東亞三角渦蟲DjPLCβ4基因克隆及在胚胎發(fā)育中的組織特異性表達
2020-06-11 11:08:32甄輝趙博生
四川動物 2020年3期
關鍵詞:小鼠
甄輝, 趙博生
(山東理工大學發(fā)育與進化生物學研究所,山東淄博255000)
哺乳動物中,磷脂酶C(PLC)根據(jù)序列同源性和結(jié)構域的位置,可分為β、γ和δ三大類(Rhee,2001;Fukami,2002;Hwangetal.,2005;Nakaharaetal.,2005;Zhouetal.,2005),是一種存在于細胞膜上的關鍵酶。在大腦中,PLC-β家族的表達譜比較清楚,PLC-β1、β3和β4是最常見的酶型且有其獨特的表達模式。PLC-β1在大腦皮層和海馬中高表達,在小腦中表達較少(Rossetal.,1989);PLC-β4在小腦中表達最高,在大腦皮層和海馬中的表達量極低,而PLC-β3在整個腦中的表達量都很低(Tanaka & Kondo,1994)。
PLCβ4基因的異常表達與癲癇、精神分裂癥、亨廷頓病、抑郁癥等腦部疾病相關(Yangetal.,2016)。研究結(jié)果表明,PLCβ4基因缺陷小鼠Musmusculus的視覺系統(tǒng)會出現(xiàn)功能障礙,PLCβ4在視桿外部的初始光層疊后的視覺處理中起到重要作用(Jiangetal.,1996)。PLCβ4基因突變小鼠表現(xiàn)出運動障礙和運動性共濟失調(diào)的癥狀,尤其是后肢更嚴重(Kimetal.,1997)。PLCβ4還與生物鐘功能相關,研究發(fā)現(xiàn),在小鼠的肝臟中,PLCβ4基因和蛋白的表達受晝夜節(jié)律的調(diào)控,而不依賴于外部光周期(Kleinetal.,2008)。PLCβ4在睡眠-覺醒和體溫節(jié)律中也發(fā)揮著重要的作用。
神經(jīng)系統(tǒng)是后生動物中最復雜的器官系統(tǒng)之一。神經(jīng)網(wǎng)絡調(diào)節(jié)動物生理學和控制行為等方面。因此,了解神經(jīng)系統(tǒng)如何在整個生命中發(fā)育、功能和重塑是一個有趣且具有挑戰(zhàn)性的研究熱點。與線蟲或果蠅相比,渦蟲因其強大再生能力而備受關注(Baguna,2012;Rink,2013)。此外,渦蟲是擁有前腦部化(“大腦”)的最原始生物,這很可能代表人類大腦的祖先(Sarnat & Netsky,1985,2002)。東亞三角渦蟲Dugesiajaponica屬扁形動物門Platyhelminthes渦蟲綱Turbellaria三腸目Tricladida(劉德增,1989),有2種繁殖方式:有性生殖和無性生殖。無性生殖是指渦蟲本體受到外界或自身條件影響而斷裂,并再生出完整個體。有性生殖會產(chǎn)生具有多個受精卵和外卵黃細胞組成的卵囊,胚胎在卵囊內(nèi)發(fā)育,發(fā)育過程分為8個階段(Yuanetal.,2010,2014;甄輝等,2016)。每個卵囊里含有1~10個處于不同發(fā)育階段的受精卵,受精卵的周圍充滿了大量的外卵黃,其在卵囊中發(fā)育直至幼蟲(Hartenstein & Ehlers,2000)。
為了研究PLCβ4在渦蟲胚胎發(fā)育中的表達譜,本文通過RT-PCR和RACE技術克隆得到東亞三角渦蟲PLCβ4基因,并利用整體原位雜交技術研究了其在渦蟲胚胎發(fā)育中的時空表達圖譜。
1 材料與方法
1.1 實驗材料
東亞三角渦蟲胚胎采自山東省淄博市博山泉河頭;成體為實驗室培養(yǎng)單系,飼養(yǎng)在魯山山泉水中。胚胎經(jīng)DEPC水沖洗后,用解剖針扎破,4%多聚甲醛室溫固定30 min,30%、50%、75%甲醇梯度脫水,100%甲醇-20 ℃保存待用。實驗前成體饑餓1周。胚胎每組實驗樣本量為8個,成體實驗樣本量為10條。PMD18-T載體為TaKaRa,EscherichiacoliDH5α為實驗室保存。
1.2 引物設計與PCR擴增
引物設計PLCβ4(F和R)(表1)和PCR步驟參照甄輝等(2016)。利用TRIZOL試劑提取渦蟲總RNA,測量RNA濃度,利用Fermentas反轉(zhuǎn)錄試劑盒合成cDNA模板。由上海Sangon公司合成。以渦蟲cDNA為模版,利用F和R引物進行PCR擴增,反應程序:94 ℃預變性5 min;94 ℃變性50 s,57 ℃退火60 s,72 ℃延伸60 s,共30個循環(huán);72 ℃延伸10 min。PCR擴增結(jié)束后,利用OMEGA膠回收試劑盒回收。將純化回收的PCR產(chǎn)物連接到PMD18-T載體上送上海博尚生物技術有限公司測序。
1.3 通過RACE技術獲得PLCβ4全長
根據(jù)測序獲得的部分PLCβ4序列設計RACE 5’和3’的引物(表1)。RACE-PCR反應程序參照甄輝等(2016)。5’-和3’-cDNA模板利用Clontech BD SMARTAM RACE cDNA Amplification Kit反轉(zhuǎn)錄試劑盒反轉(zhuǎn)錄。RACE-PCR的循環(huán)程序如下:94 ℃預變性2 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸45 s,共35個循環(huán);72 ℃延伸2 min。PCR擴增產(chǎn)物通過膠回收純化,并連接到PMD18-T載體上送上海博尚生物技術有限公司測序。
1.4 生物信息學分析
生物信息學分析的軟件和步驟參照甄輝等(2016)。序列用DNAtools進行蛋白質(zhì)序列翻譯,在NCBI中BLASTP(http://blast.ncbi.nlm.nih.gov),尋找存在的同源基因。用DNASTAR中的MegAlign分析同源蛋白質(zhì)序列之間的相似性,利用MEGA 7的最大似然法對其進行進化樹分析,重復1 000次。
1.5 DIG-Labeling RNA探針合成
根據(jù)測序獲得的基因序列設計引物(表1)。首先,利用PCR獲得3’端有T7啟動子的PCR產(chǎn)物,沉降純化回收后連接到PMD18-T載體上送上海博尚生物技術有限公司測序。DIG-Labeling RNA探針合成體系:模板(3’端帶有T7啟動子的PCR產(chǎn)物1 mg),T7 RNA polymerase(Promega,Madison,美國),DTT(Promega,Madison,美國),DIG-Labeling RNA Mix(Roche)和RNAse Inhibitor(Fermentas,Waltham,美國),37 ℃孵育12 h。
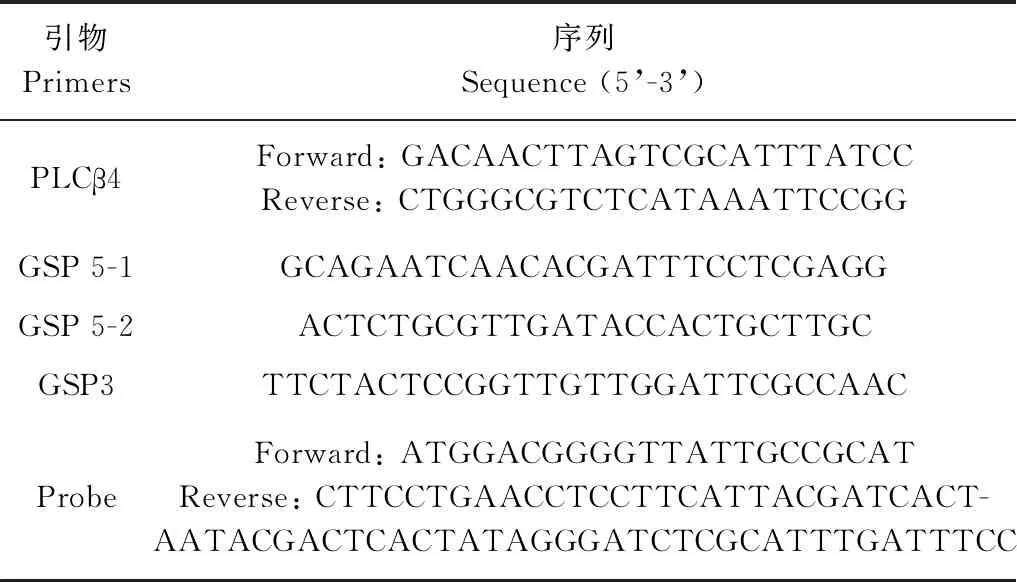
表1 實驗引物設計Table 1 Primers used in this study
1.6 原位雜交
原位雜交參照甄輝等(2016)和Zhen等(2019)的方法。胚胎固定時經(jīng)DEPC水沖洗后,用5 mm解剖針扎破,幼蟲經(jīng)5%NAC處理5 min,4%多聚甲醛室溫固定30 min,30%、50%、75%甲醇梯度脫水,100%甲醇-20 ℃保存待用或直接用于原位雜交。原位雜交時甲醇(75%、50%、30%)梯度復水;10 μg·mL-1Proteinase K室溫下消化10 min;4%多聚甲醛室溫固定30 min;0.1 mol·L-1三乙醇胺加0.25%乙酸酐進行乙酰化;56 ℃預雜交2 h;雜交時加入1 μg·mL-1RNA探針,56 ℃雜交16 h。雜交后洗滌,10%馬血清溶液封閉2 h;anti-Dig antibody (1∶2 000),4 ℃孵育過夜;室溫條件下,馬來酸緩沖液清洗8次,AP Buffer孵育10 min;加入5-bromo-4-chloro-3-indolylphosphate和nitroblue tetrazolium(BCIP/NBT;Roche)顯色液后,放置暗處顯色;在合適信噪比時終止反應,用Nikon AZ100體視顯微鏡進行拍照。
2 結(jié)果
2.1 基因克隆
利用RT-PCR引物擴增的PCR產(chǎn)物大小為1 084 bp;利用5’-RACE引物GSP-1、GSP-2擴增的PCR產(chǎn)物大小分別為1 246 bp和963 bp,利用3’-RACE引物GSP3擴增的PCR產(chǎn)物大小為418 bp(圖1:A)。將測序獲得序列比對拼接,結(jié)果顯示,從東亞三角渦蟲中克隆得到的DjPLCβ4基因全長為3 245 bp,最大開放閱讀框為2 907 bp,編碼969個氨基酸組成的蛋白質(zhì),該蛋白質(zhì)的預測分子量約為110.55 kDa,5’非翻譯區(qū)為42 bp,3’非翻譯區(qū)為296 bp,5’端有1個起始密碼子,3’非翻譯區(qū)包含多聚腺苷酸化信號位點和poly(A)尾,由此認為,克隆得到的cDNA為全長序列,GenBank登錄號詳見表2。
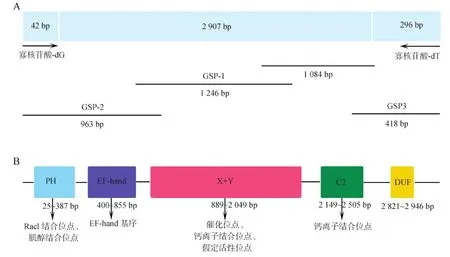
圖1 DjPLCβ4基因獲得的引物位置(A)和結(jié)構域(B)Fig. 1 Schematic representation of the amplified gene region (A) and structure (B) of DjPLCβ4 gene

表2 本研究所用物種PLCβ4基因序列的GenBank登錄號Table 2 GenBank accession numbers of PLCβ4 genes used in this study
2.2 生物信息學分析
在NCBI BLASTP(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析獲得的氨基酸序列,該氨基酸包含PH結(jié)構域、EF-hand結(jié)構域、X+Y結(jié)構域、C2結(jié)構域和DUF結(jié)構域(圖1:B)。相似性結(jié)果顯示,與其他物種PLCβ4蛋白相似性在36%以上,與選取的脊椎動物PLCβ4蛋白相似性為55%。選擇6種代表性物種,將不同物種的PLCβ4蛋白的不同結(jié)構域用MegAlign比對,結(jié)果表明,DjPLCβ4是一個非常保守的蛋白質(zhì),與其他物種的PH結(jié)構域、EF-hand結(jié)構域、X+Y結(jié)構域、C2結(jié)構域的相似性很高。為了進一步驗證DjPLCβ4的進化地位,選擇了7種代表性物種的PLC[β(1~4)、γ(1,2)、δ(1,3,4)、η(1,2)、ζ1、ε1]蛋白氨基酸序列,MEGA 7構建的進化樹顯示,DjPLCβ4與其他物種PLCβ4聚為一支,并位于兩側(cè)對稱動物的基部(紅色方框),符合進化地位,進一步驗證了DjPLCβ4是PLCβ4的一員(圖2)。因此,將其命名為DjPLCβ4。
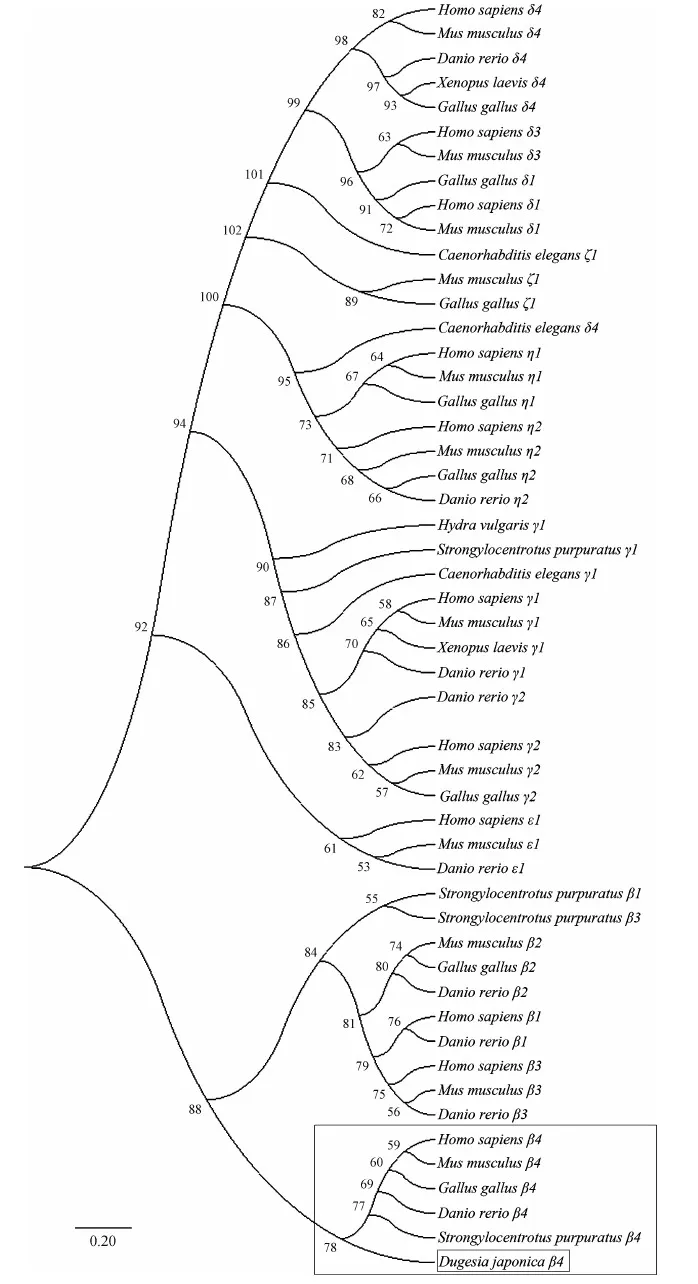
圖2 MAGE 7(最大似然法)構建DjPLCβ4的系統(tǒng)進化樹(bootstrap=1 000)
Fig. 2 Phylogenic tree of DjPLCβ4 constructed by MAGE 7 (Maximum likelihood tree)(bootstrap=1 000)
2.3 原位雜交
為探索DjPLCβ4基因在渦蟲胚胎發(fā)育中的表達圖譜,利用原位雜交技術檢測DjPLCβ4基因在不同胚胎時期的表達圖譜。結(jié)果顯示,第1、2階段(胚胎卵裂期)沒有檢測到DjPLCβ4基因的陽性信號(圖3:B);第3階段,胚胎開始分化,形成胚胎咽,外卵黃被胚胎咽吸收,隨著外卵黃不斷變大,合胞體被擠到最外層,并形成含有胚胎細胞的胚帶,DjPLCβ4基因主要表達于胚胎咽和胚帶(圖3:C)。第4階段,外卵黃繼續(xù)被胚胎咽吸入,胚胎體積達到最大并開始變形,DjPLCβ4基因主要表達于神經(jīng)原基、胚胎咽及胚帶(圖3:D);第5階段,合胞體出現(xiàn)解體,胚胎逐漸變?yōu)楸鈭A形,細胞和組織開始分化形成表皮細胞和肌肉細胞,DjPLCβ4基因主要表達于咽原基和神經(jīng)原基(圖3:E);第6階段,胚胎不斷變大,大量細胞出現(xiàn)分化,腹神經(jīng)更加明顯,且第一個器官咽形成,DjPLCβ4基因主要表達于咽和神經(jīng)系統(tǒng),且頭部信號較明顯(圖3:F);第7、8階段,胚胎大腦形成,神經(jīng)系統(tǒng)進一步成熟,胚胎邊緣逐漸形成原始的神經(jīng)系統(tǒng),DjPLCβ4基因主要表達于神經(jīng)系統(tǒng)(圖3:G)。
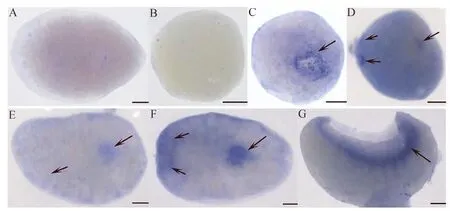
圖3 DjPLCβ4基因在東亞三角渦蟲胚胎發(fā)育中的表達Fig. 3 Expression of DjPLCβ4 gene by whole-mount in situ hybridization during embryonic development of Dugesia japonica
A. 對照組, B. 第1和2階段, C. 第3階段, D. 第4階段, E. 第5階段, F. 第6階段, G. 第7和8階段; 比例尺=100 μm
A. control, B. stages 1 and 2, C. stage 3, D. stage 4, E. stage 5, F. stage 6, G. stages 7 and 8; scale bars=100 μm
隨后,胚胎發(fā)育成熟,孵化出幼蟲,1周后長成成體。利用整體原位雜交技術檢測到DjPLCβ4基因主要表達于成體的腦和咽(圖4:B)。
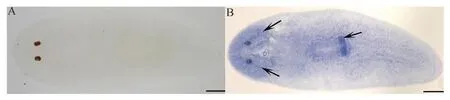
圖4 DjPLCβ4基因在東亞三角渦蟲整體中的表達圖式Fig. 4 Expression analysis of DjPLCβ4 gene in the intact planarian
A. 對照組, B.DjPLCβ4基因表達于腦和咽(箭頭); 比例尺=500 μm
A. control, B.DjPLCβ4gene expressed in the brain and pharynx (arrows); scale bars=500 μm
3 討論
磷脂酰肌醇代謝是一個重要的細胞內(nèi)信號系統(tǒng),參與多種細胞功能,包括激素分泌、神經(jīng)遞質(zhì)傳遞、生長因子信號、細胞膜運輸和細胞骨架調(diào)節(jié)等(Janetopoulos & Devreotes,2006;Santariusetal.,2006;Cockcroft & Carvou,2007)。磷脂酰肌醇特異性磷脂酶C是該系統(tǒng)中的關鍵酶,水解磷脂酰肌醇4,5-二磷酸[PI(4,5)P2]生成肌醇1,4,5-三磷酸(IP3)和二酰甘油(DAG)2個第二信使,以響應激素、神經(jīng)遞質(zhì)、生長因子激活受體和其他分子(Nakamura & Fukami,2009)。從哺乳動物中克隆出13種PLC同工酶,根據(jù)結(jié)構和調(diào)控激活機制可分為以下6類:PLC-δ(1,3,4)、PLC-β(1~4)、PLC-γ(1,2)、PLC-ε、PLC-ζ、PLC-η(1,2)(Rhee,2001;Fukami,2002;Hwangetal.,2005;Nakaharaetal.,2005;Zhouetal.,2005)。每一種同工酶都由亞型特異的保守結(jié)構域組成。所有的PLC同工酶都含有催化的X+Y結(jié)構域以及各種調(diào)控結(jié)構域,包括C2結(jié)構域、EF-hand結(jié)構域和PH結(jié)構域。在東亞三角渦蟲中獲得DjPLCβ4基因,利用NCBI BLASTP (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)對編碼的氨基酸序列進行分析,發(fā)現(xiàn)該氨基酸包含PH結(jié)構域、EF-hand結(jié)構域、X+Y結(jié)構域和C2結(jié)構域,這與上述文獻結(jié)果相同。進化分析結(jié)果顯示,DjPLCβ4與選取物種的PLCβ4聚為一支,并位于兩側(cè)對稱動物的基部,符合進化地位,進一步證明DjPLCβ4是PLCβ4的一員。
PLCs在器官形成和胚胎發(fā)育中發(fā)揮著重要的作用。研究表明,PLCβ1、β3、γ1和γ2在小鼠卵細胞中表達(Dupontetal.,1996;Mehlmannetal.,1998;Wangetal.,1998);在精子中檢測到β1~3、γ1,2、δ2、δ4和ζ的表達(Mehlmannetal.,1998;Fukamietal.,2001;Wuetal.,2001;Albaetal.,2002;Parringtonetal.,2002)。受精和卵子激活是胚胎發(fā)育的第一步,研究發(fā)現(xiàn),PLC-δ4和PLC-γ與這一現(xiàn)象密切相關,PLC-δ4基因敲除小鼠表現(xiàn)出雄性不育,且體外受精實驗發(fā)現(xiàn),這些動物的精子僅能夠誘導較少卵子的激活,并且不會引起鈣振蕩(Fukamietal.,2001)。由于鈣振蕩增加是啟動卵子激活的關鍵,而PLC-δ4是精子中誘導鈣振蕩和卵子激活的重要基因(Wassarman,1999)。PLC-β的分布非常廣,在神經(jīng)系統(tǒng)、感覺器官及腺體的表達量顯著高于其他位置。成熟小鼠卵細胞在mRNA水平上表達2種PLCβ同工酶β1和β3(Dupontetal.,1996)。PLCβ3缺陷的胚胎在第4細胞期前死亡,表明該酶參與卵子激活或早期胚胎發(fā)育(Wangetal.,1998)。在成年小鼠中,PLC-β4在蒲肯野細胞、丘腦、中隔及眼睛的視桿細胞有高表達(Watanabeetal.,1998)。小腦皮層中層的蒲肯野細胞中,PLC-β4高表達,敲除后小鼠會出現(xiàn)運動失調(diào)(Kimetal.,1997)。而小鼠丘腦皮質(zhì)PLCβ4缺失則會導致其失神發(fā)作(Cheongetal.,2009)。PLCβ4與小腦發(fā)育過程中的纖維突觸清除有關(Kanoetal.,1998)。PLCβ4在大腦發(fā)育和發(fā)揮正常功能中起著不可替代的作用。研究還發(fā)現(xiàn),丘腦mGluR1-PLCβ4通路是控制睡眠結(jié)構的關鍵(Hongetal.,2016)。在渦蟲胚胎發(fā)育的第3階段檢測到DjPLCβ4基因在胚胎咽出現(xiàn)信號,之后在分化出的神經(jīng)原基和咽原基中也檢測到陽性信號,隨著胚胎腹部神經(jīng)加厚并豐富,陽性信號增多,且頭部更加明顯。DjPLCβ4基因主要集中在成體的腦和咽。因此,DjPLCβ4基因在渦蟲胚胎發(fā)育時主要參與渦蟲神經(jīng)原基的形成和分化。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
