南寧老虎嶺林區多種外來植物入侵程度與影響因子模型解釋
2020-06-08 00:43:12黃小榮
植物研究 2020年3期
黃小榮
(廣西壯族自治區林業科學研究院,南寧 530002)
外來物種(alien species)是指人類有意或無意地在其過去或現在自然分布之外引入的物種[1~2]。隨著人們對入侵物種、入侵影響和入侵控制的日益重視[3],探索外來物種入侵機理成為當前生態學研究熱點[4~5];通常用與入侵有關的環境因子來解釋入侵機理,包括道路條件、林冠郁蔽度、林下植被高度和鄉土物種豐富度等因素。道路是外來入侵物種傳播的主要途徑之一[6],道路修建干擾生態環境使路邊更容易被外來植物拓殖[7],植物種子附著在車輪上可沿路傳播數米至數千米[8~9]。對于森林群落,林冠郁蔽度和林下植被高度可能影響外來入侵,光照條件較差限制喜光入侵植物的種子萌發和生長繁殖,林下植被高度可能阻礙風傳型植物種子落地生根。土著物種豐富度與外來入侵的關系存在一定的爭議,在不同生境可能出現正相關、負相關或不相關[10]。自然群落通常存在多個外來種和多種環境因子,但大多數生物入侵研究只注重單一物種入侵[11],或只考慮單個因子和主效應;現實生態系統過程的復雜性,導致至關重要的生態效應往往不是主效應,而是與其他效應的互作[11]。為了有效地管理外來物種,在眾多外來物種持續存在的情況下,我們必須了解多個外來物種與多個影響因子之間的互作效應。
廣西具有沿邊沿海地理位置和溫暖濕潤的氣候,是外來植物入侵重災區。老虎嶺位于廣西南寧市北郊,近些年各種基因收集種質資源庫的建立、林區水泥路的鋪設、附近苗圃育苗、養蜂等活動,為外來植物的傳播、生長和繁衍提供了機會,外來植物的種群數量和分布范圍持續增加,改變了原有的植被風貌。老虎嶺林區分布有各種林齡、樹種和撫育時間的林分,為比較外來植物入侵與林區各種環境因子的關聯性提供了天然的條件。本研究在老虎嶺進行了分區樣方調查,擬比較不同林型、林齡、撫育時間、路邊與林內的外來植物入侵程度,探討多種因素對入侵的影響,為多因素對多個外來植物入侵的互作效應分析提供一個具體而詳細的范例。
1 材料和方法
1.1 研究區概況
研究區在老虎嶺林區一、二林班,調查樣地南起廣西林科院樹木園,北至老虎嶺水庫西北部的珍貴樹種任豆資源庫,南北、東西相距均為1 600 m,中心坐標108°20.4′E,22°55.8′N,海拔135~217 m。研究區地處南亞熱帶季風氣候區,年均降雨量1 304 mm。大部分土壤為砂頁巖發育而成的磚紅壤性紅壤,低山丘陵地貌。廣西林科院自1956年建院以來,在老虎嶺陸續營建了松杉人工林、引種樹木園以及加勒比松(Pinuscaribaea)、濕地松(P.elliottii)等國外松資源林;至1974年,老虎嶺的栽培喬灌木達684種。1995年在高速路橋邊原松樹皆伐跡地建立了馬尾松(Pinusmassoniana)優質種源林,2016年底進行了疏伐、砍灌和補植大葉櫟。2009~2015年,在老虎嶺水庫西南部、西部和西北部的松杉林皆伐跡地上分別營建了斑皮桉(Eucalyptusmaculata)、鄧恩桉(E.dunnii)和珍貴樹種基因庫;2016年將該區的土道鋪筑成水泥路;林分的撫育工作外包給同一包工隊,在每年1~2月砍灌割草。老虎嶺林區樹木基本連片,相對僻靜和冷清,除造林和撫育施工、苗圃育苗和養蜂之外其他干擾甚少。
1.2 樣地設置和測量
老虎嶺林區的外來物種分6個區(A~F)進行樣方調查,區間間隔200 m以上。每個區沿樣線調查20個路邊小樣方和20個林內小樣方,A、C和F區在道路一側、樣線長200 m,B、D和E區在道路兩側、樣線長100 m;林內樣線在距林緣滴水線10 m處設置;沿樣線間隔10 m取一個小樣方,尺寸均為1 m×1 m;每個區的第一個小樣方隨機起點,隨后樣方等距間隔,不因人為偏好而移位[12]。6個區共240個小樣方,分為15個組:路邊A、路邊B、路邊C、路邊D、路邊E、路邊F、A區馬尾松、B區紅錐(Castanopsishystrix)、B區尾巨桉(Eucalyptusurophylla×E.grandis)、C區鄧恩桉、D區西樺(Betulaalnoides)、D區任豆(Zeniainsignis)、E區麻櫟(Quercusacutissima)、E區國外松和F區斑皮桉。樣方調查在2018年6~7月進行,記錄每個小樣方內植物名和蓋度,以小樣方中心為圓心的直徑5 m樣圓記錄高度>3 m的林冠郁蔽度,分3個類型記錄小樣方位置高于林道、齊平林道或低于林道。
1.3 外來植物入侵的定量
為了避免主觀地劃分外來種和入侵種,本文的外來物種定義包括林下所有外來植物[6]。根據萬方浩[13]、林建勇[14~15]、唐賽春[16]等編目的名錄確定哪些是外來植物,包括林下高度≤5 m的非禾本科草本、禾本科草本、灌木以及藤本植物;不包括人工挖坎種植的引進樹種;以小樣方中所有外來植物占小樣方植被總蓋度的百分比作為外來植物占比(alien ratio),代表外來植物入侵程度(invasion)。
1.4 其他因子的獲取
咨詢相關人員和查閱廣西林科院內部資料,確定各林分的林齡。將撫育時間分為4個等級:之前4~6個月撫育(M1)、之前6~20個月撫育(M2)、之前20~36個月撫育(M3)、之前36個月以上無撫育(M4)。根據調查數據確定每種植物的最大株高。以小樣方的土著物種個數作為土著物種豐富度。將蓋度0~1%、1%~5%、5%~25%、25%~50%、50%~75%、75%~95%、95%~100%轉換成1~7級多度[17],用R-FD包計算每個小樣方的多度加權優勢最大株高,它是樣方內林下植被潛在最大株高與多度乘積的加和[18]。
1.5 數據分析
用SPSS 24.0的圖表構建器進行數據的初步探索,有意義的關聯使用均值比較或一般線性模型單因素分析進一步量化,事后多重比較方差齊性時用LSD法、非齊性時用T2法。為了尋找多種因素影響外來植物入侵的最佳子集,將可能對入侵產生影響的所有因子進行SPSS自動線性建模,包括林齡、林冠郁蔽度、優勢最大株高、植被總蓋度、土著物種豐富度、是否路邊、撫育時間、分區等8個因子。之后用R-lme4量化最佳子集對入侵的影響,以分區為隨機截距,最佳子集其他因子為解釋變量建立混合效應模型,逐步簡化模型至所有項都顯著[19]。調用effects包allEffects函數提取混合效應模型中各項的效應值,用plot(allEffects(model))命令繪制互作效應面板圖;當提取混合效應模型中某一項的效應時,其他各項處于其各水平樣本數加權平均值[20];在R3.5.2[21]上進行混合效應模型分析、互作效應提取和作圖。
2 結果與分析
老虎嶺林區共調查240個植被樣方,記錄到171種植物,隸屬于116科和162屬,外來植物24種,土著植物147種;其中禾本科植物最多,有21種,其次是菊科15種;外來植物豐富度樣方均值為1.35,蓋度均值26.0%;土著植物豐富度樣方均值4.54,蓋度均值39.6%。樣方中出現的外來植物包括9種一年生草本、9種多年生草本和6種木本植物。9個林內分組的林冠郁蔽度估算總均值53%,按降序排列為B區紅錐、D區西樺、A區馬尾松、E區麻櫟、E區國外松、C區鄧恩桉、B區尾巨桉、D區任豆、F區斑皮桉,分別為86%、71%、62%、60%、58%、55%、47%、26%和10%,T2多重比較結果為a、ab、b、abcd、abc、b、cd、de和e。15個分組的優勢最大株高平均值分別為:B區尾巨桉2.00 m、E區麻櫟1.79 m、C區鄧恩桉1.76 m、A區馬尾松1.63 m、D區任豆1.47 m、E區國外松1.22 m、B區紅錐1.19 m、路邊C 1.09 m、路邊A 1.06 m、路邊E 1.00 m、路邊D 0.91 m、路邊B 0.90 m、D區西樺0.81 m、F區斑皮桉0.76 m、路邊F 0.56 m。
2.1 不同分組的外來植物入侵程度比較
外來植物入侵對15個分組的一般線性模型單因素分析方差非齊性,從T2多重比較和均值95%置信區間可知,F區外來植物占比最高;除F區斑皮桉和E區麻櫟之外,路邊的外來植物入侵顯著高于其他所有林內分組(P<0.05);斑皮桉是引進樹種且林下外來植物入侵較多,但同為引進樹種的尾巨桉和國外松中外來植物入侵很少,鄧恩桉林下完全沒有外來植物(見圖1),因此并未發現引進樹種造林與鄉土樹種造林的林下外來植物入侵有差異。圖1中15個分組的外來植物占比估算均值從上到下分別為90.1%、62.5%、61.2%、57.5%、50.2%、49.1%、37.5%、19.7%、19.5%、5.2%、5.0%、4.5%、3.1%、2.2%和0。F區的路邊和林內入侵均較高,外來植物分布可能與分區關聯較與造林樹種關聯更密切,路邊多于林內。
2.2 林齡和撫育時間是否顯著影響外來植物分布格局
研究區林分包括6個林齡段:國外松和麻櫟40 yr,馬尾松22 yr,鄧恩桉9 yr,任豆、紅錐和西樺6 yr,尾巨桉和斑皮桉3 yr,路邊0 yr;6個林齡段的外來植物占比估算均值分別為11.3%、4.5%、0、9.0%、34.5%和59.8%;T2多重比較的結果為BC、C、C、C、B和A,路邊的外來植物入侵極顯著多于其他所有林齡段分組(P<0.01),其次是3 yr林分和40 yr林分。外來植物入侵隨林齡的變化是非線性的。
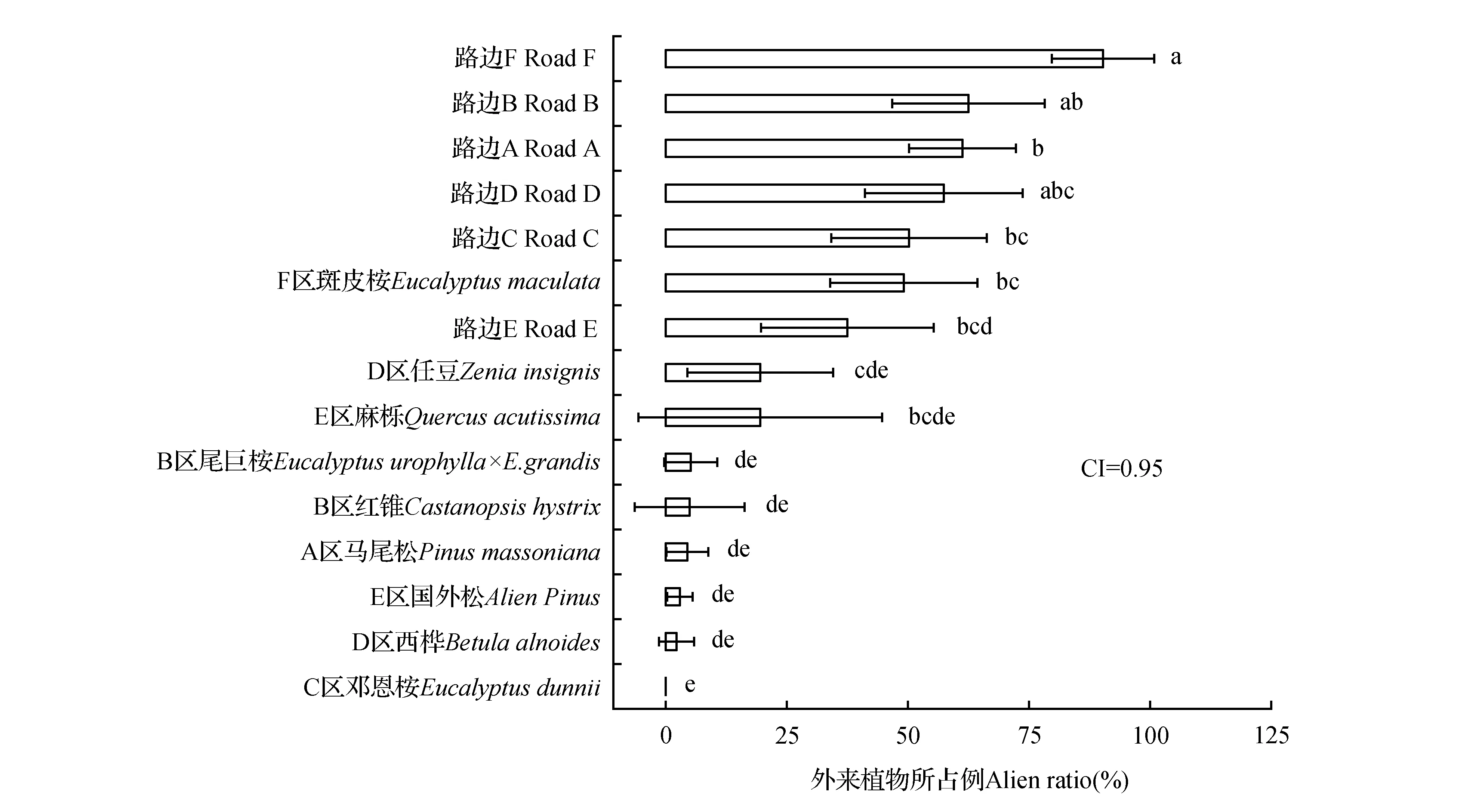
圖1 不同分組的入侵平均值、95%置信區間和SPSS一般線性模型T2多重比較結果 T2多重比較無相同字母表示在P<0.05水平上差異顯著。Fig.1 Means,95% confidence intervals and T2 multiple comparison results of invasions in 15 groups calculated by SPSS GLM-GEN Not sharing letter implied significant difference at P< 0.05 level.
之前4~6個月撫育(M1)、之前6~20個月撫育(M2)、之前20~36個月撫育(M3)、之前36個月以上無撫育(M4)等4種撫育時間的外來植物占比估算均值分別為33.0%、37.7%、4.5%和47.1%,4種撫育時間的入侵差異極顯著(F=35,df=5,P=0.000),多重比較表明M3與其他3種撫育時間的入侵差異極顯著,其他3種之間無顯著差異;只有A區馬尾松撫育時間為M3。雖然只比較撫育時間單個因子的方差分析表明不同撫育時間的入侵差異極顯著,從圖1中可見B區紅錐、B區尾巨桉、D區西樺和E區國外松的外來植物入侵與A區馬尾松的不相上下,但M1、M2和M4的其他組分拉高了它們的平均值;只比較單個因子均值、只分析主效應可能產生誤導的結論。
2.3 植被樣方位置對入侵的影響
植被樣方位置高于林道、齊平林道、低于林道對外來植物入侵影響極顯著(n=240,F=43,df=2,P=0.000),估算入侵均值分別為6.1、53.6和22.5,T2多重比較結果C、A和B,3種位置之間入侵差異均達極顯著。我們用SPSS“選擇個案”屏蔽所有路邊樣方,只對120個林內樣方進行樣方位置單因素的一般線性模型分析,樣方位置對入侵仍然影響極顯著(n=120,F=7,df=2,P=0.001),估算入侵均值分別為5.3、5.0和21.5,T2多重比較結果B、B和A;低于林道林內樣方的外來植物入侵極顯著高于其他兩種位置的林內樣方。
2.4 多因素的混合效應模型分析
以外來植物入侵為目標變量、其他有關因子為預測變量,利用SPSS自動線性建模從多種因素中篩選出解釋目標變量的最佳子集;中選模型準確性為65.2%,最佳子集包括是否路邊、林冠郁蔽度、土著物種豐富度、分區、林齡、優勢最大株高,分別占解釋率的37%、21%、19%、10%、7%和6%。以分區為隨機截距,最佳子集的其他因子為解釋變量建立混合效應模型,簡化后的最小適當模型見表1。在分區效應為平均截距、其他固定效應處于其均值的情況下,路邊對入侵的主效應為正且極顯著(P=0.000),是否路邊的外來植物入侵相差42.5%;林冠郁蔽度對入侵主效應為負(P=0.000),林冠郁蔽度每增加10%,外來植物入侵減少8.4%;樣方的優勢最大株高對入侵的主效應也為負,樣方的優勢最大株高每增加21 cm,外來植物占比減少5%(見表1)。
表1 以入侵為響應變量、分區為隨機截距、SPSS自動線性建模最佳子集為固定效應的混合效應模型分析結果
Table 1 Result of mixed effect model with alien ratio as response variable,minimum subset produced by SPSS automatic linear modeling as fixed effects,and district as random intercept
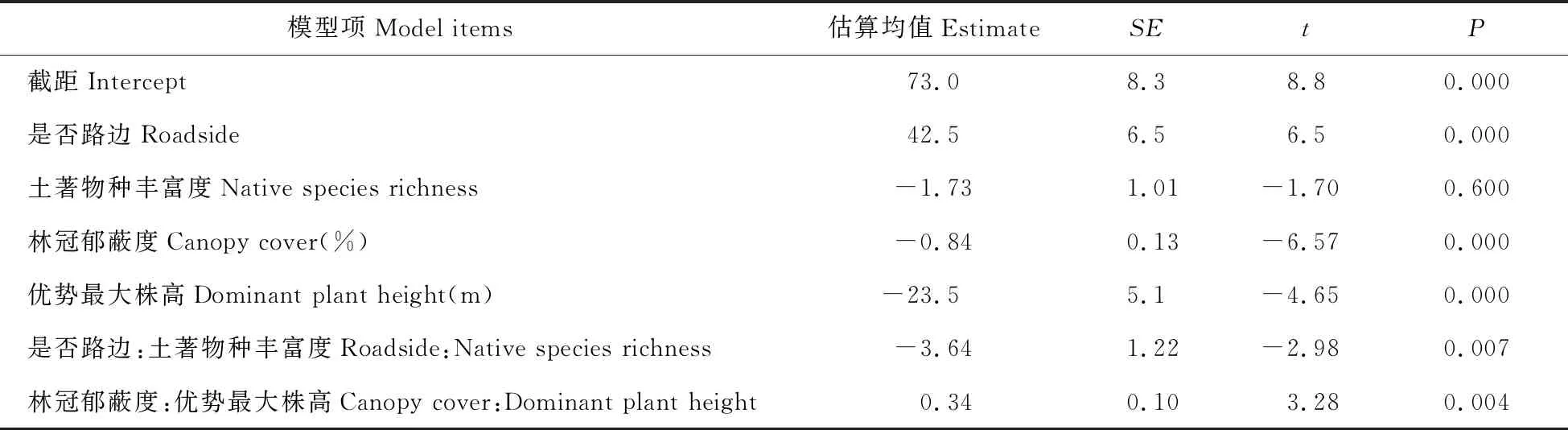
模型項Model items估算均值EstimateSEtP截距Intercept73.08.38.80.000是否路邊Roadside42.56.56.50.000土著物種豐富度Native species richness-1.731.01-1.700.600林冠郁蔽度Canopy cover(%)-0.840.13-6.570.000優勢最大株高Dominant plant height(m)-23.55.1-4.650.000是否路邊:土著物種豐富度Roadside:Native species richness-3.641.22-2.980.007林冠郁蔽度:優勢最大株高Canopy cover:Dominant plant height0.340.103.280.004

圖2 混合效應模型中的兩個互作效應 A.是否路邊*土著物種豐富度;B.林冠郁蔽度*優勢最大株高Fig.2 Two interactions in the mixed effect model A.Roadside*Snative(Native species richness); B.Canopy cover*Heightmax(Dominant plant height(m))
混合效應模型中包含兩個極顯著的互作項。在是否路邊和土著物種豐富度對入侵的互作效應中(見圖2a),當模型中其他項的效應為其均值時,不論土著物種豐富度多少,都呈現路邊入侵多于林內的趨勢。從互作效應提取數值中查看定住是否路邊時外來入侵隨土著物種豐富度的變化趨勢;先看在林內的情況,當土著物種豐富度為0、3、6、9和10時,入侵為28.3%、23.1%、17.9%、12.8%、11.0%;再看路邊的情況,土著物種豐富度為0、3、6、9和10時的入侵分別為70.8%、54.7%、38.6%、22.4%、17.1%;路邊土著物種豐富度明顯提升入侵抵抗性,但林內土著物種豐富度只能微弱增加入侵抵抗性;而當土著物種豐富度處于研究區均值(4.54)時,林內入侵約為20.5%,路邊入侵約為46.4%,相差約26%。
在林冠郁蔽度和優勢最大株高對入侵的互作效應中,當優勢最大株高較矮時(0.2~1.0 m),隨著林冠郁蔽度的減小入侵迅速增加;當樣方的優勢最大株高較高時(2 m),入侵隨林冠郁蔽度的變化不大(見圖2b)。當優勢最大株高≥4 m,林冠郁蔽度越高外來植物入侵越多,但這一趨勢的預測可靠性較低,因其置信區間較寬,可忽略不計。再從提取的互作效應數值中查看變化趨勢;林冠郁閉度為0時,優勢最大株高0.2、1.0和2.0 m對應的入侵分別為73.4%、54.6%和31.0%,負相關關系;林冠郁閉度為50%時,優勢最大株高為0.2、1、2、4和5 m的入侵分別為34.7%、29.6%、23.2%、10.3%和3.9%,兩者仍然負相關;當林冠郁閉度為70%,優勢最大株高同樣為0.2、1、2、4和5 m,外來植物入侵變為19.2%、19.6%、20.0%、20.9%和21.3%,幾乎持平不變。
3 討論
3.1 混合效應模型互作效應分析揭示共存因子的相互作用
利用混合效應模型明確地控制空間差異、具體地量化多因素的互作效應是本文的兩大特色。由于不同分區在干擾、拓殖種繁殖體庫、地形位置等方面有事先存在的差異,分區內的物種組成和植被結構存在自相關,入侵不僅與生物因子差異相關而且與區組作用相混淆,數量分析方法必需考慮影響變化的來源。本文的混合效應模型以分區作為隨機效應,允許每個分區有不同的截距,以此控制來源于分區或造林樹種的差別效應,多因素分析是在分區效應被均化后的結果。生物入侵研究目前仍難以提供可預測、可實用的知識,其中一個重要原因是大多數研究沒有明確地控制空間差異[11]。
互作效應是一個熟悉的概念,但互作效應中各因子引起的響應變化方向一直是陌生而模糊的;本文借助專業軟件的互作效應圖形化和數據提取來解釋互作效應。入侵研究通常分析單個環境因素的影響,忽略共存因素的互作效應,但實際上互作效應可能產生明顯的非加性效應,導致意想不到的后果[22];本文是從互作效應中發現路邊的土著物種豐富度明顯提升入侵抵抗性,林內的土著物種豐富度只能微弱增加入侵抵抗性,以及林冠郁蔽度和優勢最大株高在限制外來植物入侵上作用非可加。盡可能具體而清楚地了解的環境因子對外來入侵的互作效應,我們才能更有針對性地應對外來入侵威脅[23]。
3.2 模型解釋結果與前人發現的比較
建立在系統設計的第一手調查數據基礎之上的外來物種分布格局及其影響因子的研究目前還少見報道[24~25],較多前人研究是基于文獻資料和非系統設計調查。我們從道路條件、物種多樣性、林冠郁蔽度和優勢最大株高、造林樹種等幾個方面,對比前人的研究結果來闡述本文的新發現。
道路是入侵植物進入自然生態系統的途徑之一[26~27]。路邊溝渠微生境往往有較好光照、水分、接觸行走動物等條件,是入侵植物的最適宜定植點;土著植物通常對這些極端的條件比較敏感而不易定植,因此,路旁通常比周圍地區分布更多的外來植物[6]。老虎嶺林區的路邊外來植物入侵顯著高于林內;當其他因素效應處于其平均值時,路邊入侵較林內入侵高26%;而且,不論土著物種豐富度增加或減少,均呈現路邊入侵高于林內的趨勢。于永輝等在廣西的調查也發現林緣和林區道路的外來入侵植物種類最多[28];Wu等的研究結果顯示在地區尺度上離公路距離與植物入侵顯著相關[29]。除了認同路邊與外來入侵密切相關這一科學共識之外,本文具體分析了相對于林道的樣方位置高低對外來植物入侵的影響,發現在林道下方的林內樣方較易被入侵,林道上方不易被入侵,與林道齊平的林內樣方也不易被入侵;因此,在監測或防控林業外來植物時,重點應放在低于林道的森林。
物種多樣性是一個地區生態系統健康穩定的基礎[30~31]。通常認為土著物種豐富度高的地方對入侵的抵抗性強,但也有研究發現土著植物未對外來物種入侵發揮抵抗作用[23]或外來物種數量與土著種數量正相關[29]。混合效應模型分析中發現土著物種豐富度對入侵的主效應不顯著,但土著物種豐富度與路邊互作對入侵的效應極顯著;在林區道路旁,土著物種豐富度強烈地抑制外來植物入侵;但在森林內部外來植物入侵普遍較少時,土著物種豐富度對入侵的抑制作用不甚明顯。
混合效應模型中林冠郁蔽度和優勢最大株高都極顯著降低外來植物入侵。大多數小種子草本植物、樹木和灌木在林冠郁閉情況下無法定植[32];稠密的林冠和較高的林下植被可阻擋種子空降,減少外來植物繁殖體壓力;郁閉林分的建群種往往將資源利用得比較徹底,特別是光資源的利用;資源有限也使外來植物難以成功定植。Grime[32]將在較長時間尺度上才能產生的間接影響稱為過濾器效應或奠基者效應,本文的林冠郁蔽度和優勢最大株高是限制外來植物入侵的兩大過濾器(filter),但兩個因子的作用是非可加的,當林冠郁蔽度為零時優勢最大株高對入侵的抑制效應強大,隨著林冠郁蔽度的增加,優勢最大株高對入侵的抑制效應減弱,當林冠郁蔽度達70%時優勢最大株高對入侵的抑制效應幾乎為零;反之,當優勢最大株高較矮時,林冠郁蔽度對入侵的抑制效應明顯,而優勢最大株高≥2 m時林冠郁蔽度對入侵的抑制效應為零或轉變為促進作用。
造林樹種是群落優勢種、建群種,優勢種對群落結構和群落環境的形成有明顯的控制作用,對生態系統的直接效應在很大程度上由優勢種的性狀和功能多樣性決定[32]。本文的9個林內分組包含針葉林、干燥硬葉林、常綠闊葉林和落葉闊葉林;從單因素分析中可見,9種林內分組可分為高入侵(斑皮桉)、中等入侵(任豆、麻櫟)、低入侵(尾巨桉、紅錐、馬尾松、國外松和西樺)和無入侵(鄧恩桉)4個類型,兩個極端都是干燥硬葉林,沒有表現出哪類樹種較易被入侵,引進樹種造林與鄉土樹種造林對比在林下外來植物入侵方面也無明顯差異。
本文以樣方所有外來植物的相對百分比作為入侵程度指標,通過模型解釋發現:當其他因素效應處于其平均值時,路邊入侵較林內入侵高26%;樣方相對于林道的位置高低影響入侵程度,林道下方的樣方較易被入侵;土著物種豐富度對入侵的主效應不顯著,但與路邊的互作對入侵效應極顯著;林冠郁蔽度和優勢最大株高是限制外來植物入侵的兩大因素,對入侵的主效應和互作效應均顯著,但這兩個因子對入侵的作用非可加;林齡和撫育時間對外來植物入侵的影響趨勢不明顯。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中學物理·高中(2016年12期)2017-04-22 11:53:03
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
中國火炬(2014年11期)2014-07-25 10:31:58
