多菌種分步發酵提高辣木籽粕蛋白含量及酶活
2020-08-02 10:06:04郭志鵬夏學森黃文琪吳振強
中國釀造 2020年6期
郭志鵬,夏學森,黃文琪,2,吳振強
(1.華南理工大學 生物科學與工程學院,廣東 廣州 510006;2.江門市泛亞生物工程與健康研究院,廣東 江門 529000)
蛋白飼料是水產、禽畜養殖的重要資源,目前,世界各國對飼料蛋白的需求量都很大,而魚粉等傳統蛋白飼料價格正在提高,尋找潛在的、優質的蛋白飼料是目前養殖業的重要目標之一。因此,優化飼料配方、降低飼料成本是養殖業重要的研究問題[1-2]。
辣木是一種尚未充分開發的多功能新型材料,具有產量高、適應性廣、栽培簡便、抗逆性強等特點。辣木籽在生產辣木油后,剩余的辣木籽粕如果未被充分利用就被丟棄,會造成環境污染和巨大的浪費。辣木籽粕本身有較高的粗蛋白含量(25%~36%),但植物原料常常含有抗營養因子,直接作為飼料難以被消化,甚至可能導致幼崽死亡。許多微生物具有豐富的酶系,可以降解、轉化糟粕類廢渣,廢渣經過益生菌發酵處理后,能夠顯著增加其中的有益活性代謝產物[3-8],如維生素、氨基酸、小肽等營養物質的含量,降解抗營養因子[9-10],提高發酵飼料的營養價值和口感風味[11-13],延長飼料的貯存期,通過飼喂發酵飼料,可改善動物腸道微生物生態,維持腸道pH穩定,同時降低生產成本,是理想的開發辣木籽粕的方法。辣木籽作為飼料可提高肉雞的肉脂、能量、多不飽和脂肪酸[14-15],直接利用辣木葉[16-17]、辣木籽作為飼料或者將辣木葉用于制作青貯飼料的報道較多,但開發利用辣木籽粕固態發酵生產蛋白飼料的報道較少。
利用屎腸球菌、黑曲霉和米根霉對辣木籽粕進行固態發酵,通過單因素試驗和正交試驗設計對其接種順序、發酵培養基含水量、接種量和發酵時間進行發酵條件的優化,確定了開發利用辣木籽粕生產發酵蛋白飼料的最佳工藝,以期為開發利用辣木籽粕提供依據。
1 材料與方法
1.1 材料與試劑
辣木籽粕(使用前經過粉碎,粗蛋白含量35.38%,酸溶蛋白含量5.10%):由天源利生物科技(廣州)有限公司提供。
屎腸球菌(Enterococcus faecium)A1(美國國家生物信息中心(national center of biotechnology information,NCBI)登錄號MN474019),米根霉(Rhizopus oryzae)E20360(NCBI登錄號MK267423),黑曲霉(Aspergillus niger)B1(NCBI登錄號MN474007):均為前期試驗分離得到。
MRS培養基(制作平板時添加2%瓊脂),馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)培養基(制作平板時添加2%瓊脂):廣東環凱微生物科技有限公司;固態發酵培養基(辣木籽粕,根據試驗條件,加入蒸餾水調整含水量,121 ℃滅菌20 min)。
五水硫酸銅、氫氧化鈉、鹽酸(均為分析純):廣州化學試劑廠;硫酸鉀、硼酸(均為分析純):致遠化工天津有限公司;三氯乙酸(分析純):上海凌峰化學試劑有限公司。
1.2 儀器與設備
Autoclave SP510高溫滅菌鍋:日本Yamato科技有限公司;SW-CK-1F超凈工作臺:蘇州安泰空氣技術有限公司;LRH-150B培養箱:韶關市泰宏醫療器械有限公司;KDN-1自動凱氏定氮儀:上海儀電科學儀器股份有限公司。
1.3 試驗方法
1.3.1 菌種活化
在無菌操作下,挑取米根霉、黑曲霉一環,分別接種到PDA培養基上;挑取屎腸球菌一環接種到MRS培養基上。30 ℃培養48 h。
1.3.2 菌種擴大培養
米根霉、黑曲霉擴大培養:配制PDA液體培養基,分裝于250mL帶擋板的三角瓶(每瓶50 mL培養基),滅菌后分別接種活化后的米根霉、黑曲霉,30 ℃分別培養12 h、24 h;屎腸球菌擴大培養:配制MRS培養基,分裝于250 mL三角瓶(每瓶50 mL培養基),滅菌后接種活化后的屎腸球菌,37 ℃培養14 h。根據前期試驗結果,米根霉、黑曲霉和屎腸球菌分別培養12 h、24 h和14 h后,菌種到達對數生長期,有利于適應固態發酵培養基環境。
1.3.3 培養條件對辣木籽粕發酵轉化的影響
基本發酵條件:稱取一定量辣木籽粕于250 mL錐形瓶中,加入蒸餾水調整含水量,使發酵培養基總質量為40 g。121 ℃滅菌20 min,冷卻至室溫,在無菌條件下接種種子液(各種子液接種比例為1∶1∶1),30 ℃培養。發酵結束后,45 ℃烘干,磨粉,測定粗蛋白和酸溶蛋白含量、酸性蛋白酶和中性蛋白酶活性。
在上述發酵條件下進行單因素試驗,研究接種順序(①先接種屎腸球菌,發酵一半時間后接種米根霉和黑曲霉;②先接種米根霉和黑曲霉,發酵一半時間后接種屎腸球菌;③同時接種屎腸球菌、米根霉和黑曲霉)、培養基含水量(20%、30%、40%、50%、60%)、接種量(10%、15%、20%、25%、30%)和發酵時間(1 d、2 d、3 d、4 d、5 d)對辣木籽粕發酵后粗蛋白和酸溶蛋白含量、酸性蛋白酶和中性蛋白酶活性的影響。各組試驗重復3次。
1.3.4 工藝條件優化正交試驗
根據單因素試驗的結果,接種順序、含水量、接種量和發酵時間以表1水平進行L9(34)的正交試驗。

表1 辣木籽粕發酵條件優化正交試驗因素與水平Table 1 Factors and levels of orthogonal experiments for fermentation conditions optimization of Moringa oleifera seed meal
1.3.5 測定方法
粗蛋白的測定:改良的凱氏定氮法[18],在消解步驟中加入過氧化氫加快消解過程。
酸溶蛋白的測定[19]:稱取1.00 g樣品加入燒杯,加入50 mL 15%三氯乙酸,超聲振蕩提取15 min。提取液過濾后,取10 mL提取液用上述改良的凱氏定氮法測定樣品酸溶蛋白含量。
蛋白酶活性測定[20]:測定酸性蛋白酶和中性蛋白酶活性使用了福林酚法。在一定溫度(40±0.2)℃和相應的pH條件下(酸性蛋白酶為pH 3.0,中性蛋白酶為pH 7.2),定義在1 min內水解酪蛋白產生相當于1 μg酚基氨基酸(由酪氨酸等同物表示)的酶量為1個酶活單位,以U表示。
1.3.6 統計學分析方法
各指標采用了SPSS 22.0軟件對數據進行了極差分析和方差分析。
2 結果與分析
2.1 接種順序對辣木籽粕生物轉化的影響
由圖1可知,接種順序對酸溶蛋白、酸性蛋白酶和中性蛋白酶有顯著影響(P<0.05),對粗蛋白含量影響不顯著(P>0.05)。先接種霉菌時,各指標均達到最高值,粗蛋白和酸溶蛋白分別達到44.39%和8.25%,酸性蛋白酶與中性蛋白酶活性分別為103.12 U/g和528.44 U/g。米根霉、黑曲霉具有豐富的酶系,可將原料的大分子化合物降解為易消化的小分子化合物,也可以利用分解得到的小分子化合物合成新的生物大分子,有利于提高材料的營養價值,也為后續接種屎腸球菌提供了營養豐富的環境。接種順序對粗蛋白外的3個指標均有顯著影響,因此選擇先接種黑曲霉和米根霉,發酵一半時間后,再接種屎腸球菌進行后續試驗。
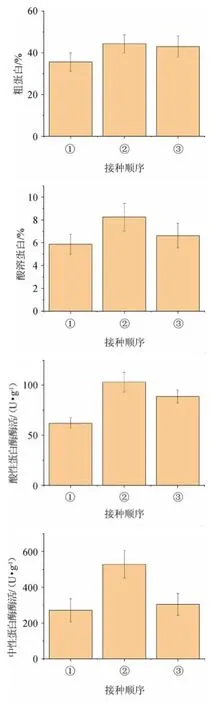
圖1 接種順序對辣木籽粕生物轉化的影響Fig.1 Effect of inoculation order on biotransformation of Moringa oleifera seed meal
2.2 培養基含水量對辣木籽粕生物轉化的影響
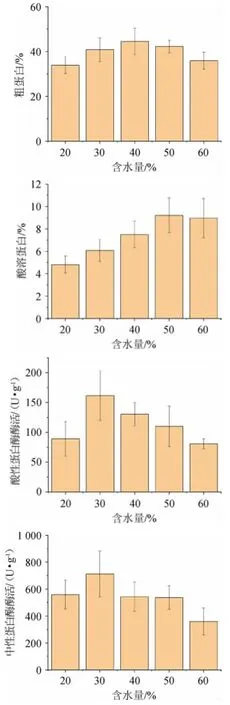
圖2 含水量對辣木籽粕生物轉化的影響Fig.2 Effect of water content on biotransformation of Moringa oleifera seed meal
由圖2可知,培養基含水量對4個指標均有顯著影響(P<0.05)。當含水量為40%時,粗蛋白含量達到最高值44.55%;當含水量為50%時,酸溶蛋白含量達到最高值9.22%;當含水量為30%時,酸性蛋白酶和中性蛋白酶活性達到最高值,分別為161.47 U/g和711.82 U/g。在固態發酵中,傳質依賴于培養基的自由水,當含水量較低時,自由水不足,傳質效率較低,會阻礙微生物的生長,當含水量較高,發酵培養基的孔隙率降低,溶氧下降,會抑制霉菌的生長。因此選擇含水量30%、40%、50%進行后續試驗。
2.3 接種量對辣木籽粕生物轉化的影響
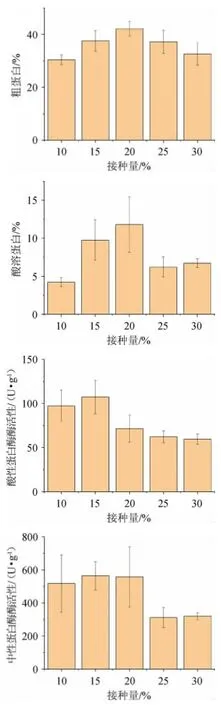
圖3 接種量對辣木籽粕生物轉化的影響Fig.3 Effect of inoculum on biotransformation of Moringa oleifera seed meal
由圖3可知,接種量對4個指標均有顯著影響(P<0.05)。在接種量為20%時,粗蛋白和酸溶蛋白含量均達到最高值,分別為42.11%和11.80%;當接種量為15%時,酸性蛋白酶和中性蛋白酶均達到最高值,分別為107.34 U/g和565.14 U/g。當接種量較低,微生物酶解發酵底物的起始效率較低,需要更多的時間生長繁殖才能對底物有較高的利用效率。隨著接種量的提高,培養基微生物量達到峰值的時間逐步縮短,不僅可以提高效率,而且由于接種微生物具有較大的生長優勢,可以減少污染雜菌的機會。當接種量過大,微生物生長過快會導致發酵底物營養物質消耗過快,代謝產物堆積,酶活性下降,過早進入衰亡期,微生物大量死亡,代謝效率下降。因此選擇接種量15%、20%、25%進行后續試驗。
2.4 發酵時間對辣木籽粕生物轉化的影響
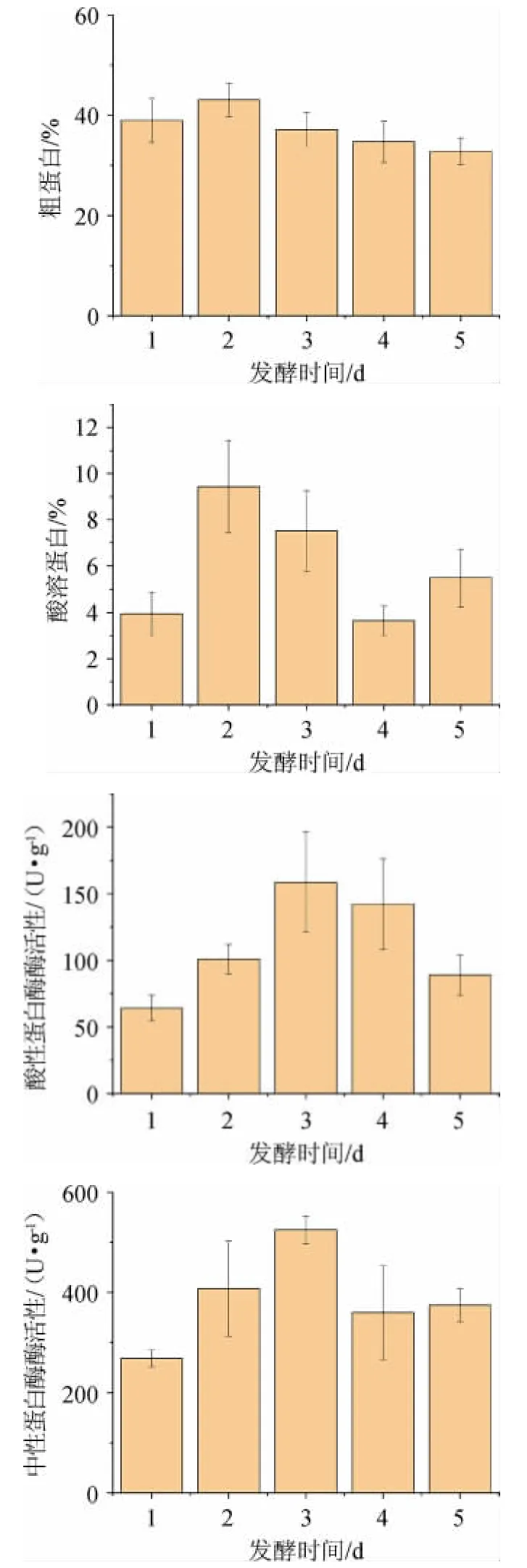
圖4 發酵時間對辣木籽粕生物轉化的影響Fig.4 Effect of fermentation time on biotransformation of Moringa oleifera seed meal
由圖4可知,發酵時間對4個指標均有顯著影響(P<0.05)。在發酵時間為2 d時,粗蛋白和酸溶蛋白達到最高值,分別為43.07%和9.44%;當發酵時間為3 d時,酸性蛋白酶和中性蛋白酶達到最高值,分別為158.72 U/g和524.77 U/g。隨著發酵時間的延長,微生物合成代謝產物逐步積累,但后期又再下降,推測是在發酵后期,微生物以分解代謝為主導,隨著微生物數量的增多,代謝產物不斷積累,微生物代謝受到抑制,酶活開始下降。因此選擇發酵時間1 d、2 d、3 d進行后續試驗。
2.5 工藝條件優化正交優化
在單因素試驗的基礎上,選擇接種順序(A)、發酵培養基含水量(B)、接種量(C)和發酵時間(D)等4個因素進行L9(34)正交試驗考察發酵條件對辣木籽粕發酵前后兩種蛋白含量和兩種蛋白酶活性的影響,正交試驗結果與分析見表2,極差分析見表3,方差分析見表4。
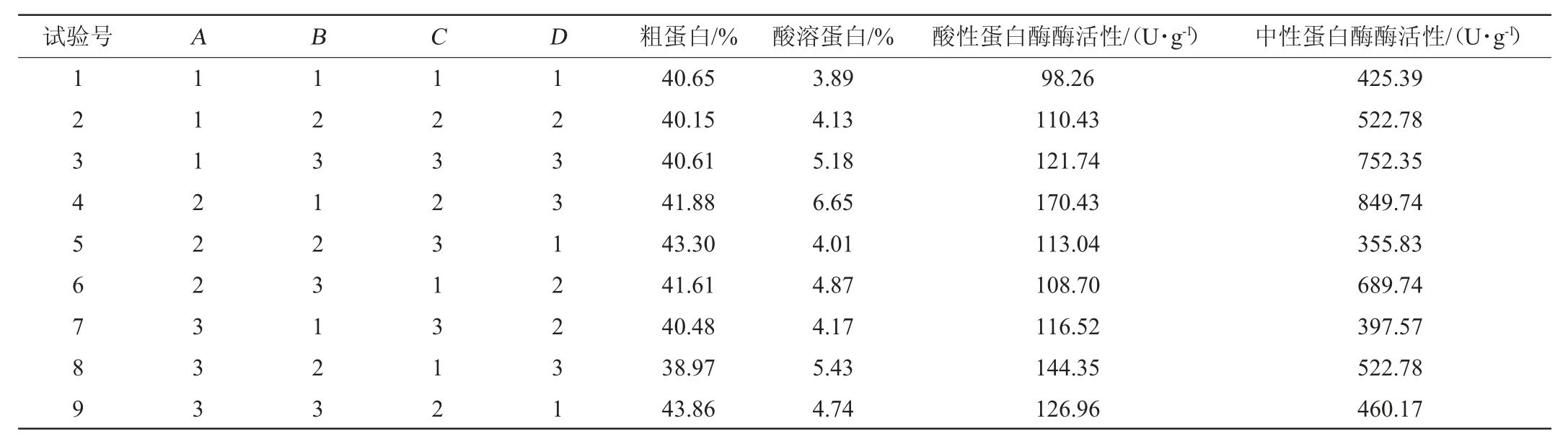
表2 辣木籽粕發酵條件優化正交試驗結果與分析Table 2 Results and analysis of orthogonal experiments for fermentation conditions optimization of Moringa oleifera seed meal
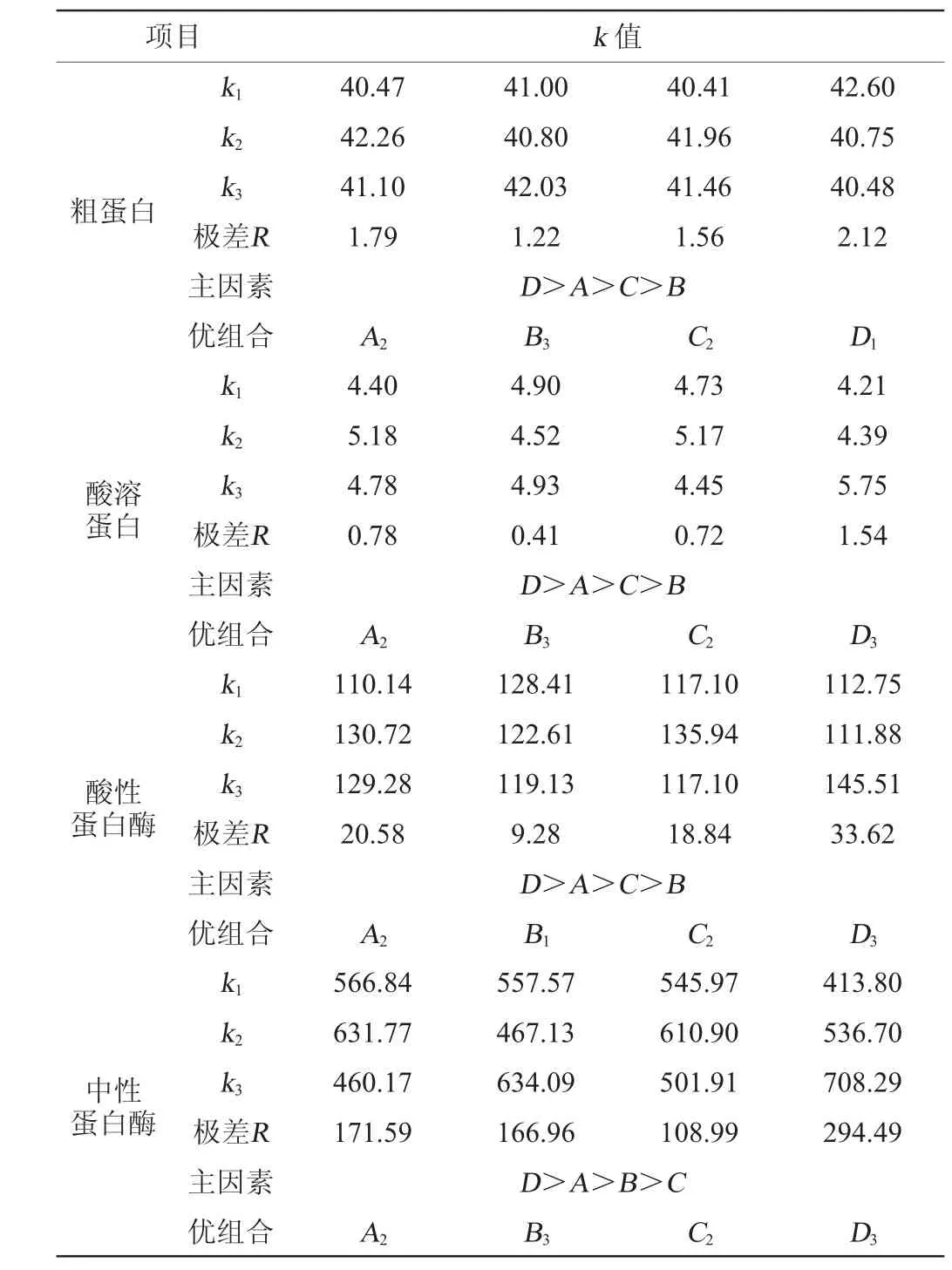
表3 正交試驗結果極差分析Table 3 Range analysis of orthogonal experiments results
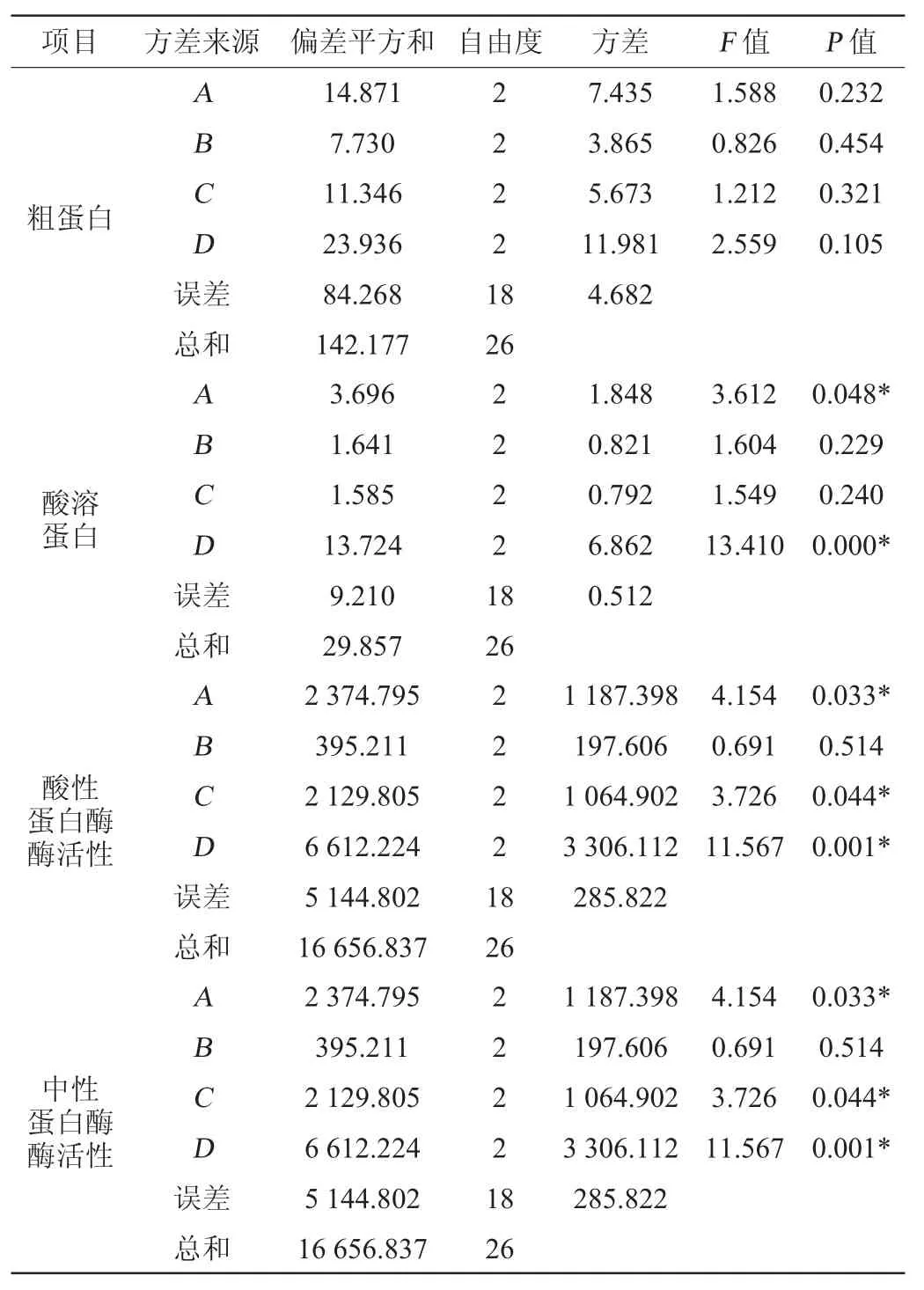
表4 正交試驗結果方差分析Table 4 Variance analysis of orthogonal experiments results
由表3可知,對于因素A和因素C,直接取A2、C2水平。根據綜合平衡法,因素B、因素D分別選取B3、D3水平。因此,固態發酵培養基的優組合為A2B3C2D3,即先接種米根霉和黑曲霉,發酵36 h后接種屎腸球菌,培養基含水量50%,接種量20%(接種比例1∶1∶1),30 ℃條件下發酵3 d。該組合進行驗證試驗(重復3次),與未發酵的辣木籽粕相比,得到的發酵辣木籽粕粗蛋白含量為46.78%,提高了32.22%;酸溶蛋白含量為11.30%,提高了121.57%;酸性蛋白酶活性為77.06 U/g,中性蛋白酶活性為711.93 U/g。除酸性蛋白酶活性較低外,粗蛋白含量、中性蛋白酶活性與正交試驗對應指標最高值(分別為43.86%和849.74 U/g)無顯著差異(P>0.05),酸溶蛋白含量顯著高于第4組(為6.65%)(P<0.05),除酸性蛋白酶活性較低外,其余指標均達到該指標的最高水平,該組合為較優組合。
由方差分析(見表4)可知,在正交試驗條件范圍內,4個因素對粗蛋白沒有顯著影響(P>0.05);接種順序與發酵時間對酸溶蛋白有顯著影響(P<0.05),發酵培養基含水量和接種量對酸溶蛋白無顯著影響(P>0.05);接種順序、接種量和發酵時間對酸性蛋白酶有顯著性影響(P<0.05),含水量對酸性蛋白酶無顯著影響(P>0.05);接種順序、含水量和發酵時間對中性蛋白酶具有顯著性影響(P<0.05),接種量對中性蛋白酶無顯著性影響(P>0.05),與極差分析結果相符。
3 結論
屎腸球菌、米根霉和黑曲霉混菌發酵辣木籽粕可提高其粗蛋白及酸溶蛋白含量,菌種接種順序對發酵效果有顯著影響,最佳條件為先接種黑曲霉和米根霉,發酵36 h后,再接種屎腸球菌,在培養基含水量50%,總接種量20%的條件下30 ℃發酵3 d。與未發酵的辣木籽粕相比,發酵后辣木籽粕粗蛋白達46.78%,提高了32.22%,酸溶蛋白達11.30%,提高了121.57%,同時酸性蛋白酶活性達77.06 U/g,中性蛋白酶活性達711.93 U/g。可見,發酵辣木籽粕綜合營養價值得到提高,具有更好的消化性能。該結果為辣木籽粕深加工開發提供了依據。
