基于睪丸尺寸比較研究中緬樹鼩的婚配制度
2020-06-11 11:08:32諶立新張浩劉璐朱萬龍王政昆
四川動物 2020年3期
諶立新, 張浩, 劉璐, 朱萬龍*, 王政昆*
(1.云南省高校西南山地生態系統動植物生態適應進化及保護重點實驗室,云南師范大學生命科學學院,生物能源持續開發利用教育部工程中心,云南省生物質能與環境生物技術重點實驗室,昆明650500;2. 昆明衛生職業學院,昆明650607; 3.昆明醫科大學第二附屬醫院,昆明650101)
婚配制度是野生動物行為學領域的重要研究內容之一,達爾文首先從進化的角度討論了婚配制度,認為它是物種的一種基本進化策略(Darwin,1871)。在隨后的研究中,尤其是Emlen和Oring(1977)關于動物婚配的經典論著發表以后,婚配制度逐漸被視為動物在某一特定環境下為獲取配偶而采取的一種行為策略。經典的婚配制度常常根據動物的交配方式、獲得配偶方式和數量等特征進行劃分,主要有4種類型:單個雄性與單個雌性在幾個繁殖季節單獨婚配(一雄一雌制);一個雄性和幾個雌性在連續的幾個婚配季節婚配(一雄多雌制);單個雌性和多個雄性在連續的幾個婚配季節婚配(一雌多雄制);雌雄性之間沒有固定的婚配關系,彼此均有多個配偶(混交制)(Cluttonbrock,1989;Clawetal.,2018)。在研究動物的婚配類型時,需要注意行為學上的一雄一雌和遺傳學上的一雄一雌在哺乳動物中單獨進化現象。行為學上的單配制被理解為雌雄個體在空間組織上的親密配對關系,而遺傳學上的單配制則強調分子遺傳學層面的排他交配情況(Reichard & Christophe,2003;Ophir,2017)。其次,以Shuster和Wade為代表的學者強調在劃分動物婚配類型時,依然需要關注該物種所處棲息地的環境條件,同時,他們也提出多達10余種婚配類型(Shuster & Wade,2003)。截止目前,哺乳動物婚配制度的研究依然是一個活躍的領域。
Short(1979)提出了“精子競爭假說”,認為一雄多雌的動物,如大猩猩屬Gorilla的物種,一個雄性獨占多個雌性交配的權利,體型大,相對睪丸較小;相比之下,在黑猩猩Pantroglodytes中,幾個雄性常常與發情的雌性產生交配方面的競爭,在這類多雄婚配的物種中,睪丸會進化得更大。此外,他還認為較大的睪丸有利于產生更多的精子,以便在性競爭中獲得更大優勢。Harcourt等(1981)首次證實了在靈長類動物中,睪丸尺寸與婚配制度之間存在相關性,交配模式引發更高精子競爭水平的婚配類型(多雄)進化出的相對睪丸大小要比一雄一雌制的大。隨后,除了靈長類動物以外,這種相對睪丸大小與婚配類型的相關性在多個類群中得到了研究,如魚類、兩棲動物、鳥類、哺乳動物和昆蟲中都得到了一致的研究結果(Jolly & Conroy,2003;Fromhageetal.,2008;Manseretal.,2017;Phametal.,2017)。
中緬樹鼩Tupaiabelangeri是外形酷似松鼠的一類雜食性小型哺乳動物,屬攀鼩目Scandentia樹鼩科Tupaiidae,它是東南亞樹鼩中分布最北的一個種,國內主要分布于云南、貴州、四川、廣西和海南島等地(彭燕章等,1991)。近年來,無論是基因組學還是蛋白組學的研究結果都表明中緬樹鼩與靈長類具有較近的親緣關系(Lietal.,2012;Fanetal.,2013)。中緬樹鼩特殊的進化地位使其正逐漸成為生物醫學研究領域中的重要模型動物,揭示該物種的婚配制度對其人工馴化和提高室內繁殖成活率具有重要的生物學意義。野生中緬樹鼩常常是獨居的,具有領域性,雄性間對抗明顯,常常驅逐其他雄性入侵者(彭燕章等,1991)。野外很難觀察到中緬樹鼩的交配,關于該物種的婚配制度主要從領域性、雙親行為、配偶選擇等方面展開:人工飼養條件下,雄性和雌性緊密結合在一起,雌雄個體總是成對棲息于同一個巢箱里,并伴隨標記行為(Martin,1968)。在籠養時2個成年雄性之間的對抗極為明顯,而雌性間卻表現出容忍,并有相互修飾和理毛等行為,這似乎暗示中緬樹鼩是以單個雄性為主的社會結構(Sorenson,1974)。Kawamichi和Kawamichi(1982)用無線電項圈研究了中緬樹鼩的家域后發現,雄性中緬樹鼩的家域總是和一個或多個雌性重疊,這似乎也暗示了中緬樹鼩在自然條件下可能屬于一雄一雌制或一雄多雌制的繁殖方式。近年來,國內中緬樹鼩的馴化繁殖逐步加大,以中國科學院昆明動物研究所、中國醫學科學院醫學生物學研究所和廣西大學等為主的單位在中緬樹鼩的養殖中常常采用雌雄比例為1∶1和 1∶2的配種方式進行,能取得較高的繁殖率,而其他比例往往導致雌性的早產或流產的現象(江勤芳等,2011)。
20世紀80年代,在行為生態學領域,一些研究發現動物的性二態可以對其行為、生態及進化等問題做出某些推測或假說,如在野生動物中,性選擇被認為是直接導致性二態發生的原因,缺乏性二態往往被認為是一雄一雌制的典型特征(Leutenegger,1987)。中緬樹鼩的性二態研究也僅僅在毛色和體質量上進行。動物性二態的定性和定量的方式也不盡相同,對其內容和含義的理解也存在諸多爭議(吳汝康,1987)。同時,Collins(1987)也指出將性二態特征作為預測中緬樹鼩婚配類型之前,需要對其所處的棲息環境詳細了解,因為環境因子往往會對動物的婚配方式產生巨大的影響。綜上所述,中緬樹鼩的婚配制度主要存在一雄一雌制和一雄多雌制2種假設,野外棲息狀態下的中緬樹鼩究竟采取何種婚配方式?成年雄性中緬樹鼩是否具有精子競爭?婚配制度與睪丸尺寸之間是否具有相關性?為了解決這些問題,本研究在樹鼩的繁殖季節(3—5月),測定了雄性中緬樹鼩的睪丸質量和體質量,并與60種靈長類中不同婚配制度物種進行對比分析,探究中緬樹鼩的婚配制度。
1 材料和方法
1.1 不同婚配制度靈長類動物睪丸質量、體質量的數據來源
研究采用的不同婚配制度靈長類動物的睪丸質量和體質量數據詳見表1,包括一雄一雌、一雄多雌、一雌多雄3種婚配制度,體質量最小的倭狨Cebuellapygmaea到最大的西非大猩猩Gorillagorilla,共計34屬60種。
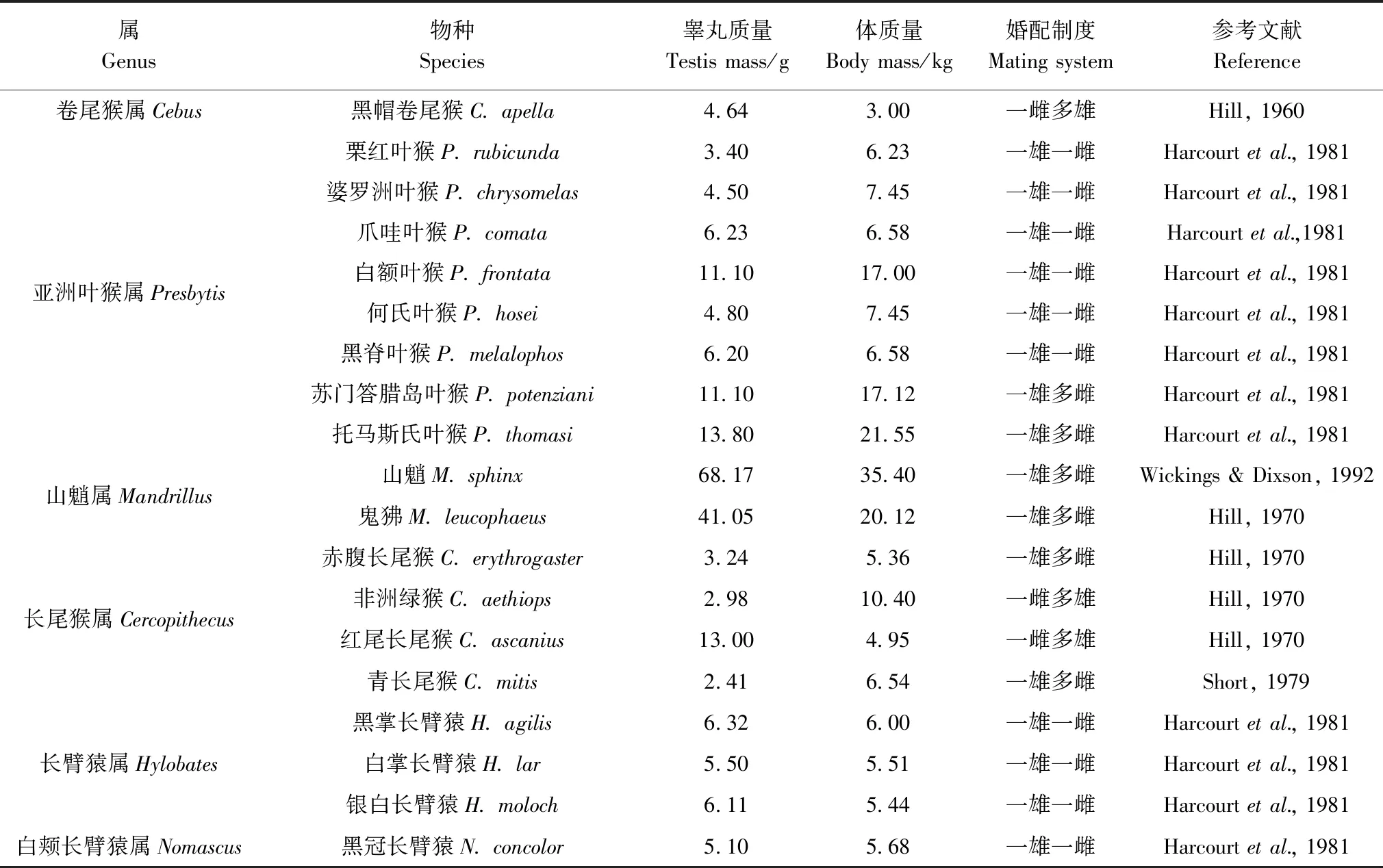
表1 不同靈長類動物及中緬樹鼩的睪丸質量和體質量Table 1 Testis mass and body mass of different primates and Tupaia belangeri
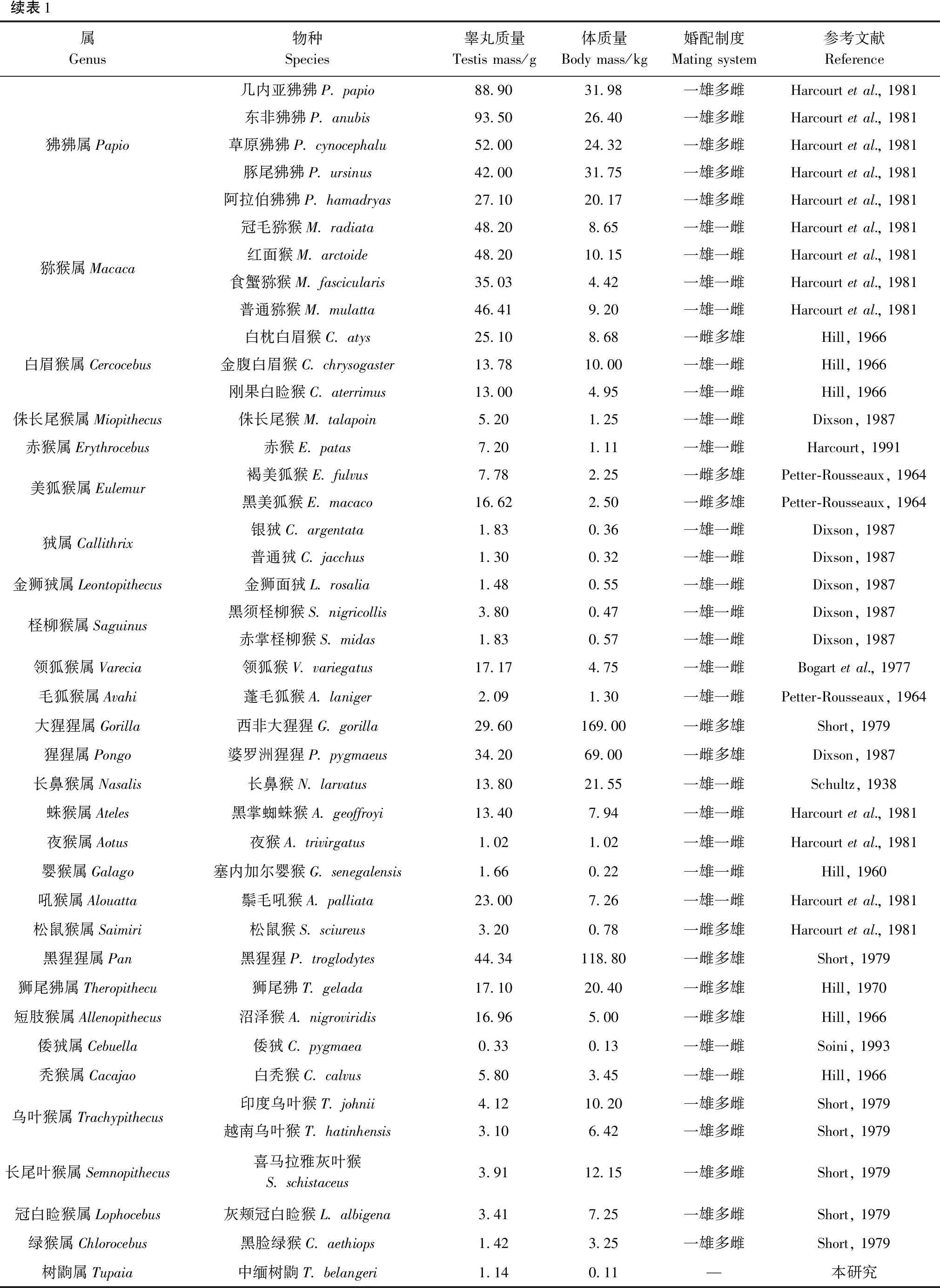
續表1屬Genus物種Species睪丸質量Testismass/g體質量Bodymass/kg婚配制度Matingsystem參考文獻Reference狒狒屬Papio幾內亞狒狒P.papio88.9031.98一雄多雌Harcourtetal.,1981東非狒狒P.anubis93.5026.40一雄多雌Harcourtetal.,1981草原狒狒P.cynocephalu52.0024.32一雄多雌Harcourtetal.,1981豚尾狒狒P.ursinus42.0031.75一雄多雌Harcourtetal.,1981阿拉伯狒狒P.hamadryas27.1020.17一雄多雌Harcourtetal.,1981獼猴屬Macaca冠毛獼猴M.radiata48.208.65一雄一雌Harcourtetal.,1981紅面猴M.arctoide48.2010.15一雄一雌Harcourtetal.,1981食蟹獼猴M.fascicularis35.034.42一雄一雌Harcourtetal.,1981普通獼猴M.mulatta46.419.20一雄一雌Harcourtetal.,1981白眉猴屬Cercocebus白枕白眉猴C.atys25.108.68一雌多雄Hill,1966金腹白眉猴C.chrysogaster13.7810.00一雄一雌Hill,1966剛果白瞼猴C.aterrimus13.004.95一雄一雌Hill,1966侏長尾猴屬Miopithecus侏長尾猴M.talapoin5.201.25一雄一雌Dixson,1987赤猴屬Erythrocebus赤猴E.patas7.201.11一雄一雌Harcourt,1991美狐猴屬Eulemur褐美狐猴E.fulvus7.782.25一雌多雄Petter-Rousseaux,1964黑美狐猴E.macaco16.622.50一雌多雄Petter-Rousseaux,1964狨屬Callithrix銀狨C.argentata1.830.36一雄一雌Dixson,1987普通狨C.jacchus1.300.32一雄一雌Dixson,1987金獅狨屬Leontopithecus金獅面狨L.rosalia1.480.55一雄一雌Dixson,1987檉柳猴屬Saguinus黑須檉柳猴S.nigricollis3.800.47一雄一雌Dixson,1987赤掌檉柳猴S.midas1.830.57一雄一雌Dixson,1987領狐猴屬Varecia領狐猴V.variegatus17.174.75一雄一雌Bogartetal.,1977毛狐猴屬Avahi蓬毛狐猴A.laniger2.091.30一雄一雌Petter-Rousseaux,1964大猩猩屬Gorilla西非大猩猩G.gorilla29.60169.00一雌多雄Short,1979猩猩屬Pongo婆羅洲猩猩P.pygmaeus34.2069.00一雌多雄Dixson,1987長鼻猴屬Nasalis長鼻猴N.larvatus13.8021.55一雄一雌Schultz,1938蛛猴屬Ateles黑掌蜘蛛猴A.geoffroyi13.407.94一雄一雌Harcourtetal.,1981夜猴屬Aotus夜猴A.trivirgatus1.021.02一雄一雌Harcourtetal.,1981嬰猴屬Galago塞內加爾嬰猴G.senegalensis1.660.22一雄一雌Hill,1960吼猴屬Alouatta鬃毛吼猴A.palliata23.007.26一雄一雌Harcourtetal.,1981松鼠猴屬Saimiri松鼠猴S.sciureus3.200.78一雌多雄Harcourtetal.,1981黑猩猩屬Pan黑猩猩P.troglodytes44.34118.80一雌多雄Short,1979獅尾狒屬Theropithecu獅尾狒T.gelada17.1020.40一雌多雄Hill,1970短肢猴屬Allenopithecus沼澤猴A.nigroviridis16.965.00一雌多雄Hill,1966倭狨屬Cebuella倭狨C.pygmaea0.330.13一雄一雌Soini,1993禿猴屬Cacajao白禿猴C.calvus5.803.45一雄一雌Hill,1966烏葉猴屬Trachypithecus印度烏葉猴T.johnii4.1210.20一雄多雌Short,1979越南烏葉猴T.hatinhensis3.106.42一雄多雌Short,1979長尾葉猴屬Semnopithecus喜馬拉雅灰葉猴S.schistaceus3.9112.15一雄多雌Short,1979冠白瞼猴屬Lophocebus灰頰冠白瞼猴L.albigena3.417.25一雄多雌Short,1979綠猴屬Chlorocebus黑臉綠猴C.aethiops1.423.25一雄多雌Short,1979樹鼩屬Tupaia中緬樹鼩T.belangeri1.140.11—本研究
1.2 雄性中緬樹鼩睪丸和體質量的測定
研究地點位于云南省昆明市以西50 km處的西山區海口林場,所用的38只中緬樹鼩均為野外捕捉:2018年3—5月,在獲得林場單位批準后,采用以蘋果為誘餌的金屬網格捕鼠籠放置在中緬樹鼩常出現的灌木叢下或倒下的樹干旁,周圍撒上誘餌。每隔1~2 h檢查一次陷阱,一旦抓獲,保留健康成年雄性中緬樹鼩,記錄基本信息(體質量、捕獲地點),并將其轉移到布織袋中稱重帶回實驗室,斷頸處死,解剖其睪丸,采用電子天平(上海精密科學儀器有限公司)測定睪丸質量,精確到0.01 g。
1.3 數據處理
2 研究結果
采用已知婚配制度的60種靈長類動物與中緬樹鼩的相對睪丸大小與體質量的對比研究發現,靈長類動物交配系統的進化與相對睪丸大小的進化有顯著的相關性(圖1),一雄一雌制物種的相對睪丸大小小于一雌多雄制和一雄多雌制。本研究中,體質量較大的靈長類物種的睪丸質量也較大(r=0.31,P<0.05),睪丸質量在3種婚配類型之間的差異有高度統計學意義(F2,57=11.36,P<0.01),其中,一雄多雌制物種的睪丸質量最大,而一雄一雌制的最輕。體質量與婚配制度之間的差異有統計學意義(F2,57=4.09,P<0.05),雄性中緬樹鼩體質量為113.29 g±9.07 g,睪丸質量為1.14 g±0.25 g(表2)。
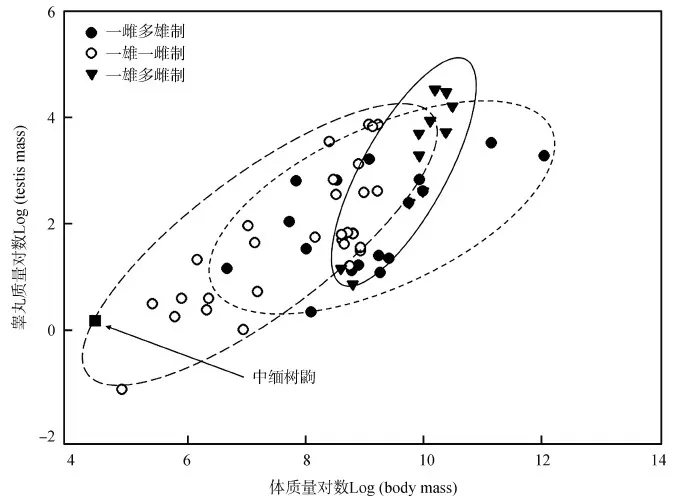
圖1 中緬樹鼩和60種靈長類動物睪丸質量對數與體質量對數的關系Fig. 1 Relationship between log testis mass and log body mass in Tupaia belangeri and 60 primate species
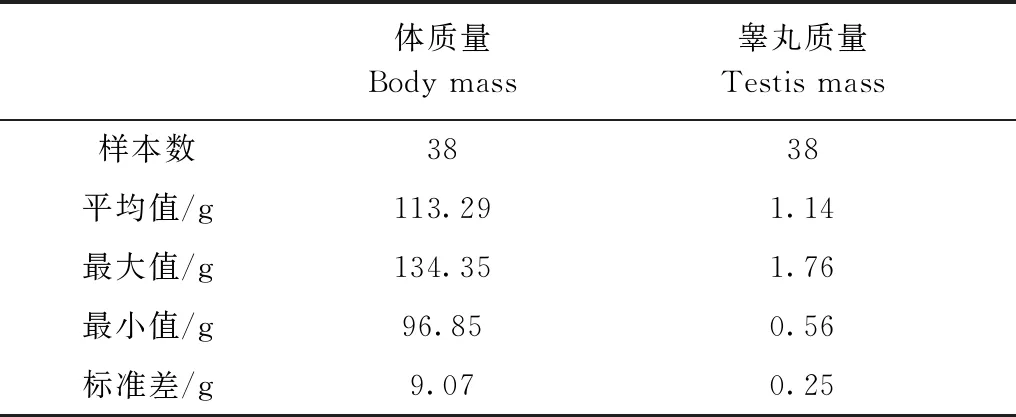
表2 雄性中緬樹鼩體質量和睪丸質量Table 2 Descriptive statistics of body mass and testis mass of male Tupaia belangeri
3 討論
雄性哺乳動物的睪丸由曲細精管與間質細胞組成,其尺寸與性行為和婚配方式息息相關,主要功能有:制造精子、生成雄性激素、吸引異性等(Vellnowetal.,2018)。精子是一類在復雜環境中以微觀水平發揮作用的特化細胞群,同一類群的哺乳動物相對睪丸大小差異是在不同婚配方式和精子競爭的水平上發展起來的(Gage & Freckleton,2003;Rowleyetal.,2019)。在雄性生殖特征中,睪丸的相對大小呈現出與精子競爭水平的一致性。不同的交配策略和雄性對雌性的壟斷情況往往可以通過一系列的行為和形態適應所反映出來(Lynch & Michael,1991)。研究表明,一雄多雌制和單配制動物在睪丸大小上區分尤為明顯,因為單配制物種的睪丸尺寸相對較小,精子競爭水平較低(Short,1997)。其次,雄性在野外往往只壟斷1只雌性,無需進化出龐大的體型或者特殊性狀來壟斷交配。近年來,有學者又在西非大猩猩、黑猩猩和紅毛猩猩Pongopygmmaeus睪丸的顯微結構和相對精子產生的比較研究中提出一種更能反映精子競爭的新指標——生精指數,該指標與精子產生密切相關,也是預測和評估哺乳動物婚配類型的一個實用性指標(Fujii-Hanamotoetal.,2011)。由此可見,睪丸這一伴隨著哺乳動物的出現而進化出來的性器官,其尺寸的進化和交配方式及精子的競爭水平息息相關,當來自不同雄性的精子爭奪受精權時,精子和睪丸的競爭就存在了,它們被視為一種強大的進化力量,最終導致了一系列豐富的性選擇適應(Kahrletal.,2019)。
野生哺乳動物的睪丸相對大小和交配系統之間具有密切的關系,使得睪丸的解剖學證據可在大體上預測或確認未知動物的婚配方式(Fietz,1999)。本研究中60種靈長類動物的體質量和睪丸質量的數據與中緬樹鼩的對比分析表明,睪丸尺寸與不同婚配類型存在相關性,一雄多雌制的靈長類動物的睪丸尺寸要比一雌多雄制和一雄一雌制的靈長類動物要大,與部分研究結果一致(Harcourtetal.,1995;Kappeler,1997)。本文首次以睪丸質量作為依據,對野生中緬樹鼩的相對睪丸大小和婚配類型之間的關系進行了探究,較小的睪丸相對比表明中緬樹鼩的精子競爭是微弱的,推測中緬樹鼩的婚配制度可能為一雄一雌制。值得注意的是,近些年,來自DNA指紋、微衛星DNA等方面的分子遺傳學證據逐漸揭示出動物的婚配制度比以往所認為的更加復雜和多樣化,為了更好地確認該物種的婚配制度,未來可結合DNA分子技術尋找野外中緬樹鼩個體間的親緣關系來完成。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
遼金歷史與考古(2019年0期)2020-01-06 07:44:44
中國生殖健康(2019年2期)2019-08-23 08:12:08
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國衛生(2016年7期)2016-11-13 01:06:26
中國衛生(2016年11期)2016-11-12 13:29:18
中國衛生(2016年9期)2016-11-12 13:27:58
