基于線粒體COⅠ基因序列的東南太平洋莖柔魚群體遺傳結構分析
2020-06-30 08:08:48蔣科技黃洪亮馬春艷張鳳英陳雪忠馬凌波
海洋漁業 2020年3期
刁 樂,宋 煒,蔣科技,黃洪亮,馬春艷,張鳳英,陳雪忠,馬凌波
(1.中國水產科學研究院東海水產研究所,農業農村部遠洋與極地漁業創新重點實驗室,上海 200090;2.上海海洋大學水產與生命學院,上海 201306)
莖 柔 魚 (Dosidicus gigas),屬 槍 形 目(Teuthoidea),柔魚科(Ommastrephidae),莖柔魚屬,是大洋性淺海種,廣泛分布于東南太平洋海域。在緯度方向上,分布范圍從美國加利福尼亞(37°N)到南美洲智利(47°S)的海域,在經度方向上,在赤道附近可向西延伸至125°W海域[1-3],也有學者認為其在赤道分布范圍可擴展到140°W(圖1)[4]。莖柔魚是頭足類中資源量最豐富的種類之一[5],目前商業性開發的漁場有加利福尼亞南部沿岸及外海漁場、墨西哥沿岸及外海漁場、哥斯達黎加沿岸及外海漁場、秘魯西海岸及外海漁場和智利外海漁場等。2004—2008年間,莖柔魚年產量穩定在70~90萬t之間[6]。其中,秘魯外海年產量最高達20萬t[7]。
目前關于莖柔魚的研究主要集中在資源豐度及 分 布[8-13]、攝 食 習 性[14-15]、生 物 學 特性[16-17]及漁場分布[18-20]等方面。在種群結構研究中主要集中在依據胴長進行劃分[21-22],也有根據繁殖季節、耳石、內殼、角質顎結構進行劃分[23-25], 例 如, NIGMATULLIN 等[3]、ARGUELLES等[26]和 YATSU等[27]分別根據成年雄性與雌性個體的胴長、性成熟胴長大小和產卵季節等對莖柔魚種群進行劃分。
隨著分子生物學技術的快速發展,運用線粒體Cytb、COI序列,微衛星標記等方法對莖柔魚的群體遺傳結構研究也逐漸增多[28-33]。mtDNA具有結構簡單、母系遺傳、進化速率快、重組率低等特點,作為一種重要的分子遺傳標記而被廣泛應用于水產生物的系統發育、生物地理學和保護生物學研究[34]。其中,細胞色素氧化還原酶Ⅰ(COⅠ)基因進化速率適中,適合種群水平差異的檢測[35-36],已廣泛應用于頭足類群體遺傳變異和系統發育研究中[28]。
莖柔魚生命周期短,大約為1年,且終生只產1次卵,其資源量很容易受到海洋環境的變化而發生波動[27,37]。本研究通過線粒體 DNACOⅠ序列的多態性對秘魯外海東南太平洋莖柔魚群體進行了遺傳結構分析,以期為其合理開發與利用提供理論依據。
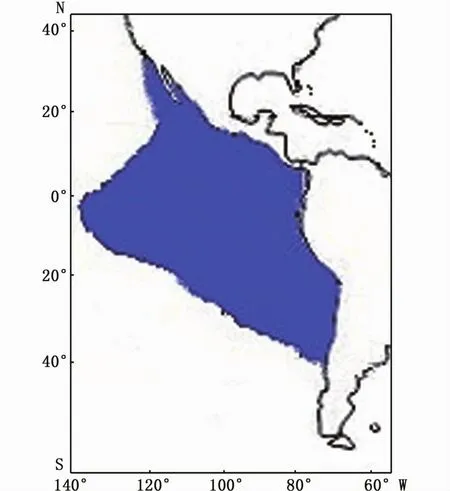
圖1 莖柔魚分布Fig.1 Distribution of Dosidicus gigas
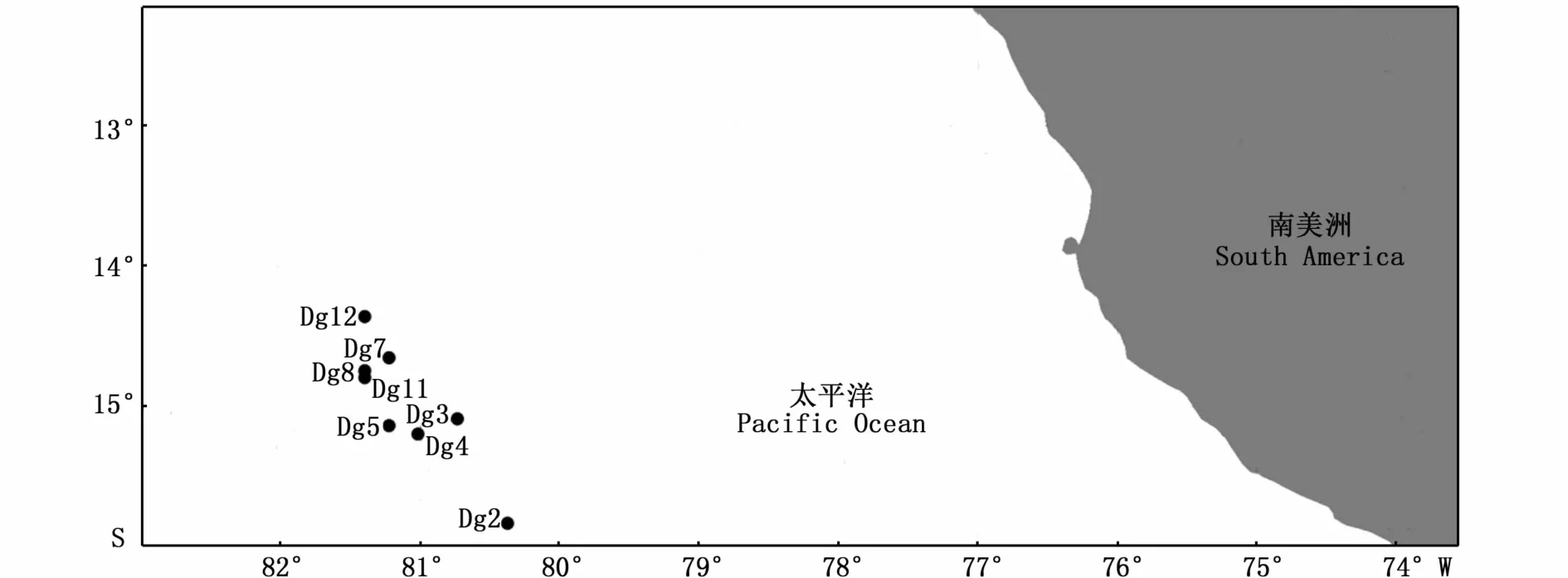
圖2 莖柔魚采樣地點示意圖Fig.2 Sampling sites of Dosidicus gigas
1 材料與方法
1.1 實驗材料
實驗用莖柔魚于2014年6—7月采集于秘魯外海(圖2)。具體采集的地點、時間和樣品數見表1。樣品采集后取套膜肌肉組織,置于95%乙醇中,-20℃保存備用。
1.2 基因組DNA的提取
采用海洋動物組織基因組DNA提取試劑盒(北京,天根生化科技有限公司)提取總DNA,提取的DNA經1%的瓊脂糖凝膠電泳檢測質量后,保存于-20℃備用。
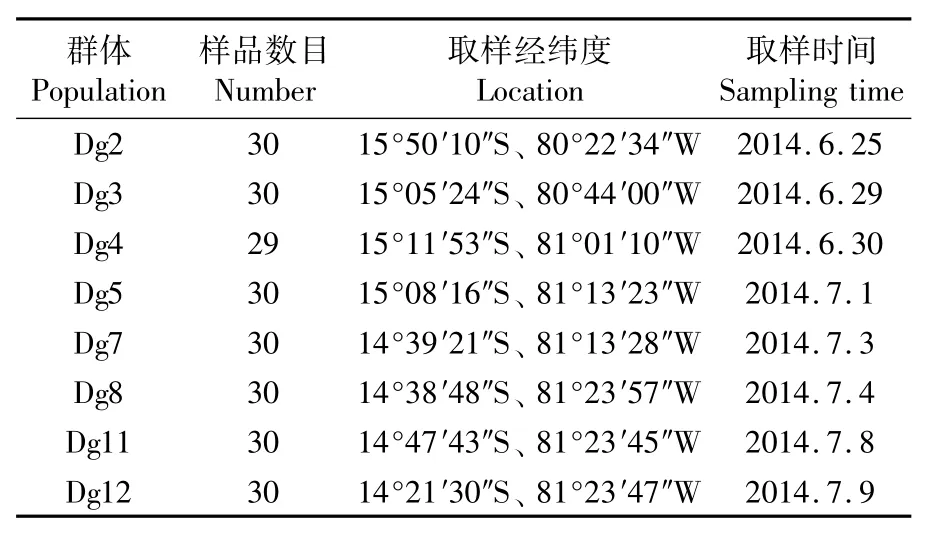
表1 莖柔魚的采集樣品數Tab.1 Sample number of Dosidicus gigas in each location
1.3 莖柔魚mtDNA COⅠ序列的擴增與測序
使用兼并引物Dg-COI-F(TCCACAAAYCAY AAAGATATTGAAC)和Dg-COI-R(ACYTCTGGGT GMCCAAARAATCA)擴增莖柔魚mtDNACOⅠ部分序列,PCR總體系為25μL:10×PCR Buffer(Mg2+plus)2.5μL,dNTP(2.5 mmol·L-1)2μL,rTaqDNA聚合酶(5 U·μL-1)0.5μL,上下游引物(10μmol·L-1)各 0.75μL,模板 DNA 1μL,加雙蒸水至總體積25μL。PCR反應程序為:94℃預變性 5min;94℃變性 30 s,50℃退火 45 s,72℃延伸50 s,35個循環;最后72℃充分延伸7 min;10℃保存。擴增產物使用1.2%的瓊脂糖凝膠電泳檢測后送上海杰李生物技術有限公司正向測序。
1.4 數據分析
利用MEGA 5.1中的Clustal W程序對序列進行排序校正,按照TrN進化模型來構建單倍型鄰近關系樹,系統樹的可靠性采用1 000次重復抽樣評估。同時計算群體內及兩兩群體間的遺傳距離,根據Kimura-2-parameter(K-2-P)模型構建莖柔魚群體間的UPGMA樹。采用遺傳分化指數(Fst)來評價兩兩群體間的遺傳差異,通過1 000次重復抽樣來檢驗兩兩群體間Fst的顯著性,使用Nm=(1Fst)/(4Fst)計算兩兩群體間的基因交流。
利用DnaSP 5.1軟件確定單倍型數目及分布、核苷酸多樣性。使用Arlequin 3.1軟件的分子方差分析(AMOVA)來評估群體間遺傳變異,通過1 000次重復抽樣來檢驗不同遺傳結構水平上的協方差的顯著性。采用Fu’sFs和Tajima’sD中性檢驗和核苷酸不配對分布來檢驗群體歷史動態,以此指示種群歷史的穩定性或種群增長情況。
2 結果與分析
2.1 序列變異特征分析
經測序和校對,獲得626 bp線粒體COⅠ基因片段。所得序列共包含42個變異位點,其中單一信息位點34個,占80.95%;多態簡約位點8個,占19.05%。存在33個轉換位點和8個顛換位點,另外1個位點同時存在轉換和顛換現象,沒有檢測到插入和缺失現象。A、T、G和C平均含量分別為 26.5%、35.3%、15.8%和 22.4%,A+T含量(61.8%)明顯高于 G+C含量(38.2%)。
2.2 單倍型在群體中的分布及遺傳關系
在所檢測的莖柔魚239個樣本中共出現46種單倍型(MK336912~MK336957),但單倍型間的序列分歧值很低,與單倍型Hap1相比,其余45種單倍型中有33種與其只存在1個核苷酸的差異,詳見圖3。群體間共享的單倍型為12個,占比26.0%,其余34個單倍型為某個群體所特有,各群體單倍型分布見圖4。Hap44為所有群體共享,其擁有頻率也最高,共150個個體擁有,占比62.7%。Hap43和Hap45為7個群體所共享,分別有15、14個個體擁有此單倍型。Hap30為4個群體共享,Hap13和Hap27分別有3個群體共享,Hap8、Hap15、Hap20、Hap22、Hap38和 Hap46均為2個群體共享。在特有的單倍型中,Dg2、Dg3、Dg4、Dg5、Dg7、Dg8、Dg11、Dg12所特有的單倍型個數分別為 7、3、4、3、4、2、4、10個。
2.3 群體遺傳多樣性和遺傳結構分析
東南太平洋莖柔魚8個群體的遺傳多樣性參數見表2。基于COⅠ基因所有序列得到的8個群體總的單倍型數、單倍型多樣性指數、多態位點數和核苷酸多樣性指數分別為46、0.599±0.038、42和0.001 38±0.001 71。從單倍型參數來看,Dg12群體的單倍型數和單倍型多樣性指數均最高,分別為 16和 0.759±0.086;Dg7群體的單倍型多樣性指數最低,為0.469±0.114。從核苷酸多樣性參數來看,Dg11群體的核苷酸多樣性指數最高,為0.003 63±0.001 21;Dg5群體的核苷酸多樣性指數最低,為 0.001 04±0.001 07。總體來看,各群體間遺傳多樣性無明顯差異,均具有較高的單倍型多樣性指數與較低的核苷酸多樣性指數,這與閆杰等[37]、劉連為等[30,32-33]得到的結果一致。
分析秘魯外海8個莖柔魚群體間的遺傳距離,群體間的平均遺傳距離在0.001 04~0.002 01之間。其中,Dg2與Dg12之間的遺傳距離最大(0.002 01),Dg5與 Dg7之間的遺傳距離最小(0.001 04)(表3)。從 UPGMA樹來看(圖5),秘魯外海莖柔魚群體間沒有明顯的分化關系,Dg5與Dg7先聚在一起然后依次與其他群體聚在一起,最后與Dg2聚在一起。

圖3 莖柔魚線粒體COⅠ基因序列變異位點Fig.3 Variable sites ofm itochondrial COⅠgene of Dosidicus gigas
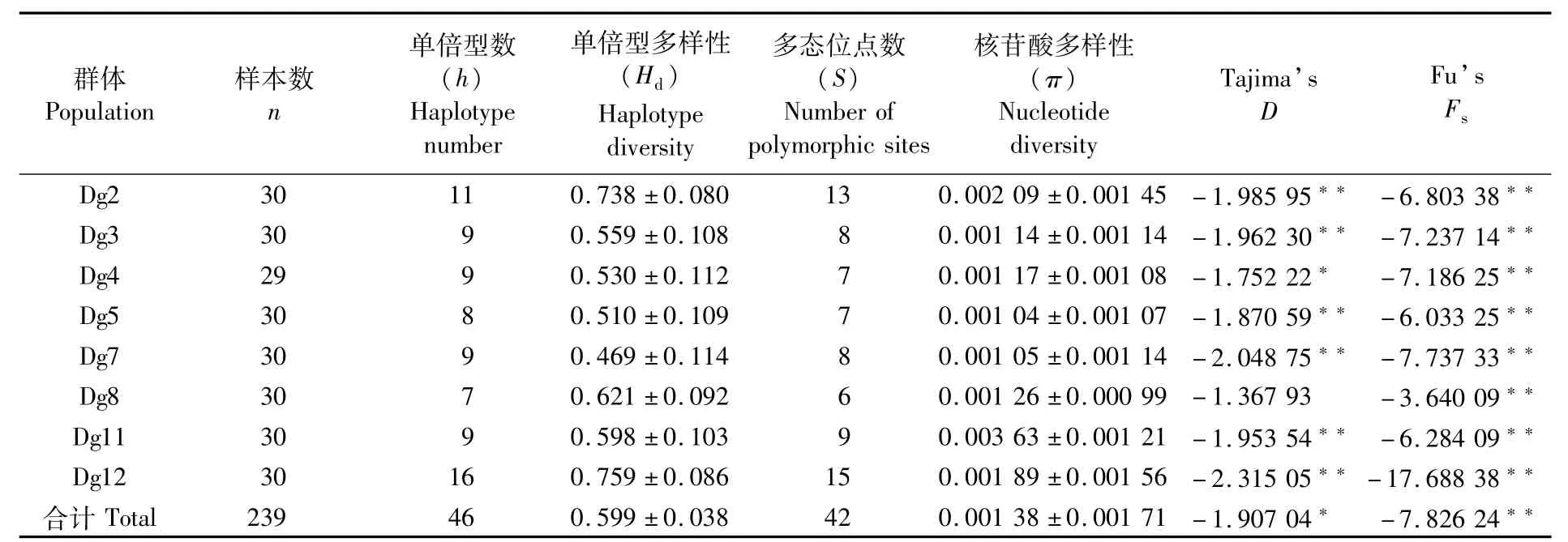
表2 不同莖柔魚群體的遺傳多樣性參數Tab.2 Parameters of genetic diversity of different populations of Dosidicus gigas
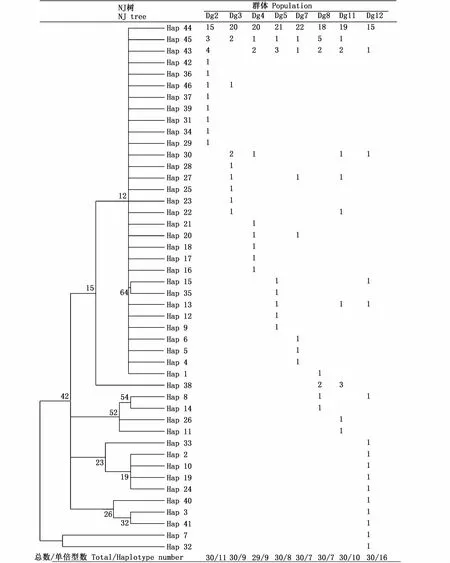
圖4 莖柔魚46個單倍型在8個群體中的分布及其分子系統樹Fig.4 Distribution of 46 haplotypes among 8 Dosidicus gigas populations and its NJmolecular phylogenetic tree
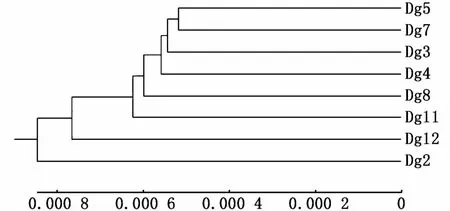
圖5 基于K im ura遺傳距離的秘魯外海莖柔魚8個群體之間的UPGMA樹Fig.5 UPGMA tree of Dosidicus gigas populations built under K-2-P genetic distance
兩兩群體間的遺傳分化系數Fst值為-0.019 12~0.027 94之間(表 3),統計檢驗均不顯著(P>0.05),說明秘魯外海莖柔魚群體間存在高度的遺傳同質性,不存在顯著差異的遺傳結構分化。基于Fst值計算群體間基因流Nm,各群體間的Nm均大于1,說明各群體間具有較高的基因交流水平(表4)。
AMOVA分析結果顯示,群體內變異和群體間的變異百分比分別為99.74%和0.26%,表明群體間不存在顯著的遺傳結構(表5)。
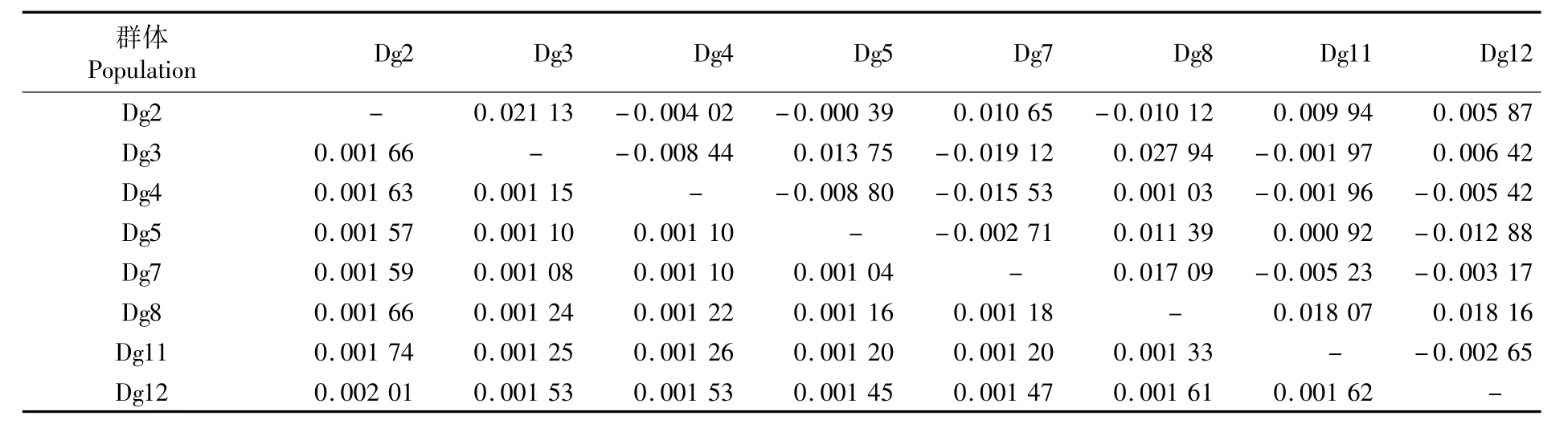
表3 莖柔魚群體間遺傳距離(對角線下)和遺傳分化系數(F st)(對角線上)Tab.3 Pairw ise genetic distances(below diagonal),fixation index(F st)(above diagonal)between every two populations of Dosidicus gigas
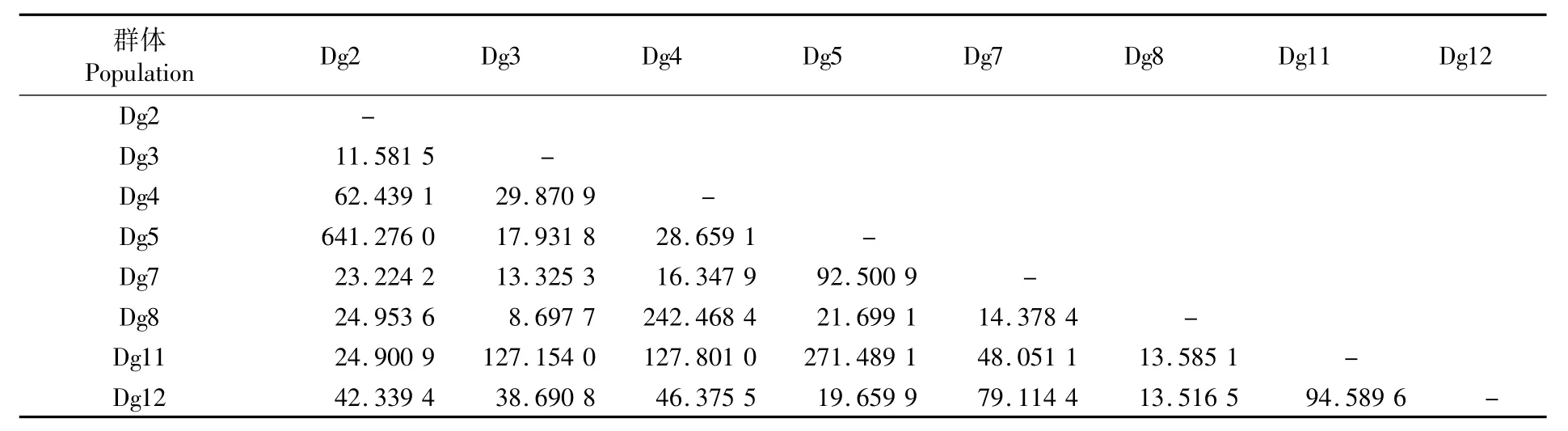
表4 莖柔魚兩兩群體間的基因流(對角線下)Tab.4 Gene flow value(below diagonal)between every two populations of Dosidicus gigas
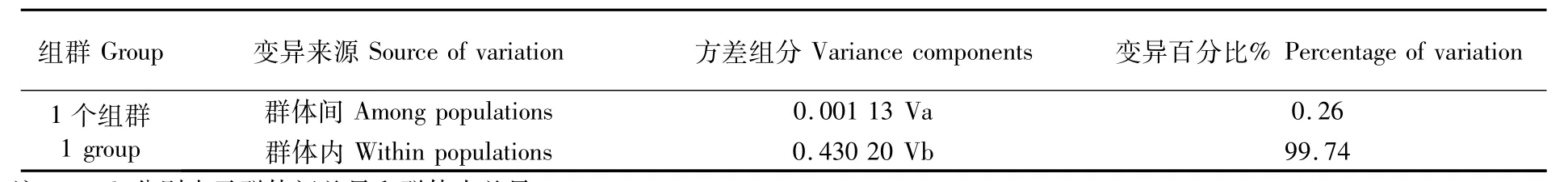
表5 莖柔魚群體遺傳差異的AMOVA分析Tab.5 Analysis ofmolecular variance(AMOVA)among populations of Dosidicus gigas
2.4 群體歷史動態分析
對Tajima’sD和 Fu’sFs值進行中性檢驗(表2)。Tajima’sD值均呈負值,且呈顯著差異,Fu’sFs值同樣均呈負值,經檢驗呈極顯著差異,研究表明莖柔魚群體在東太平洋范圍內曾經經歷過種群擴張事件。
3 討論
3.1 外部形態特征與COⅠ基因判別東南太平洋莖柔魚種群的解釋
頭足類屬于常年產卵、生長迅速且生命周期短的物種。陳新軍等[38]認為,長度頻度分析方法不適合用來研究頭足類的年齡和生長。此外,胡貫宇等[39]研究表明頭足類還具有洄游習性,導致不同世代的群體混合在一起,因而在進行日齡和生長的分析時無法排除不同世代、不同群體之間的干擾[40]。NIGMATULLIN等[3]還發現莖柔魚小個體群體主要分布在赤道附近,大個體群體主要分布在高緯度海域,這可能是因為在溫度較高的環境下,莖柔魚性成熟加快,個體較小[41];相反,在溫度較低的環境下,莖柔魚性成熟緩慢,個體較大[39]。因此,影響莖柔魚胴長的因素有很多,其中包含地理環境或物理因素,即利用胴長等外部形態特征可能很難判別東南太平洋莖柔魚種群屬性。
線粒體DNA(mtDNA)中,細胞色素氧化還原酶Ⅰ(COⅠ)基因已被證明適合頭足類群體遺傳變異和系統發育的研究且已廣泛應用[28]。劉連為等[32]針對 NIGMATULLIN等[3]劃分的大型群與中型群莖柔魚,利用mtDNA與SSR 2個分子標記對其進行分析,研究表明,大型群與中型群莖柔魚可能在產卵洄游過程中發生基因交流,建議將秘魯外海大型群與中型群莖柔魚看作1個管理單元。閆杰等[38]利用mtDNA對東太平洋公海莖柔魚種群遺傳結構進行了初步研究,認為可以大致將東太平洋公海莖柔魚以赤道為界分為南方和北方2個地理群,即將東南太平洋莖柔魚視為一個種群,本研究結果與其一致。
3.2 莖柔魚群體遺傳多樣性分析
遺傳多樣性是指地球上所有生物所攜帶的遺傳信息的總和,也是物種適應復雜多變的環境、維持生存和進化的重要基礎。一個物種的遺傳變異越大,其進化潛力就越大,對環境變化的適應能力也就越強[42]。本研究運用 mtDNACOⅠ序列對東南太平洋莖柔魚8個群體的遺傳多樣性進行了分析。結果表明,Dg12群體的單倍型數和單倍型多樣性均最高,Dg12群體在這8個群體中最靠近赤道海域,在赤道海域海流等作用下,可能與赤道海域的莖柔魚有基因交流,而這也是其單倍型多樣性最高的原因之一,這與劉連為等[33]研究結果一致。
本研究239個樣本中共獲得46個單倍型,Hap44為所有群體共享,其為150個個體擁有,推測Hap44很可能是較原始的單倍型類型[43]。種群共享及個體共有單倍型出現的次數多,說明秘魯外海莖柔魚間基因交流頻繁。8個群體間均表現為高單倍型多樣性(h>0.5)、低核苷酸多樣性(π<0.005)的模式,說明種群經歷了顯著的群體擴張,這與閆杰等[37]和劉連為等[33]的研究結果一致。根據GRANT等[44]劃分的4種單倍型多樣性和核苷酸多樣性關系模式,本研究莖柔魚群體屬于第二種模式,即高單倍型多樣性和低核苷酸多樣性。龐大的種群數量是維持自然種群內具有較高單倍型多樣性的主要基礎之一。通常一個小的有效種群經過近期快速增長成為一個大群體時,該種群單倍型多樣性會非常豐富,但沒有足夠時間使核苷酸產生變異,這就造成了“高h、低π”的遺傳多樣性模式[43]。莖柔魚是短生命周期種類,性成熟周期短,繁殖季節較長,這樣的生活史特征對群體快速增長、產生高h具有一定的促進作用。
3.3 莖柔魚群體遺傳結構分析
本研究單倍型鄰近關系樹及群體間的UPGMA進化樹均表明,東南太平洋莖柔魚群體間不具有顯著的系統地理格局。兩兩群體間的Fst值以及AMOVA分析結果也同樣表明,群體間的遺傳差距不明顯,不存在顯著差異的群體遺傳結構。閆杰等[37]、劉連為等[33]對東太平洋莖柔魚群體間遺傳結構的分析,均顯示各群體間不存在顯著的遺傳結構,本文的結果與其一致。而對秘魯外海大小型莖柔魚群體和北太平洋莖柔魚不同產卵群體間結構的分析結果顯示,秘魯外海大小型群體、北太平洋不同產卵群體間也均不存在顯著的遺傳結構[32]。
在缺少擴散障礙的海洋環境中,海洋生物的卵、幼體、成體因具有較強的擴散能力從而使群體間產生頻繁的基因交流,這將導致海洋生物在非常廣闊的分布范圍內具有遺傳同質性[43]。東南太平洋海域位置開闊,無地理阻礙,東太平洋在赤道南部存在洋流的閉合環路,海水從高緯度流向赤道,在秘魯海域形成上升流系統和秘魯離岸流。莖柔魚具有較強的游泳能力及長距離的季節洄游習性,使東南太平洋海域莖柔魚群體間不存在基因交流障礙,導致莖柔魚群體間的遺傳差距并不明顯。簡言之,秘魯外海域缺乏地理障礙,秘魯洋流、離岸流的作用、季節洄游習性以及其較強的運動能力,促使該海域莖柔魚群體間沒有顯著的遺傳結構。
綜上所述,本研究中東南太平洋取樣點的莖柔魚群體間不存在顯著的群體遺傳分化。為了更加準確地確定莖柔魚群體間的遺傳結構,在后續的研究中應該擴大取樣范圍,同時補充不同季節、大小型群體的樣本,采用不同的分子標記,進行系統分析,為莖柔魚漁業資源的管理提供豐富的數據支持。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
哲學評論(2021年2期)2021-08-22 01:53:34
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
