云南省熱帶、亞熱帶和亞高山森林凋落物層中彈尾類群落組成的研究
2020-08-03 06:06:40徐藝逸
綠色科技 2020年12期
關鍵詞:環境
徐藝逸
(1.中國科學院西雙版納熱帶植物園熱帶森林生態學重點實驗室,云南 勐臘 666303;2.中國科學院大學,北京 100049)
1 引言
土壤動物作為重要分解者,在生態系統的物質循環與營養供應中起著重要的作用。一方面土壤動物積極同化各種有用物質以建造其自身,另一方面又將其排泄產物歸還到環境中不斷改造環境。在土壤食物網中,土壤動物主要通過取食作用影響其他生物的數量以及群落結構。土壤食物網在有機質分解、碳存貯、土壤結構調節、病蟲害控制以及礦物質養分循環中起著重要作用[1]。彈尾類在土壤中十分常見,與螨類和線蟲同屬于3種最常見的優勢土壤動物類群。彈尾類對環境的變化十分敏感,土壤溫度、土壤pH值和土壤有機質含量等都可對其群落組成產生影響,所以彈尾類也可作為環境指示生物[2~5]。
關于土壤動物群落組成的研究多關注不同海拔的區別,其結果主要分為兩類:①對烏魯木齊雅瑪里克山(溫帶)土壤動物群落的研究表明:隨海拔上升,土壤動物數量、多樣性呈現遞減趨勢,不同海拔土壤動物群落間共有度較高,相似性差異不大,也有研究顯示熱帶彈尾類多度和亞熱帶東部山地大、中小型土壤動物(土壤彈尾蟲與土壤螨類)多度也有相同的結果[6~8];②衡山森林生態系統(亞熱帶)土壤動物密度最大出現在中間海拔段,海拔的升高或者降低土壤動物相應減少,在大興安嶺森林(寒溫帶)中小型土壤動物種類、數量和黃山(亞熱帶)土壤彈尾類的數量均有相似結果[9~11]。
雖然關于彈尾類群落組成在不同海拔的研究較多,但很少有人進行跨氣候帶與海拔的交叉研究。云南省分布著熱帶(西雙版納補蚌)、亞熱帶(哀牢山鎮沅千家寨地區)和亞高山(麗江玉龍雪山)3種典型氣候帶,它們處于同一經度(E 101°)的不同緯度上,這很好的排除了因經度差異造成的一系列影響。因氣候帶的不同,導致了土壤溫度、土壤pH值和土壤有機質含量等差異將影響彈尾類的群落組成情況。本研究意在回答如下問題:不同氣候帶彈尾類的群落組成情況;影響不同氣候帶彈尾類群落組成的環境因子是否相同。
2 研究地區與方法
2.1 樣地概況與設置
本研究樣地設置于西雙版納補蚌(熱帶)、哀牢山鎮沅千家寨(亞熱帶)和麗江玉龍雪山(亞高山帶)。
西雙版納傣族自治州位于云南省西南部(21°08′~22°36′N,99°56′~101°50′E)。海拔范圍從瀾滄江河谷的最低點491 m至最高峰2429 m。年平均氣溫為21.8 ℃,年平均降雨量為1493 mm[12],每年5月到10月為雨季,11月到次年4月為旱季。海拔1000 m以下的河谷地區為磚紅壤。主要的森林類型為熱帶季節雨林、熱帶季雨林和熱帶山地常綠闊葉林等。原始林的林冠高達50~60 m,由望天樹(Parashoreachinensis)占優勢,中下層喬木主要有毛猴歡喜(Sloaneatomentosa)、絨毛番龍眼(Pometiapinnata)、假海桐(Pittosporopsiskerrii)、云樹(Garciniacowa)、蟻花(Mezzettiopsiscreaghii)、染木(Saprosmaternata)等,次生林主要由殼斗科(Fagaceae)植物組成[13]。
哀牢山國家級自然保護區(鎮沅片區)位于云南省中部哀牢山地區(24°7′N,101°14′E),海拔2147 m,相對高度在2700 m以上。呈現雨季(5~10月)和旱季(11月至翌年4月)的交替。該地年平均降雨量1931.1 mm,相對濕度86%。年平均氣溫11 ℃,最冷月平均氣溫(1月)4.8 ℃,最熱月平均氣溫(7月)16.5 ℃,低溫極值-8.0 ℃,高溫極值24 ℃[14,15]。海拔2000~2700 m的植被類型主要為中山濕性常綠闊葉林,優勢植物包括殼斗科(Fagaceae)、木蘭科(Magnoliaceae)、樟科(Lauraceae)、山茶科(Theaceae)等[16]。
玉龍雪山位于滇西北麗江縣境內(27°10′~27°40′N,100°10′~100°20′E)。最高峰扇子陡海拔5596 m,相對高差3846 m。該區域年降水量935 mm,6~10月為雨季,這5個月的降水量為年降雨量的90%以上,11月至翌年5月為干季。年平均溫度12.79 ℃,最熱月為7月,月均溫17.9 ℃,最冷月是1月,月均溫5.9 ℃[17]。森林由下而上分別為云南松林、麗江云杉林、大果紅杉林和冷杉林[18,19]。從低海拔至高海拔,土壤類型依次為棕壤(2800~3200 m)、棕色針葉林土(3200~3600 m)、高山草甸土(3600~4200 m)[20]。
樣地設置:熱帶、亞熱帶和亞高山氣候帶間高差為600 m,在每個氣候帶內設置4個海拔樣帶,海拔樣帶間的垂直高差200 m,其中,熱帶海拔梯度為800 m、1000 m、1200 m、1400 m,亞熱帶海拔梯度設置為2000 m、2200 m、2400 m、2600 m,亞高山海拔梯度設置為3200 m、3400 m、3600 m、3800 m。每個海拔帶上設置4個20 m × 20 m的樣地。
2.2 土壤動物采集與鑒定
在樣地內隨機選擇3個0.6 m × 0.6 m的小樣方抓取其中的全部凋落物于布袋中,做好標簽,三處合并為一個混合樣。將野外采集的凋落物樣品置于烘蟲裝置中,利用土壤動物懼怕高溫和光亮的特性將其烘烤出來,落入酒精中即得到土壤動物的酒精浸泡樣本。依據《中國土壤動物檢索圖鑒》《中國亞熱帶土壤動物》《中國土壤動物》等資料進行彈尾類的分類鑒定(屬)并計數[21~23]。
2.3 環境數據采集
土壤溫度:2017年5~10月在樣地內布置的土壤溫度自動記錄設備(iButton紐扣式溫度計,每個小時自動記錄1個土壤溫度數據,10 cm深),獲得每個樣地的雨季土壤平均溫度(T-rain)、雨季土壤極端高溫(T-maxR)與雨季土壤極端低溫(T-minR)。
土壤水分:在取土壤動物的同時,隨機選擇3個小樣方,分別用環刀取100 cm3的土壤放入鋁盒,使用烘箱烘干法(105 ℃)測定土壤含水量(soil moisture)。
土壤pH值:使用pH測量儀測定。儀器型號為:雷磁PHS-25數顯型pH計。
土壤容重(Bulk density):容重法[24]。
土壤密度:密度計法。儀器型號為:TM-85密度計。
土壤孔隙度(Soil porosity):通過容重和比重計算獲得[24]:

凋落物厚度(Litterfall amount):用鋼尺直接測量,調查記錄野外獲取凋落物層土壤動物的3個樣方的凋落物平均厚度。
2.4 數據處理
考慮到生物群落組成多為非線性,對不同海拔梯度上的彈尾類群落組成的間接排序采用NMDS方法,直接排序采用CCA方法。CCA分析采用前置逐步回歸法篩選具有顯著相關的環境因子,通過模擬999次檢驗排序軸的顯著性。
所有分析在R軟件(Version 3.3.2)中進行,主要用到的R包有“vegan”,“ade4”,“packfor”等。
3 結果與分析
3.1 不同氣候帶彈尾類組成概況
熱帶共采集彈尾類2431只,隸屬于10科27屬。優勢屬有4個,其中球角蟲兆屬(22.1%)數量最多,其余優勢屬為近缺蟲兆屬(21.0%,等節蟲兆科)、刺齒蟲兆屬(15.9%,長角蟲兆科)和擬缺蟲兆屬(14.2%,等節蟲兆科),這4個屬合計占熱帶彈尾類總體捕獲量的73.2%;亞熱帶共采集彈尾類2315只,隸屬于8科21屬。優勢屬有2個,其中符蟲兆屬為絕對優勢屬(40.1%),另一個優勢屬為近缺蟲兆屬(13.1%),共占亞熱帶彈尾類總體捕獲量的53.2%;亞高山共采集彈尾類14404只,優勢屬有兩個,其中絕對優勢屬為符蟲兆屬(46.5%),另一優勢屬為棘蟲兆屬(28.3%,棘蟲兆科),這兩屬占亞高山彈尾類總體捕獲量的74.8%。
3.2 彈尾類群落的間接排序(NMDS)分析
NMDS分析(圖1)表明,在3個氣候帶上,彈尾類的物種組成在NMDS1軸能較好的區分,熱帶和亞高山氣候帶的物種組成有明顯差異,亞熱帶作為過渡區域與熱帶和亞高山物種組成均有部分重疊。在氣候帶內部不同海拔段彈尾類的物種組成差異不明顯。
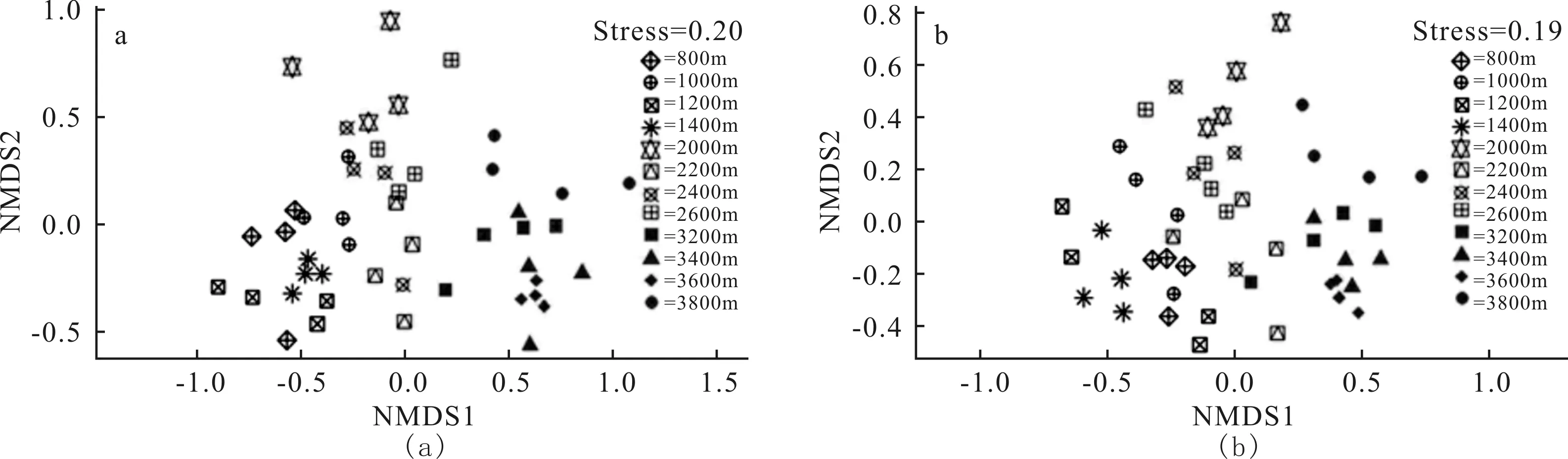
圖1 彈尾類物種組成在不同氣候帶的差異,基于Jaccard相似性指數(a)和Bray-Curtis相似性指數(b)的非參數排序(NMDS)結果
3.3 彈尾類群落的直接排序(CCA)分析
對全部海拔段的彈尾類群落組成與環境因子進行CCA分析(圖2)發現,CCA1軸和CCA2軸共同解釋了33.91%的物種信息量,顯著相關的環境因子為雨季土壤平均溫度、雨季土壤極高溫度、土壤pH值和凋落物厚度。凋落物凋落物厚度和土壤pH值與CCA1軸呈正相關,而雨季土壤最高溫和雨季土壤平均溫度和CCA1軸呈負相關。CCA1軸的正負軸較好的分割了高海拔和低海拔區域,高海拔(≥3200 m)大致分布在CCA1正軸,低海拔(≤2200 m)分布于負軸。凋落物厚度明顯與CCA2軸呈正相關,土壤pH值則呈負相關。在海拔1000 m、1200 m、1400 m、2000 m處彈尾類群落組成主要受雨季土壤極高溫度和雨季土壤平均溫度影響。在海拔3600 m、3800 m處彈尾類群落組成主要受高土壤酸堿度和凋落物厚度影響。
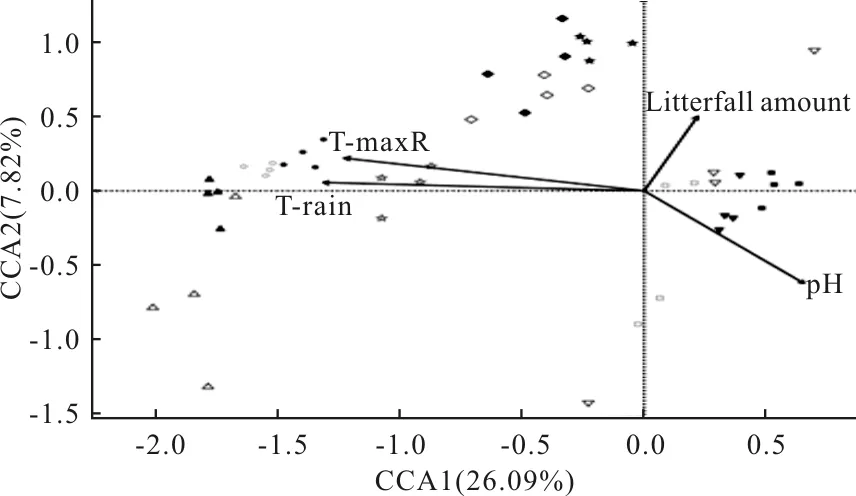
圖2 整體海拔彈尾類群落組成與環境因子的CCA排序
對熱帶地區不同海拔的群落組成與環境因子進行CCA分析(圖3)發現,CCA1軸和CCA2軸共同解釋了27.97%的物種信息量。顯著相關的環境因子為土壤pH值、凋落物厚度和雨季土壤極低溫度。土壤pH值和雨季土壤極低溫度與CCA1軸呈正相關,而凋落物厚度和CCA1軸呈負相關。CCA1軸的正負軸較好的分割了高海拔和低海拔區域,低海拔(800 m)大致分布在CCA1正軸,高海拔(1000 m、1200 m、1400 m)分布于負軸。凋落物厚度明顯與CCA2軸呈正相關。在海拔1000 m處彈尾類群落組成主要受凋落物厚度影響。在海拔3800 m處彈尾類群落組成主要受土壤酸堿度影響。
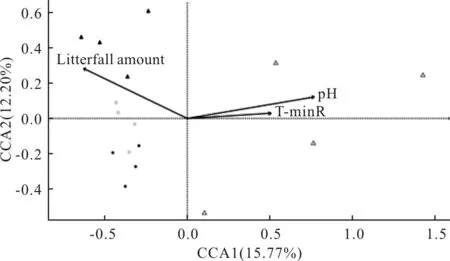
圖3 熱帶不同海拔彈尾類群落組成與環境因子的CCA排序
對亞熱帶地區不同海拔的群落組成與環境因子進行CCA分析發現(圖4),CCA1軸和CCA2軸共同解釋了33.28%的物種信息量。顯著相關的環境因子為雨季土壤平均溫度和雨季土壤極高溫度。雨季土壤極高溫度與CCA1軸呈正相關,而雨季土壤平均溫度和CCA1軸呈負相關。雨季土壤極高溫度和雨季土壤平均溫度與CCA2軸呈正相關。在海拔2200 m處彈尾類群落組成主要受雨季土壤極高溫度和雨季土壤平均溫度影響。
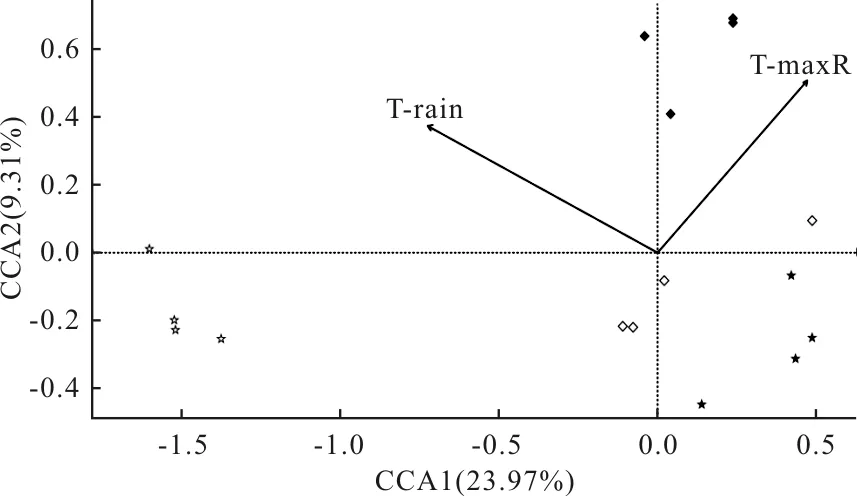
圖4 亞熱帶不同海拔彈尾類群落組成與環境因子的CCA排序
對亞高山氣候地區不同海拔的群落組成與環境因子進行CCA分析發現(圖5),CCA1軸和CCA2軸共同解釋了27.97%的物種信息量。顯著相關的環境因子為凋落物的量、土壤孔隙度、土壤pH值。土壤pH值與CCA1軸呈正相關,而凋落物厚度和CCA1軸呈負相關。凋落物厚度、土壤孔隙度和土壤pH值明顯與CCA2軸呈負相關。在海拔3800 m處彈尾類群落組成主要受凋落物厚度、土壤孔隙度影響。
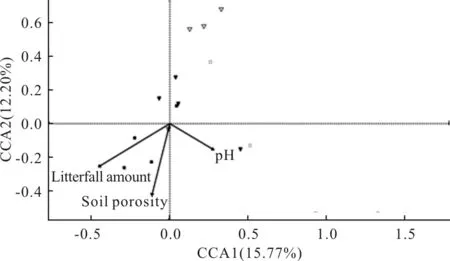
圖5 亞高山不同海拔彈尾類群落組成與環境因子的CCA排序
4 討論
從NMDS分析圖可以看出,不同氣候帶彈尾類的群落組成有所不同,熱帶和亞高山的物種組成有明顯差異,亞熱帶作為過渡區域與熱帶和亞高山物種組成均有部分重疊。這可能是由于亞熱帶本就屬于熱帶和亞熱帶的過度氣候類型,氣候特點處于二者之間,彈尾類受環境的影響,其群落組成與其他兩個氣候帶均有相似部分。在氣候帶內部不同海拔段彈尾類的物種組成差異不明顯,這可能是由于取樣處于雨季末期,溫度和土壤含水量等條件都較適宜彈尾類的生長,環境脅迫較少。
由整體彈尾類群落的直接排序(CCA)分析可以看出,亞高山彈尾類的群落組成的主要相關環境因子為土壤pH值和凋落物的厚度,可能由于亞高山氣溫常年低于其他兩個氣候帶,而本研究取樣于雨季,限制亞高山地區的彈尾類生長的溫度脅迫消失,就體現出土壤pH值和凋落物的厚度(土壤有機質)對彈尾類的群落組成的影響,這與前人發現的對彈尾類群落組成的影響因素一致[3~5];熱帶彈尾類的群落組成的主要相關環境因子為土壤平均溫度和土壤極高溫度,可能是由于熱帶在3個氣候帶中年均溫最高,而雨季的土壤溫度在全年中又為最高,雨季平均溫度和雨季極高溫可能成為彈尾類的限制因子,之前的研究也發現了彈尾類的群落組成與溫度密切相關[2,7];亞熱帶彈尾類的群落組成與本研究所涉及的環境因子相關性較小,可能由于其屬于其他兩個氣候帶的過渡氣候類型,彈尾類的群落組成受到較復雜的環境因子影響,也可能本研究沒有涉及到與其群落組成相關性較高的環境因子,有待進一步研究。分氣候帶的彈尾類群落的直接排序(CCA)分析可以看出,熱帶彈尾類群落組成主要與土壤溫度、土壤pH值和凋落物厚度相關;在亞熱帶,與彈尾類群落組成相關的主要環境因子為土壤溫度;而在亞高山,與彈尾類群落組成相關的環境因子主要是凋落物厚度、土壤孔隙度和土壤pH值。在氣候帶內部,與彈尾類群落組成的相關環境因子與整體排序的環境因子不同,可能是由于彈尾類生活在凋落物中,群落組成受小生境的影響更大。
5 結論
熱帶與亞高山彈尾類群落組成相差較大,亞熱帶作為過渡氣候帶,其群落組成與熱帶和亞高山均有相似部分;在氣候帶內部彈尾類群落組成相似度較高。從整體上看,土壤溫度、凋落物厚度和土壤pH值是與彈尾類群落組成相關的主要環境因子;而不同氣候帶與彈尾類群落組成相關的環境因子有所不同,熱帶彈尾類群落組成主要與土壤溫度、土壤pH值和凋落物厚度相關;在亞熱帶,與彈尾類群落組成相關的主要環境因子為土壤溫度;而在亞高山,與彈尾類群落組成相關的環境因子主要是凋落物厚度、土壤孔隙度和土壤pH值。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
