IRF8基因降表達、PINK1基因過表達對肺泡巨噬細胞NR8383增殖與活性的影響
2020-06-29 05:10:50李東航范濤張霖江文洋汪巍耿慶
山東醫(yī)藥 2020年17期
李東航,范濤,張霖,江文洋,汪巍,耿慶
武漢大學人民醫(yī)院,武漢430060
作為體內重要的吞噬和抗原提呈細胞,肺泡巨噬細胞在維持肺部免疫穩(wěn)態(tài)及宿主防御中發(fā)揮重要作用,并參與了急性肺損傷(ALI)在內的多種肺部疾病的發(fā)病過程[1]。干擾素調節(jié)因子8(IRF8)是一種主要表達于免疫細胞的IFN-γ誘導型核轉錄因子,在調控轉錄及調節(jié)宿主免疫、細胞因子信號轉導、細胞生長及分化等生物學過程中發(fā)揮重要作用[2]。除了調節(jié)淋巴細胞、髓細胞、樹突狀細胞的增殖分化外,IRF8還可以刺激巨噬細胞分化,抑制粒細胞分化,同時抑制細胞增殖,正向調節(jié)凋亡[3,4]。已有研究表明,IRF8是巨噬細胞中的主要自噬調節(jié)劑,可以直接激活許多涉及自噬各步驟的基因,從而促進自噬體形成和溶酶體融合,對巨噬細胞的成熟、活性和先天免疫反應至關重要[5]。PTEN誘導激酶1(PINK1)基因可以編碼具有絲氨酸-蘇氨酸激酶活性的蛋白質,其N末端為線粒體靶向序列,主要在線粒體中發(fā)揮作用[6]。作為線粒體自噬經典途徑,PINK1/Prakin信號通路介導的線粒體自噬參與多種原因導致的肺損傷過程[7,8]。2019年1~8月,我們觀察了shRNA編碼干擾IRF8基因、巨細胞病毒介導PINK1基因過表達及二者共同作用后肺泡巨噬細胞NR8383增殖能力及活性變化,探討IRF8是否通過調控巨噬細胞中PINK1相關線粒體自噬對缺血再灌注肺損傷產生影響。
1 材料與方法
1.1 主要材料與試劑 IRF8基因降表達質粒pG2.2-IRF8-Rat-444、PINK1過表達質粒pCMV3-Rat-PINK1、IRF8干擾空質粒、PINK1過表達空質粒均由實驗室構建,并經測序鑒定;小鼠肺泡巨噬細胞NR8383,F12K培養(yǎng)基、血清、0.25% Trypsin、opti-MEM(Gibco公司);LipofectamineTM2000(Invitrogen公司);兔多抗PINK1(Abclonal公司),兔多抗IRF8(Thermop公司),兔多抗GAPDH(杭州賢至生物有限公司);BCA試劑盒和RIPA裂解液(碧云天生物科技公司);PVDF膜(Millipore公司);CCK8細胞增殖及細胞毒性檢測試劑盒(Beyotime公司);乳酸脫氫酶(LDH)測試試劑盒(南京建成生物工程研究所);其他試劑均為進口或國產分析純。
1.2 實驗方法
1.2.1 細胞培養(yǎng)與轉染 解凍NR8383細胞并培養(yǎng)至細胞融合度達80%,對細胞進行傳代培養(yǎng);將對數生長期的細胞制成單細胞懸液,按5×105/孔接種6孔板,于37 ℃、5% CO2、飽和濕度培養(yǎng)至細胞融合度達60%。取對數生長期的NR8383細胞,隨機分為7組。IRF8對照組、PINK1對照組分別轉染IRF8干擾空質粒、PINK1過表達空質粒,聯(lián)合對照組共轉染用量減半的上述兩種質粒;IRF8干擾組、PINK1過表達組分別轉染IRF8干擾質粒pG2.2-IRF8-Rat-444、PINK1過表達質粒pCMV3-Rat-PINK1,聯(lián)合干擾組共轉染上述兩種質粒;正常對照組不做任何處理。轉染前2 h,換成無血清相應培養(yǎng)基;用100 μL無血清opti-MEM稀釋6 μg質粒(兩種質粒共轉時,每種質粒用量減半),取12 μL混勻的LipofectamineTM2000稀釋在100 μL opti-MEM中;每個培養(yǎng)孔加轉染復合物混合液200 μL,將細胞于37 ℃ CO2培養(yǎng)箱中培養(yǎng);4 h后吸出混合液換入正常培養(yǎng)基,于37 ℃、5% CO2、飽和濕度下培養(yǎng)48 h;熒光顯微鏡觀察各孔中綠色熒光蛋白表達情況,可見到NR8383細胞中大量表達,表明轉染成功。
1.2.2 細胞增殖能力觀察 采用CCK8法。收集各組細胞以胰酶消化,用相應培養(yǎng)基制成5×104/mL的單細胞懸液。以100 μL/孔將細胞接種96孔板,同時設置空白對照孔;周圍孔加入100 μL PBS,于37 ℃、5% CO2、飽和濕度下培養(yǎng)過夜。每孔加入10 μL CCK8,37 ℃培養(yǎng)4 h,振蕩10 min,用酶標儀測定各孔波長在450 nm時的光密度值(OD450),計算各組細胞增殖率。細胞增殖率=(實驗孔OD450-空白孔OD450)/對照孔OD450×100%。
1.2.3 細胞活性觀察 采用LDH法。取各組細胞上清,加入LDH測試試劑盒中;根據使用說明書混勻,室溫放置5 min;酶標儀測定波長450 nm處各孔OD值,計算LDH活性;以LDH活性表示細胞活性,值越高則細胞活性越弱。LDH活性(U/L)=(測定孔OD值-對照孔OD值)/(標準孔OD值-空白孔OD值)×0.2×1 000×樣本稀釋倍數。
1.2.4 細胞中IRF8、PINK1蛋白表達檢測 采用Western blotting法。取各組對數生長期細胞,提取細胞總蛋白,經BCA試劑盒測得總蛋白濃度;裂解液中加入蛋白上樣緩沖溶液,沸水浴 10 min變性;配置蛋白電泳凝膠,進行10% SDS-PAGE。用濕轉法將凝膠上的蛋白轉移到PVDF膜上,電轉結束后用含5%脫脂奶粉的TBST溶液封閉2 h;4 ℃孵育一抗(GAPDH、PINK1、IRF8)過夜,TBST漂洗5 min×5次;室溫孵育二抗2 h,TBST漂洗5 min×6次;加入ECL試劑,掃描膠片;用BandScan分析膠片灰度值,以目的蛋白與內參GAPDH灰度值比值表示目的蛋白相對表達量。
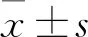
2 結果
2.1 各組細胞增殖與細胞活性比較 見表1。
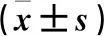
表1 各組細胞增殖與細胞活性比較
注:與聯(lián)合對照組、PINK1對照組、IRF8對照組、正常對照組比較,*P<0.05;與PINK1過表達組、IRF8干擾組比較,#P<0.05。
2.2 各組IRF8、PINK1蛋白表達比較 見表2。
表2 各組PINK1、IRF8蛋白表達比較
注:與聯(lián)合對照組、PINK1對照組、IRF8對照組、正常對照組比較,*P<0.05;與PINK1過表達組、IRF8干擾組比較,#P<0.05。
3 討論
PINK1/Parkin相關線粒體自噬是PINK1通過其N-末端線粒體信號序列在功能失調的線粒體外膜募集,發(fā)生自磷酸化并磷酸化泛素和Parkin,選擇性清除損傷、衰老線粒體的過程,對于維持線粒體的能量供應及細胞內環(huán)境穩(wěn)態(tài)發(fā)揮著重要作用[9]。在ALI引起的炎癥反應中,不同病理階段的微環(huán)境下,肺泡巨噬細胞作為前哨發(fā)揮著促炎和抗炎的雙重作用,被認為是ALI/急性呼吸窘迫綜合征(ARDS)發(fā)病機制的關鍵因素之一[1]。由此,調節(jié)肺泡巨噬細胞的功能,可能是治療包含ALI/ARDS在內的肺損傷最具前景的治療策略[1,5,10]。缺血再灌注肺損傷是導致線粒體受損的關鍵因素[11],因此引發(fā)肺泡巨噬細胞生存及穩(wěn)定的PINK1相關線粒體自噬就尤為重要[12~15]。基于IRF8是驅動巨噬細胞分化的重要轉錄因子,且為巨噬細胞自噬的主要調節(jié)劑之一。我們推測,IRF8在發(fā)生缺血再灌注肺損傷的肺泡巨噬細胞PINK1/Prakin相關線粒體自噬中發(fā)揮著調控作用。
本研究結果顯示,PINK1過表達組、IRF8干擾組肺泡巨噬細胞增殖及細胞活性受到顯著抑制,且聯(lián)合干擾組增殖能力及細胞活性的抑制作用更明顯;與PINK1過表達組、IRF8干擾組比較,聯(lián)合干擾組細胞活性抑制效應差異有統(tǒng)計學意義,但對細胞增殖能力抑制作用差異無統(tǒng)計學意義。這表明干擾IRF8基因、誘導PINK1基因過表達,在抑制肺泡巨噬細胞的增殖及活性方面具有協(xié)同作用。此外,與聯(lián)合對照組、PINK1對照組、IRF8對照組、正常對照組比較,IRF8干擾組PINK1蛋白表達增加、PINK1過表達組IRF8蛋白表達下降,且差異均有統(tǒng)計學意義;與PINK1過表達組、IRF8干擾組比較,聯(lián)合干擾組PINK1蛋白表達增加、IRF8蛋白表達下降程度更明顯,差異均有統(tǒng)計學意義。這表明干擾IRF8蛋白可以促進PINK1蛋白的表達,干擾IRF8基因對PINK1基因的轉錄具有正向調控作用。
IRF8對巨噬細胞的作用體現在調節(jié)其發(fā)育和免疫功能相關基因來介導巨噬細胞參與的炎癥反應,以及自身結合PU.1形成復合物與其他轉錄因子作用執(zhí)行其吞噬功能,抑制IRF8基因表達會明顯降低巨噬細胞的數量[5]。Gupta等[5]發(fā)現,在敲除了巨噬細胞的IRF8基因后,多數自噬基因下調、少數上調,泛素結合蛋白過度積聚,巨噬細胞的自噬活性下降,自噬體難以成熟為溶酶體;在轉入野生型小鼠的IRF8基因后,巨噬細胞內部分自噬基因恢復了表達,但轉入IRF8K79E突變基因并不能恢復自噬基因的表達。本研究結果發(fā)現,干擾IRF8基因表達,肺泡巨噬細胞數量明顯下降,增殖能力與細胞活性均降低。
PINK1基因在維持線粒體形態(tài)和功能方面有重要作用[16,17],PINK1基因的損傷缺失將導致線粒體形態(tài)發(fā)生改變,并可影響細胞內線粒體復合體的活性,引起線粒體的多種功能障礙,最終導致細胞凋亡[18]。在PINK1缺陷細胞中,自噬體的形成速率明顯高于PINK1未缺陷細胞;而恢復PINK1表達會抑制線粒體自噬,PINK1過表達則抑制毒素誘導的線粒體自噬[19]。本研究結果表明,PINK1基因過表達后,肺泡巨噬細胞活性與細胞增殖均被抑制,細胞內的線粒體自噬可能相應受到抑制;過表達的PINK1基因可能負向反饋調節(jié)IRF8基因的轉錄,表現為IRF8蛋白表達降低。
巨噬細胞的吞噬作用在先天性免疫中發(fā)揮著重要作用,參與了從抗原識別到加工、運輸呈遞以及最終吞噬殺滅等過程,在此過程中伴隨著大量能量的消耗,通常都由線粒體負責能量的供應[20]。線粒體被證實參與先天性免疫調控,線粒體抗病毒信號蛋白(MAVS)錨定于線粒體,是RIG-Ⅰ樣受體信號傳導中的關鍵接頭蛋白[21],線粒體DNA在細胞經歷非正常死亡(例如病理學損傷)后釋放并激活一系列先天性免疫通路[22]。根據本研究我們推測,干擾IRF8基因表達,可以抑制巨噬細胞增殖與細胞活性,同時上調PINK1基因表達,增加了這種抑制效應;另外,PINK1基因過表達負向調節(jié)IRF8蛋白產生,可能與減輕線粒體DNA損傷有關,二者協(xié)同抑制肺泡巨噬細胞內線粒體自噬的發(fā)生。本研究可為肺缺血再灌注損傷的治療提供新思路。但是,受IRF8調控的自噬基因分布范圍很廣,從最初的自噬體激活到捕獲靶標的最終溶酶體降解,基本上涵蓋了整個自噬途徑[5],而PINK1/Parkin相關線粒體自噬也非線粒體自噬的惟一途徑[23,24]。因此,IRF8與線粒體自噬基因之間的調控機制仍需分子通路水平的嚴謹實驗設計來研究證實。
