遮陰和鹽分對銀葉樹幼苗光合特性與葉綠素熒光參數的影響
2020-05-29 09:06:16張衛強黃芳芳甘先華黃鈺輝
生態環境學報 2020年3期
關鍵詞:影響
張衛強,黃芳芳,甘先華,黃鈺輝
廣東省森林培育與保護利用重點實驗室/廣東省林業科學研究院,廣東 廣州 510520
鹽分脅迫是影響植物生長的重要限制性因素(賈婷婷等,2018),對植物的傷害主要體現在離子脅迫、滲透脅迫及氧化脅迫等方面(潘晶等,2018),并抑制植株的光合作用來影響其生長發育(周丹丹等,2016)。光是植物光合作用和生長發育的重要影響因子,光強變化直接影響著植物的生長發育,植物對光強變化也會做出靈活的響應與適應(許大全,2013),光影響植物光合生理特性、體內營養物質吸收與分配、葉片形態與結構等生理過程,對植物生長發育產生影響(Cai,2011;Deng et al.,2012)。近年來,多數關于鹽脅迫的研究主要集中在植物耐鹽機制(王佺珍等,2017;潘晶等,2018)和鹽分脅迫對植物生長、葉綠素、光合特性、葉綠素熒光及生理生化的影響等方面(烏鳳章,2015;閆芳等,2016;陳盼飛等,2017;張雯莉等,2018;賈婷婷等,2018;潘晶等,2018),系統研究了植物對鹽脅迫的適應能力與響應機制。而植物對遮陰環境的適應性研究相對較少,主要集中在遮陰對植物生長、葉形態、光合特性、葉綠素熒光、種間競爭及氮素響應機制分配的影響等方面(劉悅秋等,2007;張云等,2014;宋洋等,2016;蔡建國等,2017;李小琴等,2019;杜瀾等,2019),分析了植物生長適宜的光照環境。
銀葉樹(Heritiera littoralis)隸屬于梧桐科銀葉樹屬,生長于潮間帶與陸地,不具有胎萌、氣生根及耐高滲透壓等典型紅樹植物特征(王伯蓀等,2002;曾聰等,2006),將其歸為半紅樹植物(王伯蓀等,2002)。深圳市鹽灶銀葉樹種群分布較集中,是中國最古老的銀葉樹種群,已表現出瀕危或易危的特征(簡曙光等,2004),尤其是濱海沼澤生境銀葉樹群落呈現為逐漸衰退的趨勢(陳曉霞等,2015),銀葉樹群落內,光照和鹽分是影響銀葉樹幼苗生長與更新的重要環境因子(郭樂東等,2019),改善濱海濕地銀葉樹群落內光照環境和降低土壤高鹽分是提升銀葉樹幼苗生長與更新能力的有效途徑。目前,有關銀葉樹的研究主要集中在生物學特性(簡曙光等,2005)、群落分布(簡曙光等,2004)、群落結構與多樣性(陳曉霞等,2015)、耐鹽機理(王秀麗,2014)、遺傳多樣性(Jian et al.,2002)及銀葉樹群落優勢樹種分布與土壤環境、群落競爭的關系(郭樂東等,2019)等方面。開展遮陰與鹽分對銀葉樹幼苗光合及葉綠素熒光研究鮮有報道。葉綠素熒光誘導作為聯系光合作用和環境關系的內在指標,被廣泛應用在植物抗逆性研究中(張國盛等,2017)。本研究以銀葉樹實生幼苗為研究對象,通過測定不同處理下(遮陰和施鹽)葉片葉綠素含量、光合光響應參數及葉綠素熒光動力學參數,研究了遮陰和施鹽處理下銀葉樹幼苗的光合生理及適應能力,以期為銀葉樹種群保育和更新提供科學依據。
1 材料與方法
1.1 研究區概況
試驗設在廣東省林業科學研究院(廣州)苗圃內,海拔25 m,典型亞熱帶季風氣候,年平均溫度23 ℃,最低月平均氣溫13.3 ℃(1月),最高月平均氣溫38.1 ℃(8月),年降水量1638 mm,4—9月的降水量占全年的80%,年平均濕度79%。
1.2 試驗設計
2018年9月選取96株長勢良好、生長一致的1年生實生苗移栽于上口徑16 cm、下口徑11.5 cm、高13.5 cm的容器,單株定植,栽培基質(黃心土與泥炭土,配置比例為5∶1)一致。實生苗平均樹高33.91 cm,平均地徑0.86 cm,容器苗置于大棚內,進行常規栽培管理,肥水管理一致。試驗前,完成了深圳鹽灶古銀葉樹群落沼澤生境的土壤鹽分含量與透光率的測定,土壤鹽分含量均值為2.5%,透光率均值約20%,以此為試驗設計依據,于2018年10月實施光照和鹽分交互處理試驗,光處理為全光照(自然光照)及20%透光率,鹽處理為不施鹽及2.5%鹽水處理,分別為全光照無鹽(CK)、全光照施鹽(FLS)、遮陰無鹽(SWS)及遮陰施鹽(SAS),共計4個處理,每個處理24株。選用黑色遮陽網進行遮光處理,施鹽頻度為每周3次,每次200 mL,60 d后停止。施鹽結束后,隨機抽取12盆進行土壤鹽分含量測定,土壤鹽分含量介于2.79%—3.01%之間,均值為2.90%。
1.3 光合光響應參數測定
光合光響應參數測定于2019年2月底至3月初進行,不同處理隨機挑選4盆長勢一致的植株,每株上選取2片長勢一致的葉片。采用LI-COR公司生產的LI-6800型便攜式光合測定儀測定葉片光合光響應參數。使用開放氣路,空氣流速為 0.5 L·min-1。光強選擇從 1800 μmol·m-2·s-1開始,依次降為:1800、1500、1200、1000、800、600、400、200、100、50、20、0 μmol·m-2·s-1。測定前葉片在1800 μmol·m-2·s-1光強下誘導 30 min,采用 CO2注入系統控制葉室CO2濃度,維持在400 mmol·mol-1左右。測定時每一光強點下最小光適應停留時間 2 min,最大光適應5 min。觀測時間均為上午9:00—11:00,每葉片重復3次,連續測定3 d。依據實驗數據繪制光合作用的光響應曲線(Pn-PAR),采用非直角雙曲線模型(Herrick et al.,1999)進行擬合,可求得最大凈光合速率(P′max)、表觀量子效率(a)、暗呼吸速率(Rd)等參數。
非直角雙曲線表達式為:
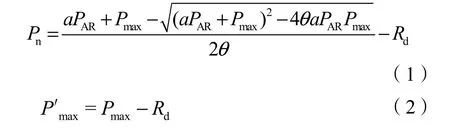
式中,Pn為凈光合速率,PAR為光合有效輻射,a為表觀量子效率,Pmax為最大總光合速率,P′max為最大凈光合速率,Rd為暗呼吸速率,θ為光合曲線彎曲程度曲度;光補償點(LCP)和光飽和點(LSP)采用Photosyn Assistant軟件擬合求得。
1.4 葉綠素熒光參數測定
葉綠素熒光參數測定與光響應參數同步,每組挑選4盆長勢一致的植株,每株上選取2片長勢一致的葉片。利用配置調制熒光葉室(Li-6400-40)的便攜式光合測定系統對葉片熒光動力學參數進行測定,測定前采用鋁箔紙包裹葉片暗適應,先測定初始熒光(Fo)、最大熒光(Fm)、最大PSII的光能轉換效率(Fv/Fm),然后打開活化光(1800 μmol·m-2·s-1),測定光適應下初始熒光(Fo′)、光適應下最大熒光(Fm′)、光適應下穩態熒光(FS)、光適應下最大可變熒光(Fv′),并計算光化學淬滅系數(qP)、非光化學猝滅系數(NPQ)、表觀量子傳遞速率(ETR)及實際光化學效率(ΦPSⅡ)等參數。測定時間為上午9:00—11:00和下午14:00—16:00,每葉片重復3次,連續測定3 d。吸收光能用于天線熱耗散 (D)=(1-Fv′/Fm′)×100%,光化學耗散 (P)=qP×Fv′/Fm′×100%,PSII反應中心非光化學能量耗散 (E)=(1-qP)×Fv′/Fm′×100%(Ehlert et al.,2008)。
1.5 葉綠素含量測定
葉綠素含量測定于2019年3月初進行,隨機選取不同處理銀葉樹各6株,摘取新鮮葉片,擦拭干凈,去主脈,剪碎混勻。稱取0.2 g,每處理3次重復,以95%丙酮研磨,提取,過濾,定容至25 mL,采用分光光度計測定提取液的吸收光譜,然后讀出各提取液分別在664、647、466 nm處的吸光度。計算每克葉片的葉綠素a(Chla)、葉綠素b含量(Chlb)(Pires et al.,2011)。
1.6 數據分析與統計方法
利用Microsoft Excel 2016整理原始數據,利用Two-way ANOVA分析光照和鹽分處理對光合特性和葉綠素熒光參數的影響,利用 Duncan法進行各參數指標間顯著性檢驗,所有分析在SPSS 16.0統計軟件進行。
2 結果與分析
2.1 遮陰與施鹽對銀葉樹葉綠素的影響
光和鹽處理對葉綠素含量、葉綠素a/b影響不顯著(P>0.05),二者不具有交互作用;光處理對葉片Chlb、Chl(a+b)及Chla/b存在顯著與極顯著的影響,而對Chla影響不顯著;鹽處理對Chla、Chlb、Chla/b及Chl(a+b)存在極顯著的影響;鹽分對葉綠素含量影響高于光照(表1)。在無鹽條件下,CK與 SWS處理間 Chla或 Chl(a+b)差異不顯著(P>0.05),而 Chlb或 Chla/b存在顯著差異(P<0.05);遮陰提高了 Chla、Chlb及 Chl(a+b)含量,增幅分別為10.26%、48.49%及18.67%,而遮陰顯著降低了Chla/b值(P<0.05),降幅達38.78%。在施鹽條件下,SAS處理下葉片Chlb和Chl(a+b)顯著高于FLS處理相應值(P<0.05),表明遮陰顯著提高了葉片 Chlb與 Chl(a+b)含量,增幅分別為127.27%與 84.09%,而遮陰顯著降低了 Chla/b值(P<0.05),降幅為 25.33%。無論全光照或遮陰,施鹽均顯著降低了 Chla、Chlb及 Chl(a+b)含量(P<0.05)。不同處理下 Chla、Chlb及 Chl(a+b)變化趨勢一致,均表現為SWS>CK>SAS>FLS。
2.2 遮陰與施鹽對銀葉樹光合光響應參數的影響
當 PAR 為 0 μmol·m-2·s-1時,Pn為負值,在弱光下,隨著PAR增強,Pn急劇升高,兩者呈線性關系,表明此時光是光合作用的主要限制因素;在無鹽條件下,當 PAR 達到 1000 μmol·m-2·s-1時,Pn上升趨勢減緩并趨向平穩;而在施鹽條件下,當PAR為 200 μmol·m-2·s-1時,Pn上升趨勢減緩,隨著 PAR的增加,Pn呈平緩下降趨勢(圖1)。光和鹽處理對葉片AQY、P′max及LSP影響不顯著(P>0.05),二者不具有交互作用;而光和鹽處理對葉片Rd和LCP存在極顯著的影響(P<0.01),二者存在極顯著的交互作用(表2);光處理對Rd、LCP及AQY存在極顯著與顯著影響,對P′max和LSP影響不顯著;鹽處理對AQY、P′max、Rd及LSP影響極顯著(P<0.01);鹽分對AQY、P′max、LSP影響顯著高于光照。在無鹽條件下,遮陰顯著提高了葉片AQY(P<0.05),在施鹽條件下,遮陰卻降低了AQY。無鹽下P′max與LSP顯著高于施鹽(P<0.05),說明施鹽對P′max與LSP影響顯著。全光無鹽下葉片Rd、LSP及LCP顯著高于其它處理(P<0.05),表明遮陰或施鹽降低了Rd、LSP及LCP。在遮陰或施鹽處理下,銀葉樹通過降低LCP、LSP來提高對弱光的利用能力,并通過降低呼吸消耗以維持植株正常生長,從而表現出了較強的適應能力。
2.3 遮陰與施鹽對銀葉樹熒光參數的影響
初始熒光(Fo)和最大熒光(Fm)反映PSII反應中心完全開放和完全關閉時的熒光產量(周丹丹等,2016);可變熒光(Fv)代表原初電子受體(QA)的還原狀態(張守仁,1999);Fv/Fo反映PSII最大量子效率(閆芳等,2016)。光和鹽處理對銀葉樹幼苗葉片Fo、Fm、Fv及Fv/Fo熒光參數影響不顯著(P>0.05),二者不存在交互作用;而光處理或鹽處理對Fm、Fv、Fv/Fo影響極顯著(P<0.01),鹽處理對Fo、Fm、Fv、Fv/Fo影響高于光處理(表3)。銀葉樹幼苗受到鹽分脅迫后葉片Fo有所增加,可能與PSII反應中心的 D1蛋白失活有關(Yang al.,2001),而Fm、Fv及Fv/Fo均有不同程度的降低,其中降幅最大為FLS處理,與對照存在顯著差異(P<0.05),說明FLS處理下銀葉樹幼苗葉片PSII反應中心已因鹽脅迫而受到了傷害,其潛在活性和原初光能轉換效率明顯減弱,產生了明顯的光抑制(閆芳等,2016)。
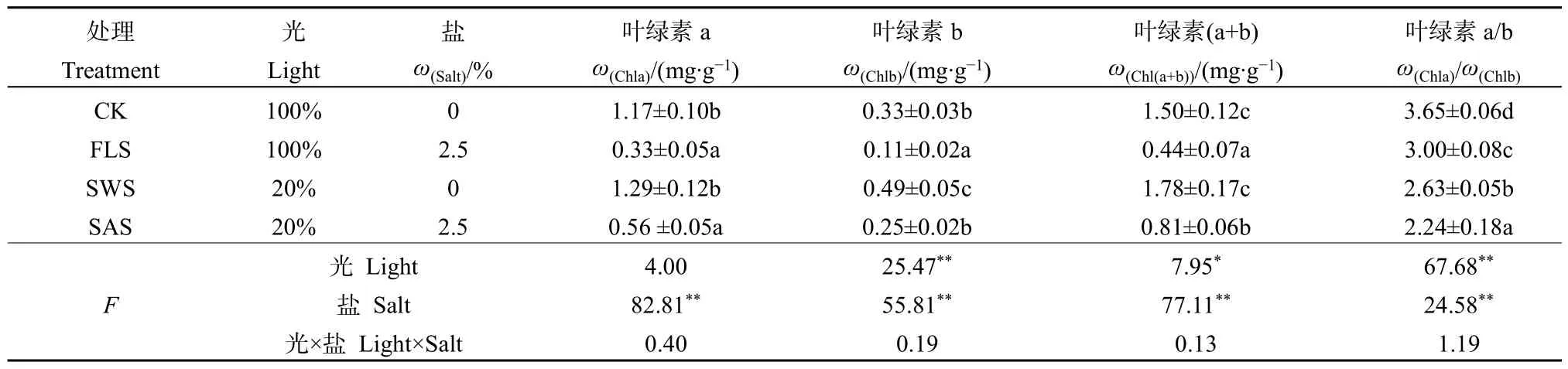
表1 遮陰與施鹽對銀葉樹葉綠素含量及比值的影響Table 1 Effects of shade and salt treatments on chlorophyll content and proportion in leaves of Heritiera littoralis
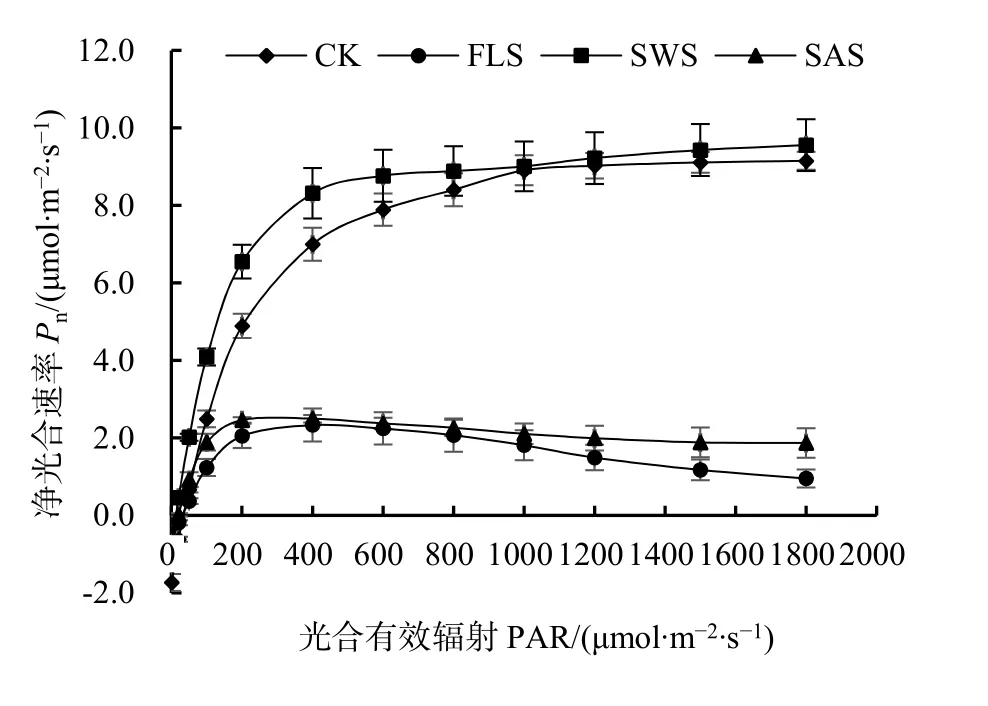
圖1 不同處理下銀葉樹葉片的光響應曲線Fig.1 Light response curves of net photosynthetic rate (Pn)in Heritiera littoralis under shade and salt treatments
Fv/Fm是暗反應下PSII最大光化學效率,Fv/Fm值變化可以反映光抑制的程度,比值越高其發生光抑制的程度越低(曹永慧等,2012);Fv′/Fm′是 PSII有效光化學量子產量,反映開放的PSII反應中心原初光能捕獲轉化效率(閆芳等,2016)。光和鹽處理對Fv/Fm、Fv′/Fm′影響不顯著(P>0.05),二者不存在交互作用;而鹽或光處理均對Fv/Fm與Fv′/Fm′產生顯著(P<0.05)和極顯著(P<0.01)的影響,鹽分的影響高于光照(表 4)。在無鹽或施鹽條件下,遮陰提升了PSII原初光能的轉化效率(Fv/Fm)和PSII有效光化學量子產量(Fv′/Fm′);而在全光或遮陰條件下,施鹽降低了Fv/Fm和Fv′/Fm′。不同處理下Fv/Fm和Fv′/Fm′分別介于 0.54—0.80 和0.25—0.49之間,兩者變化規律一致。Fv/Fm在CK、SWS及SAS處理間顯著差異(P>0.05),但顯著高于FLS(P<0.05),表明在FLS下銀葉樹幼苗發生光抑制程度明顯(圖 2);Fv′/Fm′在 CK、FLS 及 SAS間差異不顯著(P>0.05),但 CK 處理Fv′/Fm′明顯高于FLS處理,表明FLS處理下銀葉樹幼苗原初光能轉換效率降低。
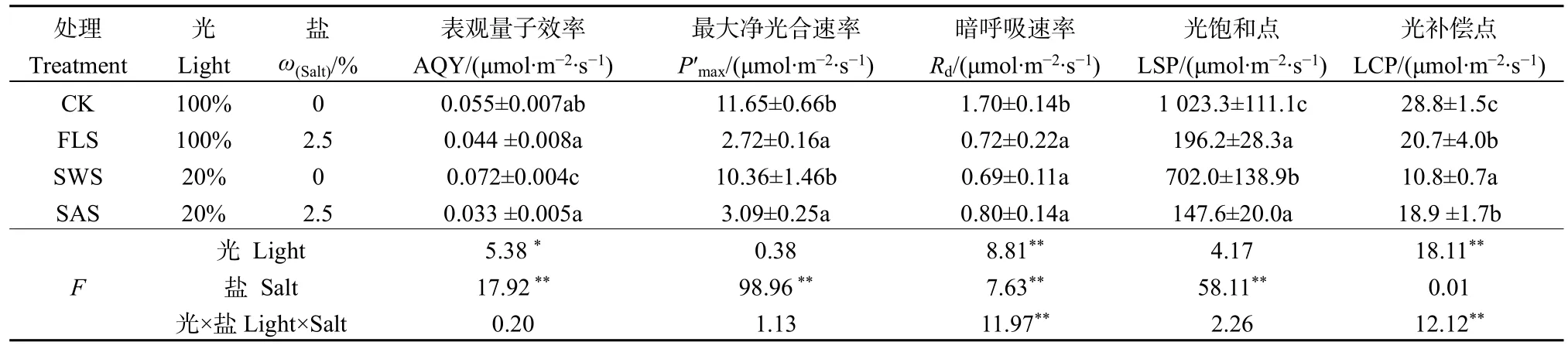
表2 遮陰與施鹽對銀葉樹光響應特性的影響Table 2 Effects of shade and salt treatments on photosynthesis-light response parameters of Heritiera littoralis
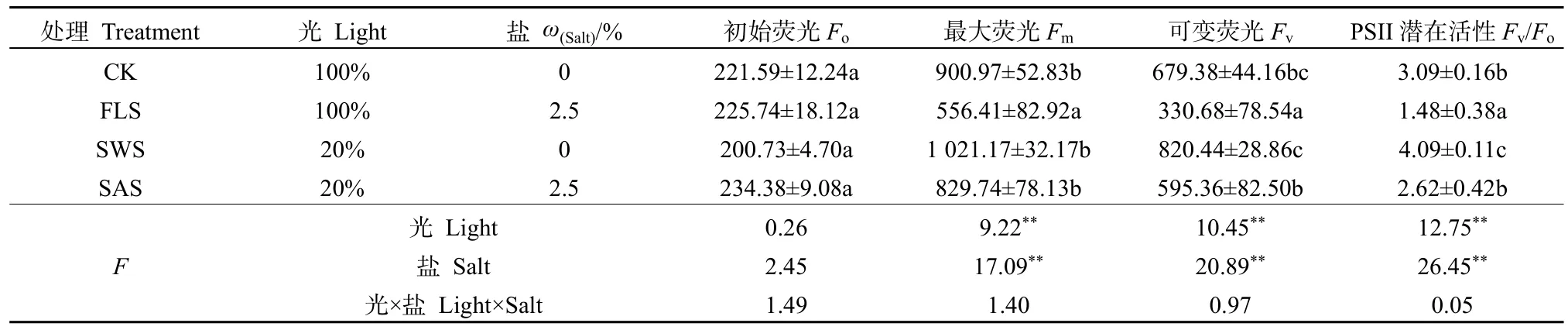
表3 遮陰與施鹽對銀葉樹葉片Fo、Fm、Fv及Fv/Fo的影響Table 3 Effects of shade and salt treatments on Fo,Fm,Fv,Fv/Fo' of Heritiera littoralis
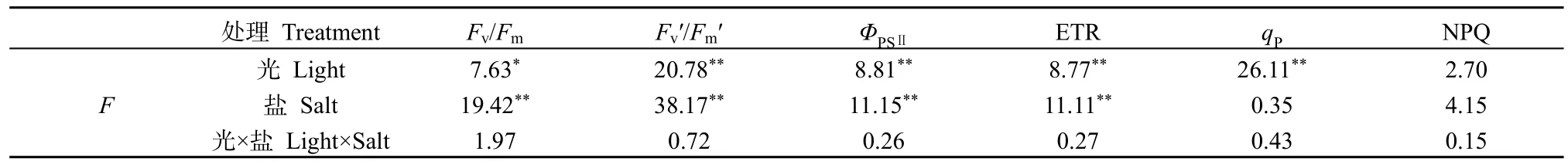
表4 遮陰與施鹽對銀葉樹葉片Fv/Fm等熒光參數的影響力分析Table 4 Effects of shade and salt treatments on fluorescence Parameters of Fv/Fm in Heritiera littoralis
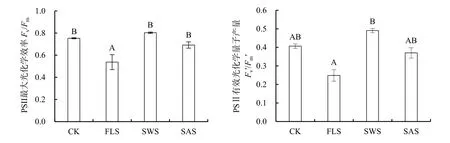
圖2 遮陰與施鹽對銀葉樹PSII最大光化學效率(Fv/Fm)和PSⅡ有效光化學量子產量(Fv′/Fm′)的影響Fig.2 Effects of shade and salt treatments on maximal quantum efficiency of PSII(Fv/Fm)and actual photochemical efficiency of PSII (Fv′/Fm′)of Heritiera littoralis
PSII實際光合效率(ΦPSⅡ)是反映電子傳遞活性與傳遞速率。表觀光合量子傳遞效率(ETR)是光合機構吸收光能發生電荷分離產生電子并沿電子傳遞鏈向下傳遞的速率,反映實際光強條件下的表觀電子傳遞效率(史彥江等,2012)。光和鹽處理對ΦPSⅡ與ETR影響不顯著(P>0.05),二者沒有交互作用(圖3);而鹽或光處理均對ΦPSⅡ與ETR產生極顯著影響(P<0.01),鹽分的影響高于光照(表4)。不同處理下ΦPSⅡ與ETR變化規律一致,均表現為CK> SWS>FLS>SAS,CK下ΦPSⅡ或ETR顯著高于其它處理(P<0.05),表明SWS、FLS及SAS處理顯著降低了PSⅡ反應中心電子傳遞活性及傳遞效率。
光化學猝滅系數(qP)反映了PSII反應中心的開放程度,qP越大,表明PSII的電子傳遞活性越大(Van Kooten et al.,1990);非光化學淬滅系數(NPQ)是 PSII天線色素吸收的光能不能用于光合電子傳遞而以熱的形式耗散掉的光能部分,反映了光系統對過剩光能的耗散能力(張守仁,1999;閆芳等,2016)。光和鹽處理對qP與 NPQ影響不顯著(P>0.05),二者沒有交互作用,鹽處理對qP與NPQ不顯著(P>0.05),而光處理對qP影響極顯著(P<0.01)(表4)。qP在CK與FLS處理間差異不顯著(P>0.05),顯著高于 SWS與 SAS處理(P<0.05),SWS與SAS處理下qP僅為CK的52.14%和38.80%,表明SWS與SAS處理下PSII中開放程度與電子傳遞活性降低(圖 4)。NPQ表現為CK>SWS>FLS>SAS,與ΦPSⅡ與ETR變化趨勢一致,CK處理下NPQ值顯著高于SAS(P<0.05),說明 SAS處理顯著降低了光合色素所吸收的光能用于保護性耗散比例(圖4)。
2.4 遮陰與施鹽對銀葉樹吸收光能的影響
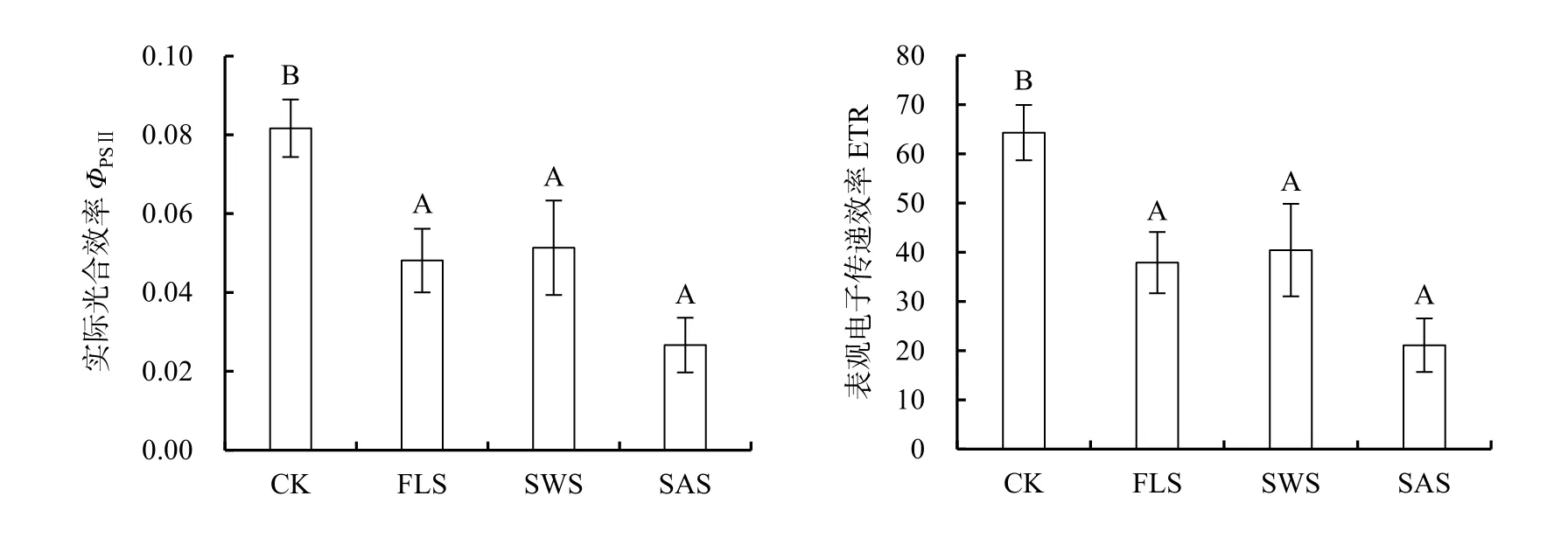
圖3 遮陰與施鹽對銀葉樹實際光合效率(ΦPSⅡ)和表觀電子傳遞速率(ETR)的影響Fig.3 Effects of shade and salt treatments on actual photosystem II efficiency (ΦPSⅡ) and electron transport rate (ETR) of Heritiera littoralis
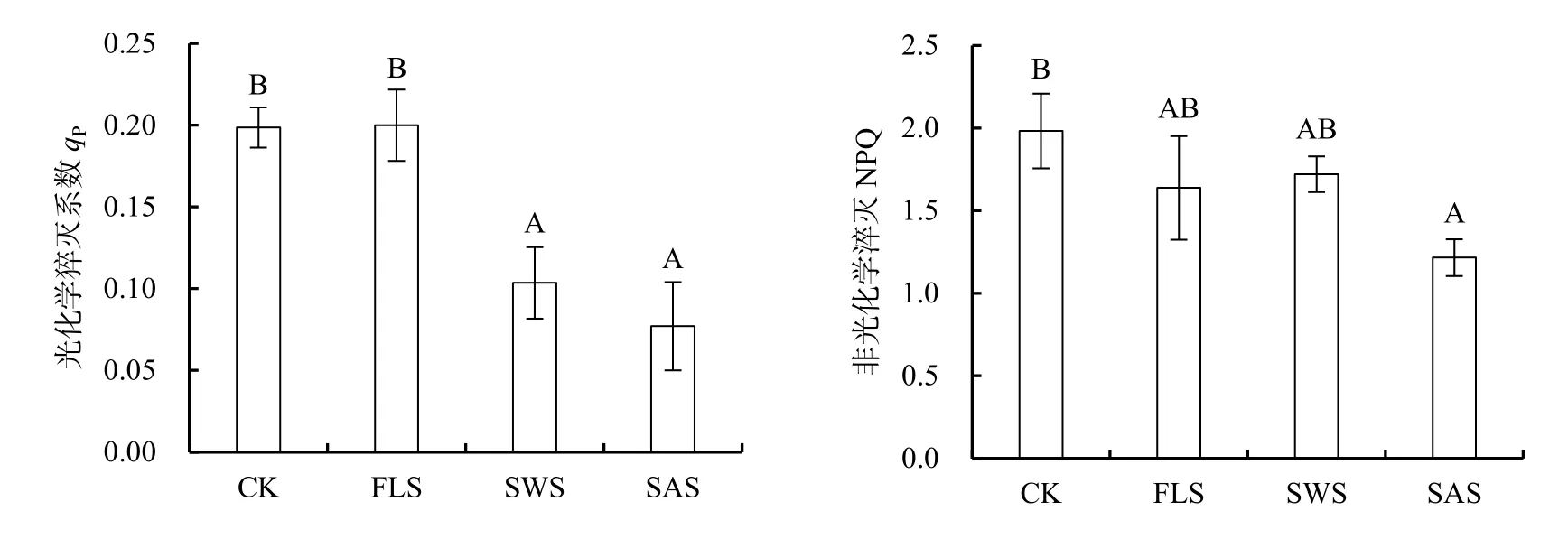
圖4 遮陰與施鹽對銀葉樹光化學淬滅系數(qP)和非光化學淬滅系數(NPQ)的影響Fig.4 Effects of shade and salt treatments on photochemical quenching (qP),non-photochemical quenching (NPQ) of Heritiera littoralis
植物葉片吸收的光能分為天線熱耗散的能量(D)、光化學反應的能量(P)及反應中心有非光化學反應耗散的能量(E)(Depuydt et al.,2009),分析吸收光能的分配將有助于了解植物葉片對吸收光能的分配策略(胡文海等,2015)。光和鹽處理對D、E、P影響不顯著(P>0.05),二者沒有交互作用;光或鹽處理對D、E、P影響達到極顯著(P<0.01),鹽分對D、P影響高于光照(表5)。P表現為:CK>SWS>FLS>SAS,表明CK下銀葉樹幼苗將吸收的光能較多的用于光化學反應,而SAS處理下用于光化學反應光能份額最低。FLS和SAS處理下天線熱耗散(D)與對照相比均有不同程度的升高,此時葉片具有較高的可調控生理代謝,能夠將過剩的光能較多地分配給熱耗散(蔡建國等,2017),而熱耗散比例的增加反映了FLS和SAS處理下銀葉樹幼苗光能捕獲效率降低,銀葉樹幼苗處于光抑制狀態。E表現為SWS>SAS>CK>FLS,在SWS和SAS處理下,較高的E值表明PSII光化學反應和保護性調節機制沒有發揮作用,積累了大量的激發能,這有可能會對PSII反應中心產生破壞作用(蔡建國等,2017)。正常環境下,銀葉樹幼苗葉片吸收的能量約60%用于熱耗散,約32%用于非光化學反應,僅有8%的能量用于光化學反應,說明在飽和光環境下銀葉樹葉片將過剩光能以熱耗散途徑釋放,削弱了反應中心過量激發能的積累(蔡建國等,2017)。
3 討論與結論
3.1 討論
3.1.1 遮陰與施鹽對葉綠素的影響
葉綠素作為光合色素中重要的色素分子,參與了光合作用中光能的吸收、傳遞和轉化(閆芳等,2016)。在無鹽條件下,遮陰增加了銀葉樹幼苗葉片Chla、Chlb及Chl(a+b)含量,促進了葉片中葉綠素b的合成,Chlb含量增幅顯著,增加了葉綠素b對藍紫光部分的吸收能力(劉悅秋等,2007),表明銀葉樹幼苗處于弱光環境下的生理適應及對弱光利用能力強。在全光條件下,施鹽顯著降低了Chl(a+b)、Chla、Chlb含量及Chla/b比值,這與閆芳等(2016)研究結果一致,這可能是由于鹽分脅迫下植物體內活性氧和葉綠素水解酶活性增加,破壞了葉綠體結構,葉綠素合成緩慢,分解增加,導致葉綠素含量降低(Agastian et al.,2000),進而影響光能吸收、轉化,導致光合速率下降(李海波等,2006)。有研究表明,陽性植物的Chla/b約為3,耐蔭植物的Chla/b約為2.3(孫小玲等,2010),銀葉樹幼苗正常環境下葉片Chla/b為3.65,屬陽性植物。
3.1.2 遮陰與施鹽對光合光響參數的影響
LSP和 LCP分別代表植物光合作用的光照強度上限和下限,同時也反映了植物葉片對強光和弱光的利用能力(蔡建國等,2017)。耐陰植物的LCP小于 20 μmol·m-2·s-1,LSP 為 50—1000 μmol·m-2·s-1或更低(Boardman,1977)。正常環境下銀葉樹LSP和 LCP 分別為 1023、28.8 μmol·m-2·s-1,說明銀葉樹為陽性植物。在無鹽條件下,遮陰顯著提高了銀葉樹幼苗AQY,卻顯著降低了LSP、LCP及Rd,這與王凱等(2009)、宋洋等(2016)及蔡建國等(2017)的研究結果類似,說明遮蔭處理下銀葉樹幼苗葉片通過降低LSP和LCP來提高對弱光的利用能力,同時降低呼吸消耗以維持植株正常生長,表現出較強的適應能力(蔡建國等,2017)。施鹽顯著降低了銀葉樹幼苗AQY、P′max及LSP,致使銀葉樹幼苗利用弱光能力和光合潛力降低,這可能是由于鹽脅迫激發滲透脅迫,導致氣孔關閉,引起光合作用的氣孔限制(賈婷婷等,2018;許大全,2013)。
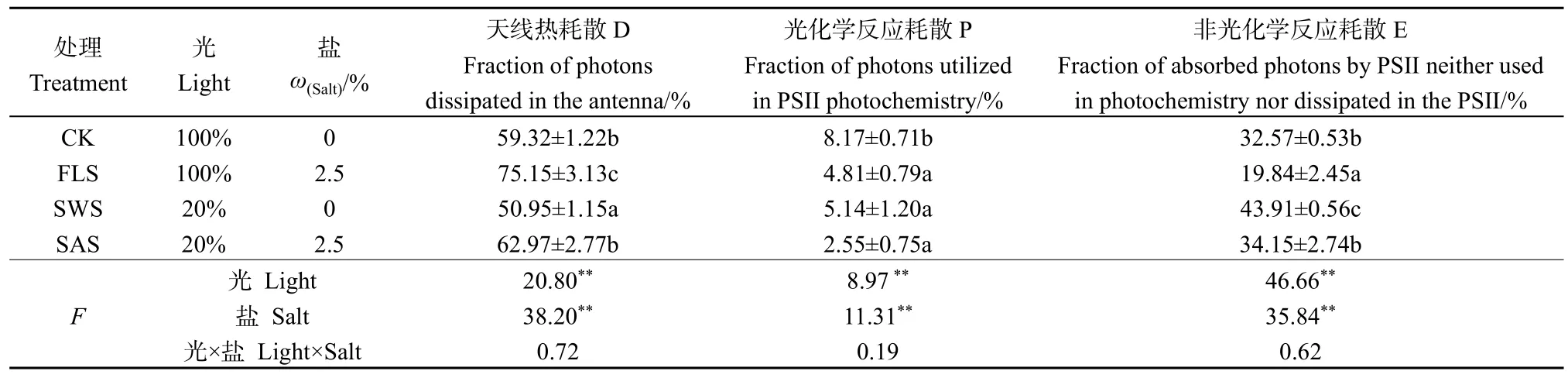
表5 不同處理下銀葉樹葉片吸收光能的分配Table 5 The distribution of light energy absorbed in Heritiera littoralis under shade and salt treatments treatments
3.1.3 遮陰與施鹽對葉綠素熒光參數的影響
Fv/Fm反映PSII反應中心最大PSII光能轉換效率(王文林等,2013)。正常情況下,沒有環境脅迫并經過充分暗適應的植物葉片Fv/Fm是比較恒定的,一般為 0.80—0.85(許大全,2013)。FLS處理下Fv/Fm與對照相比顯著下降了 28.57%,表明此時鹽分脅迫使PSII受到了傷害,最大PSII光能轉換效率減弱,植物體內發生了光合作用的光抑制。不同處理下銀葉樹幼苗ΦPSⅡ與 ETR變化趨勢一致,均表現為全光無鹽 (CK)>遮陰無鹽 (SWS)>全光施鹽(FLS)>遮陰施鹽 (SAS),表明遮陰或施鹽使ΦPSⅡ與ETR降低,而施鹽對ΦPSⅡ與ETR的影響高于遮陰,這主要是由于鹽脅迫使銀葉樹幼苗發生了明顯的光抑制,致使PSII潛在活性中心受到損傷,PSII的光化學活性受抑制,光能轉換效率與電子傳遞能力下降(閆芳等,2016;烏鳳章,2015),而遮陰一定程度增加了PSII潛在活性,降低了PSII反應中心因失活而出現光抑制(蔡建國等,2017)。銀葉樹幼苗qP表現為全光施鹽 (FLS)>全光無鹽 (CK)>遮陰無鹽 (SWS)>遮陰施鹽 (SAS);在無鹽或施鹽條件下,遮陰降低了銀葉樹幼苗qP,PSII反應中心的開放程度與電子傳遞活性降低,銀葉樹天線色素吸收的光能有相當一部分用于熱耗散,降低光能對光合系統產生破壞。高的 NPQ有利于過剩光能的及時耗散,避免光系統的損害,并能夠將更多的光能用于光合電子傳遞(馮漢青等,2013;周丹丹等,2016);不同處理下NPQ變化趨勢與ΦPSⅡ與ETR趨勢一致,表明遮陰或施鹽降低了PSII天線色素吸收光能以熱的形式耗散的比例,尤其是在SAS處理下,PSII過剩光能以熱耗散的比例減少,降低了光能轉化效率與光合電子傳遞能力,增加了過剩光能對光系統的破壞的風險。
3.2 結論
植物葉綠素含量、光合光響應參數及葉綠素熒光參數均可作為快速準確的判斷銀葉樹幼苗耐陰與耐鹽程度的重要指標。光和鹽對葉綠素含量、光合光響應參數(AQY、P′max及LSP)及葉綠素熒光參數的影響沒有交互作用,鹽對葉綠素含量、光合光響應參數(AQY、P′max及LSP)及葉綠素熒光參數(Fo、Fm、Fv、Fv/Fo、ΦPSⅡ、ETR 及 NPQ)的影響高于光照。在正常環境下,銀葉樹為陽性樹種,其幼苗表現出較高的耐陰性及對弱光的利用能力。鹽脅迫使銀葉樹幼苗發生了明顯的光抑制,PSII的光化學活性受抑制,過剩光能對光系統的破壞的風險增加,光能轉換效率與電子傳遞能力下降,光合作用效率大幅降低。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
