miR-26b-3p靶向調(diào)控TRA2B表達(dá)抑制神經(jīng)膠質(zhì)瘤細(xì)胞的增殖和轉(zhuǎn)移
2020-05-23 01:19:54隋銳張燁姚冰孫佩欣樸浩哲
分子診斷與治療雜志 2020年3期
關(guān)鍵詞:檢測(cè)
隋銳 張燁 姚冰 孫佩欣 樸浩哲
神經(jīng)膠質(zhì)瘤是常見(jiàn)的一類惡性腫瘤,其發(fā)病率在我國(guó)逐年增加,并且往往伴隨著較差的預(yù)后,嚴(yán)重威脅人們的身體健康[1-2]。術(shù)后腫瘤的復(fù)發(fā)及轉(zhuǎn)移是限制神經(jīng)膠質(zhì)瘤腦移植發(fā)展的關(guān)鍵問(wèn)題[3]。目前治療的主要方法包括手術(shù)切除和介入化療栓塞等手段,但以上方式的療效仍有待進(jìn)一步確證,其臨床效果均不理想[4]。因此提早診斷神經(jīng)膠質(zhì)瘤的發(fā)生和采取相應(yīng)措施將會(huì)極大地改善神經(jīng)膠質(zhì)瘤患者的預(yù)后情況。微小RNA(miRNAs)是長(zhǎng)度為20~22nt 的非編碼RNA,miRNAs與目的mRNA 結(jié)合從而調(diào)控mRNA的表達(dá)[5]。miRNAs 具有極其重要的作用,它參與細(xì)胞增殖分化、應(yīng)激及凋亡等一系列生物學(xué)過(guò)程,因而在腫瘤研究中具有很大意義[6]。miR-26b-3p 在多種惡性腫瘤組織中檢測(cè)到表達(dá)上調(diào),比如宮頸癌、胰腺癌以及肺癌等,在癌癥的發(fā)生發(fā)展以及預(yù)后過(guò)程中發(fā)揮重要作用[7-9]。但是國(guó)內(nèi)關(guān)于miR-26b-3p 在神經(jīng)膠質(zhì)瘤增殖與轉(zhuǎn)移中的作用研究相對(duì)缺乏。我們將利用熒光定量PCR 技術(shù)檢測(cè)膠質(zhì)瘤患者組織中miR-26b-3p 的表達(dá)情況,并運(yùn)用細(xì)胞學(xué)實(shí)驗(yàn)探究miR-26b-3p 在神經(jīng)膠質(zhì)瘤細(xì)胞侵襲中起的作用。
1 材料和方法
1.1 標(biāo)本來(lái)源
收集2016年8月至2017年8月來(lái)本院腫瘤科就診的32例神經(jīng)膠質(zhì)瘤組織切片和癌旁組織切片(癌組織3~4 cm 處)。入組標(biāo)準(zhǔn):①不存在其他惡性腫瘤或其他系統(tǒng)性疾病;②屬于原發(fā)性神經(jīng)膠質(zhì)瘤。排除標(biāo)準(zhǔn):①手術(shù)前進(jìn)行過(guò)放療、化療以及免疫治療的患者;②服用抗腫瘤藥物或進(jìn)行放療等治療的患者。
1.2 細(xì)胞培養(yǎng)與轉(zhuǎn)染
①取出凍存神經(jīng)膠質(zhì)瘤細(xì)胞SHG-44,水浴鍋加溫融化(水溫37℃),低溫低速離心4 分鐘,棄上清,加入1 mL RPMI-1640 培養(yǎng)液,再放入培養(yǎng)箱進(jìn)行培養(yǎng)(5% CO2,溫度為37℃)。②在鼎盛生物公司合成miR-26b-3p inhibitor和mimic,取出兩組細(xì)胞作為miR-26b-3p inhibitor組和miR-26b-3p mimic 進(jìn)行細(xì)胞轉(zhuǎn)染,按Lipofectamine?2000 Transfection Reagent 說(shuō)明書(shū)實(shí)行細(xì)胞轉(zhuǎn)染操作;陰性對(duì)照組不做處理,進(jìn)行后續(xù)實(shí)驗(yàn)。
1.3 癌細(xì)胞Total RNA的抽提
轉(zhuǎn)染24 h 后,用胰酶對(duì)各組SHG-44 細(xì)胞消化,收集細(xì)胞懸液;離心管進(jìn)行低速離心3 min,再加入1.5 mL Trizol 裂解液,重懸混勻。取離心之后的水相放在1.5 mL 離心管,加入異丙醇震蕩混勻。離心管低速離心12 min 后采用600 μL,75%的酒精洗滌四次RNA 沉淀。加入100 μL RNasefree 溶解RNA,使用NanoDrop 測(cè)定RNA 濃度,再放在-20℃冰箱備用。
1.4 miRNA的反轉(zhuǎn)錄與定量PCR檢測(cè)
按照試劑盒說(shuō)明書(shū)進(jìn)行體外反轉(zhuǎn)錄實(shí)驗(yàn)得到cDNA 產(chǎn)物,從而獲得cDNA 模版。反應(yīng)體系為TaqMan?Universal PCR Master Mix 20 μL,用ΔCT 表示miR-26b-3p 相對(duì)表達(dá)量(ΔCT=|CTmiR-26b-3p-CTU6|),以U6 作為內(nèi)參。以同樣的方法檢測(cè)TRA2B在RNA水平的表達(dá),PCR 擴(kuò)增引物如表1。

表1 各引物序列信息Table1 Sequence information of each primer
1.5 CCK8 檢測(cè)細(xì)胞增殖活力
轉(zhuǎn)染24 h 后,將SHG-44 細(xì)胞接種在96 孔板(6×103細(xì)胞/孔),每孔分別加入12 μL CCK-8 溶液,放置在培養(yǎng)箱中。當(dāng)?shù)竭_(dá)時(shí)間節(jié)點(diǎn)24、48和72 h,分別檢測(cè)各組細(xì)胞吸光度值(OD450nm)。
1.6 細(xì)胞劃痕實(shí)驗(yàn)
取各組SHG-44 細(xì)胞接種6 孔板(5×106個(gè)/孔),使用培養(yǎng)箱過(guò)夜培養(yǎng)。用無(wú)菌槍頭進(jìn)行劃痕,采用1xPBS 對(duì)各組細(xì)胞洗滌5次,加入培養(yǎng)基后拍照為0 h 時(shí)細(xì)胞的遷移情況。繼續(xù)進(jìn)行培養(yǎng),24 h 后顯微鏡下拍照,兩次拍照之間的劃痕距離可衡量各組細(xì)胞的遷移能力。
1.7 統(tǒng)計(jì)分析
采用SPSS 22.0 軟件進(jìn)行分析,計(jì)數(shù)資料用n(%)表示,行χ2檢驗(yàn);計(jì)量資料用()表示,兩組間比較用t檢驗(yàn),多組間比較使用單因素方差分析;P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 組織切片HE 染色結(jié)果
對(duì)神經(jīng)膠質(zhì)瘤組織切片和癌旁組織切片進(jìn)行HE 染色發(fā)現(xiàn),神經(jīng)膠質(zhì)瘤組織組織出現(xiàn)少量點(diǎn)狀壞死,細(xì)胞間有部分炎性細(xì)胞的浸潤(rùn);而癌旁組織結(jié)構(gòu)完整,無(wú)明顯細(xì)胞壞死(圖1)。

圖1 腦組織HE 染色結(jié)果比較(HE,×200)Figure1 Comparison of HE staining results of brain tissue(HE,×200)
2.2 miR-26b-3p與TRA2B 在神經(jīng)膠質(zhì)瘤組織中的表達(dá)
通過(guò)q-PCR檢測(cè)癌組織和癌旁組織切片中miR-26b-3p 相對(duì)表達(dá)量發(fā)現(xiàn),神經(jīng)膠質(zhì)瘤組織中其相對(duì)表達(dá)量顯著高于癌旁組織,差異有統(tǒng)計(jì)學(xué)意義(P<0.05);神經(jīng)膠質(zhì)瘤組織中TRA2B相對(duì)表達(dá)量顯著低于癌旁組織,差異有統(tǒng)計(jì)學(xué)意義(P<0.05),見(jiàn)表2。進(jìn)一步對(duì)患者癌組織中miR-26b-3p與TRA2B相對(duì)表達(dá)量進(jìn)行分析發(fā)現(xiàn),miR-26b-3p與TRA2B的表達(dá)呈顯著負(fù)相關(guān)(r=-0.550,P<0.05),見(jiàn)圖2。
2.3 各組SHG-44 細(xì)胞中miR-26b-3p與TRA2B 的相對(duì)表達(dá)量比較
利用q-PCR檢測(cè)各組細(xì)胞miR-26b-3p 的相對(duì)表達(dá)量,miR-26b-3p inhibitor組中的miR-26b-3p 的表達(dá)顯著降低,miR-26b-3p mimic組中miR-26b-3p的表達(dá)顯著升高,與陰性對(duì)照組比較,差異有統(tǒng)計(jì)學(xué)意義(P<0.05),如表3。
表2 不同腦組織中miR-26b-3p和TRA2B 的相對(duì)表達(dá)量(±s)Table2 The relative expression of miR-26b-3p and TRA2B in different brain tissues(±s)

表2 不同腦組織中miR-26b-3p和TRA2B 的相對(duì)表達(dá)量(±s)Table2 The relative expression of miR-26b-3p and TRA2B in different brain tissues(±s)
神經(jīng)膠質(zhì)瘤組織癌旁組織t 值P 值n 32 32 miR-26b-3p 相對(duì)表達(dá)量2.35±0.15 1.57±0.21 6.12<0.001 TRA2B 相對(duì)表達(dá)量0.69±0.10 1.55±0.12 11.56<0.001
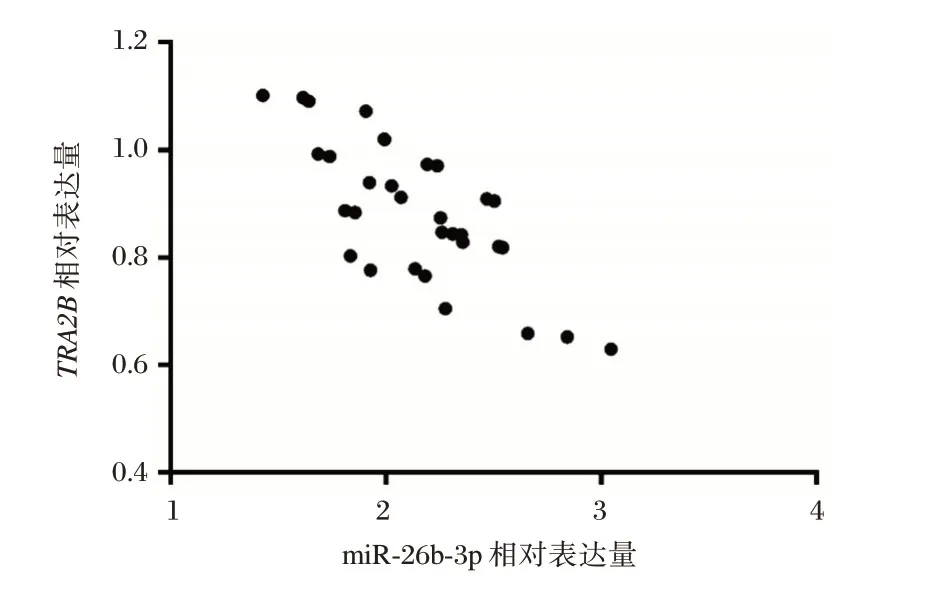
圖2 神經(jīng)膠質(zhì)瘤組織中miR-26b-3p與TRA2B表達(dá)的相關(guān)性分析Figure2 Correlation Analysis of the expression of miR-26b-3p and TRA2B in glioma tissues

表3 各組SHG-44 細(xì)胞中miR-26b-3p和TRA2B的相對(duì)表達(dá)量Table3 The relative expression of miR-26b-3p and TRA2B in SHG-44 cells of each group
2.4 miR-26b-3p 對(duì)SHG-44 細(xì)胞增殖活性的影響
檢測(cè)轉(zhuǎn)染24、48與72 h 后各組SHG-44 細(xì)胞吸光值,miR-26b-3p mimic組的增殖率在轉(zhuǎn)染24、48和72 h 后增值率顯著高于陰性對(duì)照組(P<0.05);miR-26b-3p inhibitor組的增殖率在各個(gè)時(shí)間點(diǎn)增值率均低于陰性對(duì)照組,均具有統(tǒng)計(jì)學(xué)意義(P<0.05),如圖3。

圖3 miR-26b-3p 對(duì)SHG-44 細(xì)胞增殖活性的影響Figure3 Effect of miR-26b-3p on the proliferation of SHG-44 cells
2.5 miR-26b-3p 對(duì)SHG-44 細(xì)胞遷移能力的影響
轉(zhuǎn)染24 h 后,利用細(xì)胞劃痕實(shí)驗(yàn)檢測(cè)各組SHG-44 細(xì)胞的遷移能力,miR-26b-3p inhibitor組SHG-44 細(xì)胞遷移距離小于陰性對(duì)照組(P<0.05);miR-26b-3p mimic組SHG-44 細(xì)胞遷移距離顯著高于陰性對(duì)照組(P<0.05),見(jiàn)圖4。

圖4 不同組細(xì)胞遷移能力的比較(臺(tái)盼藍(lán)染色,×200)Figure4 Comparison of cell migration in different groups(臺(tái)盼藍(lán)染色,×200)
3 討論
目前普遍認(rèn)為細(xì)胞的惡性增殖是癌癥發(fā)生的必要環(huán)節(jié),近年來(lái)對(duì)神經(jīng)膠質(zhì)瘤的研究發(fā)現(xiàn),miRNA 通過(guò)其對(duì)關(guān)鍵基因表達(dá)的調(diào)控,比如P53,在癌細(xì)胞的增殖、遷移、侵襲以及凋亡過(guò)程中起著重要作用[10-11]。在神經(jīng)膠質(zhì)瘤細(xì)胞中表達(dá)上調(diào)的miRNA 包括miRNA-183,miRNA-21,miRNA-221 等;在神經(jīng)膠質(zhì)瘤細(xì)胞中表達(dá)下調(diào)的miRNA 有miRNA-122,miRNA-145 以及miRNA-26a 等,其表達(dá)水平不盡相同,作用機(jī)制各不相同[11-12]。
有研究發(fā)現(xiàn),miR-122 在正常腦組織中表達(dá)水平較高,占全部miRNAs 表達(dá)量的70%,而其表達(dá)的下調(diào)與癌癥的發(fā)生發(fā)展密切相關(guān),其在神經(jīng)膠質(zhì)瘤細(xì)胞中的下調(diào)會(huì)導(dǎo)致其靶基因如腦細(xì)胞核因子3A(Hepatocyte nuclear factor 3A,HNF3A)的表達(dá)升高,而HNF3A均是腦細(xì)胞細(xì)胞周期重要的調(diào)控蛋白,從而導(dǎo)致細(xì)胞周期紊亂,引發(fā)癌癥的發(fā)生[13-14]。Huang 等對(duì)SHG-44 細(xì)胞系的研究發(fā)現(xiàn),當(dāng)提高細(xì)胞內(nèi)源miR-122 的表達(dá)水平時(shí),細(xì)胞中caspase9/3 蛋白的表達(dá)水平明顯提高,同是細(xì)胞凋亡率顯著提高;進(jìn)一步對(duì)miR-122 靶基因進(jìn)行分析發(fā)現(xiàn),其可能是通過(guò)抑制內(nèi)源Bcl-w 蛋白的表達(dá),而實(shí)現(xiàn)對(duì)細(xì)胞凋亡的調(diào)控,表明在神經(jīng)膠質(zhì)瘤細(xì)胞中miR-122 的表達(dá)上調(diào),可抑制內(nèi)源Bcl-w 蛋白表達(dá)從而調(diào)控神經(jīng)膠質(zhì)瘤細(xì)胞的凋亡過(guò)程,在神經(jīng)膠質(zhì)瘤的發(fā)生發(fā)展中起著重要作用[15]。在癌細(xì)胞中對(duì)miR-26b-3p 的表達(dá)進(jìn)行研究發(fā)現(xiàn),miR-26b-3p在癌細(xì)胞中的表達(dá)水平顯著高于癌旁組織,而下調(diào)miR-26b-3p 在癌細(xì)胞系中的表達(dá)會(huì)明顯抑制血細(xì)胞的生長(zhǎng)增殖;進(jìn)一步通過(guò)流式細(xì)胞儀對(duì)細(xì)胞的凋亡情況進(jìn)行檢測(cè)發(fā)現(xiàn),下調(diào)癌細(xì)胞系Panc-1 內(nèi)源miR-26b-3p 表達(dá),癌細(xì)胞的凋亡率顯著提高,集聚能力明顯下降,表明miR-26b-3p 在癌癥的發(fā)生發(fā)展中起著癌基因的作用[16]。
本研究在神經(jīng)膠質(zhì)瘤細(xì)胞中對(duì)miR-26b-3p 及其靶基因的進(jìn)一步探究,發(fā)現(xiàn)miR-26b-3p 在神經(jīng)膠質(zhì)瘤患者癌組織中表達(dá)明顯上調(diào);在神經(jīng)膠質(zhì)瘤組織中TRA2B的表達(dá)水平顯著下降。Pearson相關(guān)性分析顯示,在32例神經(jīng)膠質(zhì)瘤患者神經(jīng)膠質(zhì)瘤組織中,TRA2B的表達(dá)水平與miR-26b-3p 表達(dá)呈顯著負(fù)相關(guān),進(jìn)一步證實(shí)了miR-26b-3p 對(duì)TRA2B的靶向調(diào)控作用。近來(lái)研究發(fā)現(xiàn),TRA2B表達(dá)水平降低可導(dǎo)致多種腫瘤細(xì)胞的遷移和侵襲能力減弱[17]。這表明miR-26b-3p 通過(guò)靶向調(diào)控TRA2B的表達(dá),在神經(jīng)膠質(zhì)瘤細(xì)胞增殖和遷移過(guò)程中發(fā)揮重要作用。
綜上所述,本研究首次在神經(jīng)膠質(zhì)瘤中研究miR-26b-3p 的表達(dá),發(fā)現(xiàn)在神經(jīng)膠質(zhì)瘤癌細(xì)胞中,miR-26b-3p 的表達(dá)明顯上調(diào),而TRA2B的表達(dá)被抑制;在神經(jīng)膠質(zhì)瘤細(xì)胞系中的研究進(jìn)一步證實(shí),miR-26b-3p 通過(guò)靶向調(diào)控癌細(xì)胞中TRA2B的表達(dá),促進(jìn)神經(jīng)膠質(zhì)瘤細(xì)胞的增殖和遷移,與腫瘤的發(fā)生發(fā)展密切相關(guān)。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
