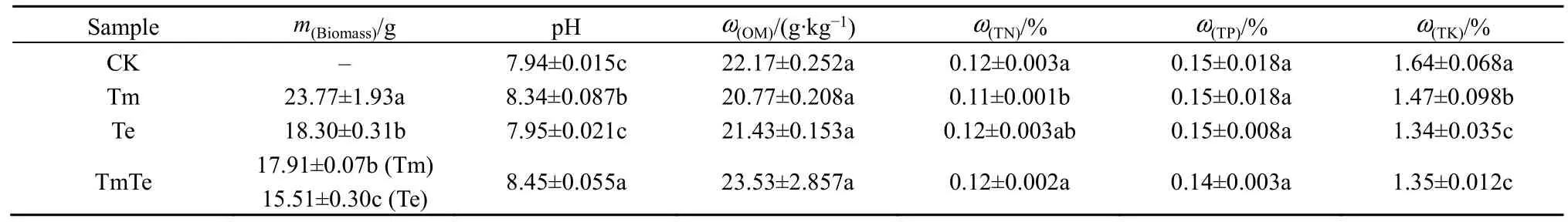
表1 植物生物量及土壤理化性質比較Table 1 Comparison of plant biomass, soil physical and chemical properties
2.2 土壤細菌的OTU豐度和α多樣性
對 12個樣品進行高通量測序,共產生 223.72 Mb干凈數據,經過拼接和過濾處理后,獲得 16S rDNA標簽序列,并根據97%的序列相似性劃為不同的OTU。OTUs豐度稀釋曲線(圖1)顯示,隨著測序數量的增加,稀釋曲線斜率逐漸降低,趨向平坦但未達平臺期,說明大多數樣本的測序數據量足夠,能夠反映樣品中的物種組成特征。
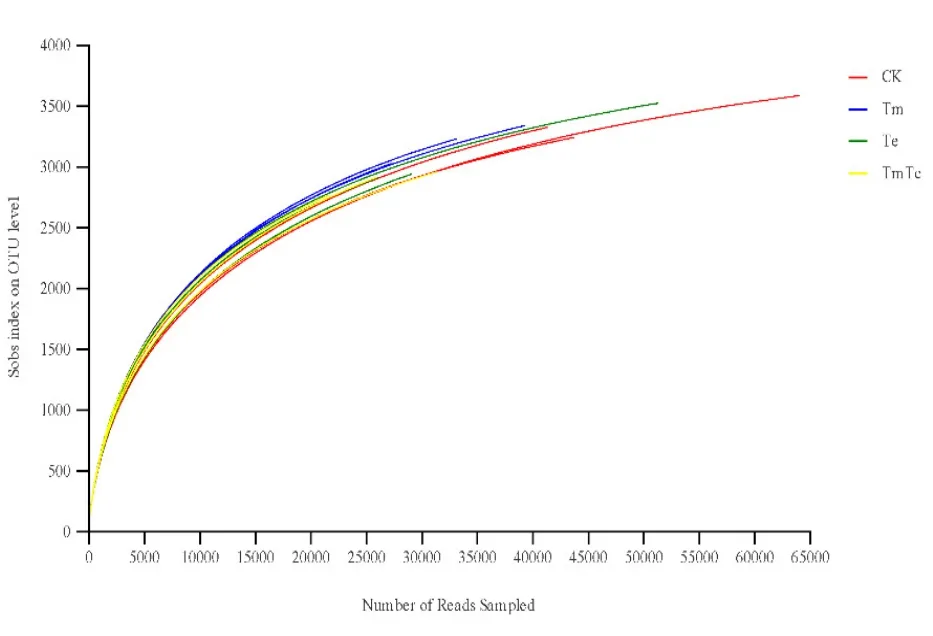
圖1 樣品在遺傳距離0.03下的稀釋曲線圖Fig. 1 The rarefaction of different samples obtained at the genetic distances of 0.03
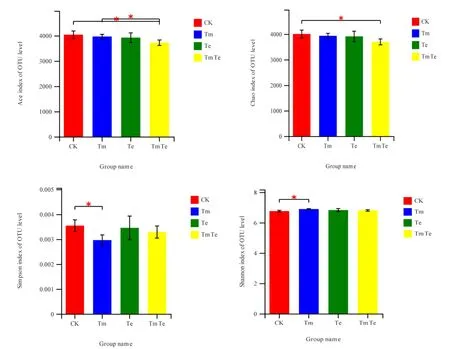
圖2 樣品在遺傳距離0.03下的細菌α多樣性指數Fig. 2 Alpha diversity for different samples obtained at genetic distance of 0.03
土壤細菌豐富度指數和多樣性指數如圖 2所示,CK、Tm、Te與TmTe的Simpson指數均值分別為0.0035、0.0030、0.0035、0.0033,Shannon指數均值分別為6.75、6.89、6.82、6.80,Tm與CK表現出了明顯差異,Simpson指數顯著低于 CK(P=0.033)、Shannon指數顯著高于CK(P=0.024),說明印加孔雀草單優群落的細菌群落多樣性最高,其它處理未表現出與CK的顯著性差異;CK、Tm、Te以及 TmTe的 Ace指數均值分別為 4035.9、3964.7、3918、3718.2,Chao指數的均值分別為4000.4、3928.6、3907.2、3691.7,TmTe的Ace指數顯著低于CK(P=0.039)與Tm(P=0.035)、Chao指數顯著低于CK(P=0.049),說明印加孔雀草與萬壽菊混種處理群落豐富度最低,印加孔雀草單優群落的豐富度高于混種處理,萬壽菊單優群落與其他處理未表現出顯著性差異。
2.3 高通量序列生物信息學分析
通過測序共產生4615個細菌OTU(圖3),其中 4個處理共有的 OTUs為 3169個,占總數的68.67%。CK的細菌群落數最高,平均含4189個細菌OTU;Tm次之,為4110個OTU;TmTe的細菌群落數最低,為3814個OTU。且TmTe和Te相比,TmTe特有的細菌OTU數目為321個,占8.42%。結果說明,不同處理帶來了不同程度的土壤細菌OTU差異,混種處理土壤細菌OTU數目最低,印加孔雀草單優群落處理有使OTU數目升高的趨勢,這與α多樣性中的群落豐富度的研究結果一致。
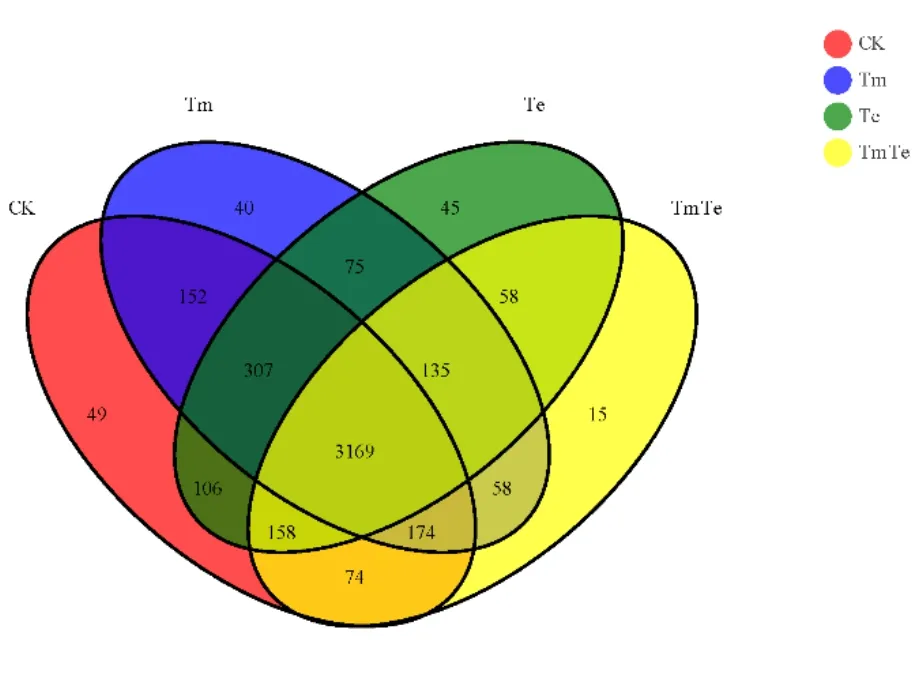
圖3 細菌OTUs分布韋恩圖Fig. 3 Venn graph of bacteria OTUs distribution
2.4 細菌群落組成
選取各樣品中細菌在門分類水平上最大豐度排名前 10的物種,生成物種相對豐度堆積圖(圖4)。由圖4可知,4個處理的土壤樣品都具有豐富的物種,其中豐度最高的 5個門為變形菌門(Proteobacteria)、酸桿菌門(Acidobacteria)、放線菌門(Actinobacteria)、綠彎菌門(Chloroflexi)及芽單胞菌門(Gemmatimonadetes),共計占各處理的80%以上。12個樣品中所檢測到的細菌門的數量相似,但不同處理不同門所占的比例不同。相比于裸土(CK),其他3個處理中不同門所占比例都有不同程度的變化。進一步分析3個處理(Te、TmTe、Tm)的群落組成平均值,發現隨著印加孔雀草密度的增加(Te、TmTe、Tm),放線菌門、綠彎菌門以及芽單胞菌門所占的比例呈現出先升高再降低的趨勢,變形菌門呈現先降低后升高的趨勢,而酸桿菌門呈現不斷上升的趨勢,但差異不顯著。這表明不同門下的細菌在印加孔雀草入侵過程中發揮著不同的功能,而酸桿菌門有可能在其入侵過程中起到比較重要的作用。
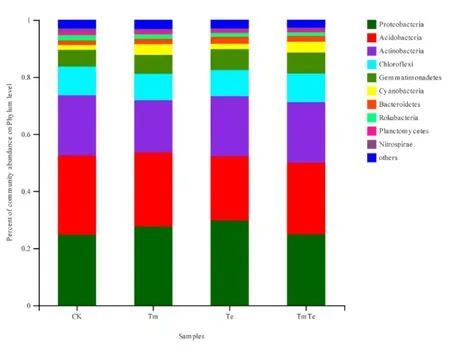
圖4 門水平上的細菌相對豐度柱形圖Fig. 4 The relative abundance of bacteria in Phylum level
2.5 Lefse多級物種差異判別分析
抽取出在門至屬水平上各處理序列數總和大于等于100個OTU的細菌,利用LEfSe軟件,可以得到Tm、Te、TmTe分別與CK相比顯著富集且具有顯著差異的菌,這些細菌與各自處理的植物群落的生長與維持關系密切,分別記為Tm vs CK、Te vs CK和 TmTe vs CK,對其進行韋恩圖分析(圖5)。得到對印加孔雀草入侵有利的細菌,為共同存在于Tm vs CK和TmTe vs CK中但不屬于Te vs CK的10個OTU類群。對其進行進一步分析(表2)顯示,這10個OTU類群隸屬于變形菌門、放線菌門、藍細菌門(Cyanobacteria)及酸桿菌門4個門,其數量分別為3、3、3、1個。其中,變形菌門下的有多囊粘細菌科(Polyangiaceae)、侏囊菌科(Nannocystaceae)等3個科下的3個菌屬;放線菌門下的有假諾卡氏菌科(Pseudonocardiaceae)、Ilumatobacteraceae科等2個科下的3個菌屬;藍細菌門下的有念珠藻科(Nostocaceae)、席藻科(Phormidiaceae)等2個科下的 3個菌屬以及酸桿菌門下Blastocatellaceae科下的 1個菌屬。另外,TmTe顯著性差異于其他處理的菌為圖中 29個OTU類群,Tm顯著性差異于其它處理的菌為圖中33個OTU類群。
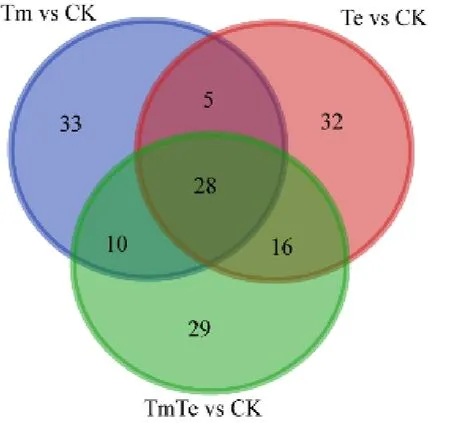
圖5 3個處理分別顯著富集且具有顯著差異于CK的細菌韋恩圖Fig. 5 Venn graph of bacteria with significant enrichment and difference between the three treatments and CK
2.6 環境因子關聯分析
選取各樣品中細菌在門分類水平上最大豐度排名前 10的物種,通過計算環境因子與菌群之間的 Spearman等級相關系數,生成環境因子相關性Heatmap圖(圖6)。由圖6可知,不同菌群與各種環境因子體現出不同程度的相關性關系。其中,硝化螺旋菌門(Nitrospirae)、酸桿菌門、棒狀桿菌門(Rokubacteria)、浮霉菌門(Planctomycetes)、放線菌門、變形菌門、擬桿菌門(Bacteroidetes)、綠彎菌門以及芽單胞菌門與土壤全磷、全鉀含量呈不同程度的正相關,與土壤pH呈現出了不同程度的負相關,與土壤全氮和土壤有機質含量無明顯相關性,個別呈現出了微弱的負相關關系。其中,放線菌門與土壤全鉀含量呈極顯著正相關(R=0.71735,P=0.00863),硝化螺旋菌門、變形菌門、擬桿菌門、綠彎菌門以及芽單胞菌門與土壤全鉀含量呈顯著正相關。但是,藍細菌門(Cyanobacteria)與環境因子之間的關系不同于其他菌門,具體表現與土壤全氮、全磷、全鉀含量呈微弱的負相關,與pH、有機質含量呈微弱的正相關。
3 結論與討論
3.1 討論
高通量序列生物信息學及α多樣性分析結果表明,印加孔雀草單優群落的土壤細菌群落多樣性最高且顯著高于CK,其它處理未表現出與CK的顯著性差異,這說明印加孔雀草的入侵可以提高土壤細菌群落多樣性,且入侵植物印加孔雀草對于土壤細菌多樣性的作用效果要大于與其共生的萬壽菊。宋振等(2016)在研究同為菊科的入侵植物黃頂菊的根際微生物群落結構時發現,黃頂菊單優群落的土壤細菌群落多樣性顯著高于本地植物,這與本研究的結果是一致的;入侵植物空心蓮子草(Altemanthera philoxeroides)可以通過改變土壤微生物菌群的結構和豐富度從而影響土壤有機碳的形成,其入侵提高了土壤微生物群落的多樣性(李珂,2019),印加孔雀草提高土壤細菌群落多樣性從而形成有利于自身生長的微生態環境,進而提高生物量以促進自身生長及發育,這很有可能是其入侵成功的重要原因之一。有研究表明,紫莖澤蘭與本地植物存在競爭時,其土壤細菌的群落豐富度與未入侵土壤及紫莖澤蘭單優群落土壤之間未表現出明顯差異(朱珣之等,2015);互花米草(Spartina alterniflora)入侵改變了土壤微生物群落和提高了根際微生物豐富度,而高豐富度的根際微生物影響了土壤養分循環和植物間的競爭關系,從而促進互花米草快速生長,并成功入侵(章振亞等,2012)。在本研究中,印加孔雀草與萬壽菊混種處理的群落豐富度最低且顯著低于 CK,印加孔雀草、萬壽菊單種處理都未表現出與CK的顯著性差異,說明印加孔雀草與萬壽菊種間競爭使得土壤細菌群落豐富度降低,這可能是萬壽菊與印加孔雀草相互作用的結果。

表2 在印加孔雀草入侵過程中起重要作用的10種菌在各水平上的分類及數目Table 2 Classification and the number in 10 bacteria that play an important role in invasion process of T. minuta L.
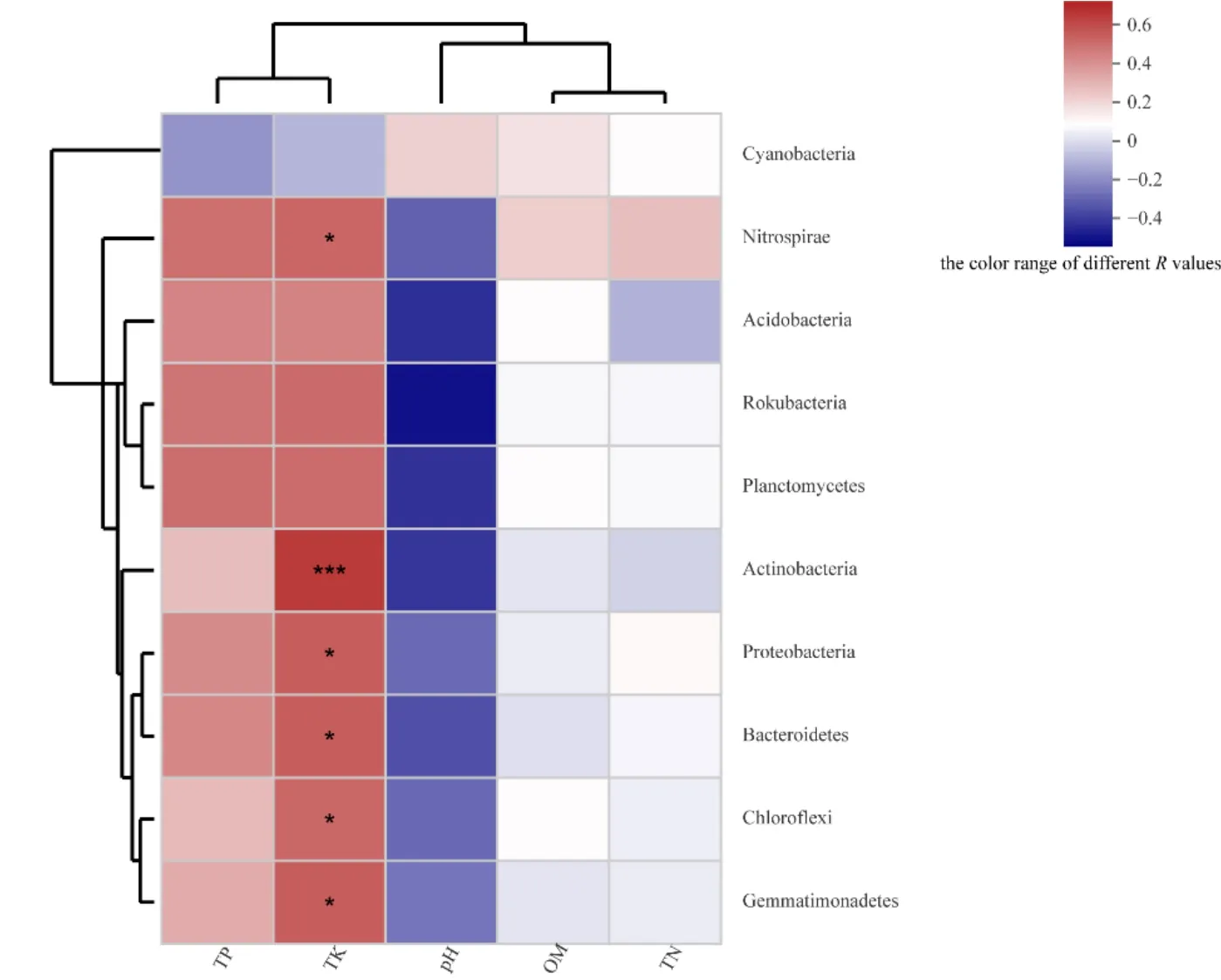
圖6 門分類水平上最大豐度排名前10的細菌與環境因子的相關性Heatmap圖Fig. 6 Heatmap map of the correlation between bacteria and environmental factors in the top 10 most abundant species on Phylu m level
本研究各處理中豐度最高的5個門依次為變形菌門、酸桿菌門、放線菌門、綠彎菌門和芽單胞菌門。通過 Lefse多級物種差異判別以及進一步的Venn分析,得到共同存在于Tm vs CK和TmTe vs CK中但不屬于Te vs CK的類群,為4個門(變形菌門、藍細菌門等)下的8個科(多囊粘細菌科、侏囊菌科、念珠藻科等)的共10個OTU類群,這些類群可能在印加孔雀草在入侵過程中起著重要作用。例如,多囊粘細菌科與侏囊菌科屬于粘細菌,隸屬于變形菌綱的粘球菌目。粘細菌是一類捕食菌,也是繼放線菌和真菌之后又一重要的活性次級代謝產物產生菌(王春玲等,2019),廣泛分布于全球各地的土壤中,能夠形成對溫度、干旱、輻射、鹽脅迫均具有抗性的粘孢子,而具有較高的環境適應力(Shimkets,1991),其群落和代謝產物對土壤微生態環境的改變很可能有利于印加孔雀草的入侵。而與TmTe顯著差異的存在于Tm、Te的29個OTU類群可能有利于維持印加孔雀草入侵的土壤微生態系統結構,Tm顯著性差異于 Te、TmTe的33個OTU類群則可能有利于印加孔雀草單優群落狀態的維持,但是否是這些 OTU類群及其功能還需要進一步深入研究。
不同門類的細菌同時受到土壤理化性質和植被特性等環境因子的影響而產生差異(Zhou et al.,2017),本研究中環境因子關聯分析結果表明,菌群與環境因子存在不同程度的顯著相關性,印加孔雀草入侵后,其根際土壤可能會快速形成穩定的細菌群落結構以及多樣性,有利于提高印加孔雀草其自身對環境的適應能力以及單優群落的形成。環境因子關聯分析也表明,不同門下的菌群與各種環境因子體現出不同的相關性特征,放線菌門與土壤全鉀含量呈極顯著正相關,硝化螺旋菌門、變形菌門、擬桿菌門、綠彎菌門以及芽單胞菌門與土壤全鉀含量呈顯著正相關,而藍細菌門與環境因子的關系則表現出了和其他菌門較為不同的特點。藍細菌是發現的最早能夠利用光合作用釋放氧氣的原核生物(Bryant,1995),在自然界中不僅分布在淡水、海水等水體中,也廣泛分布在各類土壤以及極端環境中(Nicholas et al.,1992)。本研究中藍細菌表現出與環境因子特殊的相關性可能是由于印加孔雀草在快速繁殖擴散過程中對土壤有機物合成和氧氣釋放有著較為特殊的需求。
植物根系分泌的很多次生代謝物質可作為植物與土壤微生物互作的信號分子物質而影響植物根際土壤微生物群落結構(Ueda et al.,2010;Ahuja et al.,2012),如豚草的次生代謝產物對土壤微生物具有一定的選擇抑制作用(趙昕,2003),黃頂菊次生代謝產物紫云英苷可以改變土壤微生物群落結構(張瑞海,2017)等。目前已有研究發現,印加孔雀草植株能夠分泌萜烯類化感物質從而抑制伴生植物的生長(Liza et al.,2008;Scrivanti et al.,2003),是否這些物質也導致了印加孔雀草土壤微生物的改變從而有利于自身群落的穩定,或者有其他特殊的次生代謝產物影響了土壤微生物群落,需要進一步深入研究。另外,本研究采用了入侵地未生長過印加孔雀草的單一褐土類型土壤,然而不同土壤類型的起始土壤細菌群落的組成存在差異,這種差異很可能會影響入侵種對土壤的響應,印加孔雀草對不同土壤類型的響應是否與本試驗結果存在不同,這還需要大量重復試驗進行研究。
3.2 結論
印加孔雀草入侵能夠改變入侵地土壤細菌多樣性。印加孔雀草顯著提高了土壤細菌群落多樣性,其與萬壽菊混種使得土壤細菌群落豐富度顯著降低,且印加孔雀草競爭效果大于非入侵植物萬壽菊;土壤中細菌相對豐度最高的前5個門分別是變形菌門、酸桿菌門、放線菌門、綠彎菌門和芽單胞菌門;不同菌群與環境因子表現出了不同程度的正負相關關系,其中放線菌門、硝化螺旋菌門、變形菌門、擬桿菌門、綠彎菌門以及芽單胞菌門與土壤全鉀含量呈顯著正相關,藍細菌門和環境因子的相關關系不同于其他菌門;多囊粘細菌科、侏囊菌科、念珠藻科等8個科的10個OTU類群有可能在印加孔雀草入侵過程中起著重要作用。本研究結果為揭示印加孔雀草入侵的土壤微生態機制提供了理論依據。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
