Seipin敲低大鼠PC12細(xì)胞株的構(gòu)建和對細(xì)胞凋亡的影響*
2020-05-04 07:58:50呂菊胡玉梅任真奎謝鵬張春林焦玲禹文峰
貴州醫(yī)科大學(xué)學(xué)報(bào) 2020年3期
關(guān)鍵詞:效果
呂菊,胡玉梅,任真奎,3,謝鵬,張春林,焦玲,禹文峰**
(1.貴州醫(yī)科大學(xué) 分子生物學(xué)重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽 550004;2.貴州醫(yī)科大學(xué) 地方病與少數(shù)民族疾病教育部重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽 550004;3.黔西南州人民醫(yī)院 檢驗(yàn)科,貴州 興義 562400;4.貴州醫(yī)科大學(xué) 基礎(chǔ)醫(yī)學(xué)院,貴州 貴陽 550025;5.貴州醫(yī)科大學(xué)附院 神經(jīng)內(nèi)科,貴州 貴陽 550004)
帕金森病(parkinson disease,PD)的病理特征是中樞和外周特定神經(jīng)元群的神經(jīng)退行性病變和殘存的多巴胺(dopamine,DA)能神經(jīng)元內(nèi)出現(xiàn)的嗜酸性小體-路易小體(lewy body,LB),主要發(fā)生在黑質(zhì)致密部[1]。PD的發(fā)病機(jī)制主要是紋狀體DA的缺失使患者控制肌肉的能力受到限制;紋狀體DA含量的降低使膽堿能系統(tǒng)的功能發(fā)生障礙,這是產(chǎn)生PD各種運(yùn)動癥狀的生化基礎(chǔ)[2]。PD目前仍是臨床難以治愈的神經(jīng)退行性疾病,DA替代治療只能在最初給藥時(shí)緩解病人癥狀[3],因此尋找新的PD機(jī)制是可能根治疾病的方法。2001年,Magre等[4]首次在人類患者11號染色體上發(fā)現(xiàn)一種名為Berardinelli-Seip先天性脂肪代謝障礙2型(berardinelli-seip congenital lipodystrophy 2,BSCL2/Seipin)新基因的突變,該基因的突變起初被認(rèn)為與脂肪代謝障礙有關(guān)。BSCL2基因編碼的Seipin蛋白是一種位于內(nèi)質(zhì)網(wǎng)的跨膜蛋白,該蛋白在脂肪組織和中樞神經(jīng)系統(tǒng)中高表達(dá)[5-6],因此Seipin的功能研究也主要集中在這兩種組織病變中。近年研究發(fā)現(xiàn)Seipin基因突變與多種運(yùn)動神經(jīng)元疾病存在一些聯(lián)系,如腦內(nèi)Seipin的功能異常與運(yùn)動性神經(jīng)病和Silver綜合征有關(guān),小鼠神經(jīng)元丟失Seipin會導(dǎo)致焦慮和抑郁等[7-11]。這些研究結(jié)果提示Seipin異常可能是導(dǎo)致多種神經(jīng)退行性疾病的重要因素。目前已有研究表明,Seipin突變能引起內(nèi)質(zhì)網(wǎng)應(yīng)激蛋白CHOP的增加、進(jìn)而誘發(fā)細(xì)胞凋亡的發(fā)生[12]。因此,本實(shí)驗(yàn)主要研究對大鼠腎上腺嗜鉻細(xì)胞瘤(pheochromocytoma cells 12,PC12)細(xì)胞中Seipin基因進(jìn)行干擾,觀察Seipin缺陷后引起的細(xì)胞生物學(xué)變化,為進(jìn)一步研究帕金森疾病中神經(jīng)元的凋亡機(jī)制提供新的方向。
1 材料與方法
1.1材料、主要試劑及儀器
1.1.1材料 PC12細(xì)胞(中國科學(xué)院上海細(xì)胞庫),Seipin shRNA慢病毒、陰性對照慢病毒及慢病毒載體GV248(上海吉凱基因生物公司),RNAi靶序列為ACGCTCGGTGATGCTTCATTA,陰性對照靶序列為TTCTCCGAACGTGTCACGT。
1.1.2主要試劑與儀器 Dulbecco's modified eagle medium (DMEM) 培養(yǎng)基(美國Gibco公司),0.25%Trypsin-EDTA(美國Thermo Fisher公司),Penicillin Streptomycin雙抗(美國Life Technologies公司),Protease Inhibitor Cocktail、Protease/Phosphatase Inhibitor Cocktail、裂解液及苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)(美國CST公司),兔多克隆抗體BSCL2/Seipin(美國Abcam公司),4′,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI)、中性樹脂、Hochest凋亡染色試劑及TdT-mediated dUTP Nick-End Labeling (TUNEL)細(xì)胞凋亡檢測試劑盒(中國碧云天生物科技公司);倒置熒光顯微鏡(日本Nikon公司),曝光儀(美國Gene公司)。
1.2方法
1.2.1PC12細(xì)胞的培養(yǎng)與種板 PC12細(xì)胞使用含有10%胎牛血清和雙抗(青霉素和鏈霉素)的DMEM培養(yǎng)基于5%CO2的37 ℃培養(yǎng)箱中培養(yǎng),待細(xì)胞密度到達(dá)70%~80%時(shí),棄舊培養(yǎng)液,無菌PBS緩沖液加入漂洗,吸去PBS緩沖液;0.3 mL胰酶消化細(xì)胞加入培養(yǎng)基終止消化,巴氏吸管吹打貼壁細(xì)胞,將所有的液體轉(zhuǎn)入新的無菌15 mL Epp管中,1 000 r/min離心5 min,棄液,加入3 mL培養(yǎng)基重懸細(xì)胞并計(jì)數(shù);制備30個/L細(xì)胞懸液3 mL,取100 μL/孔加入96孔板用于慢病毒預(yù)感染實(shí)驗(yàn),取500 μL/孔加入48孔板用于慢病毒正式感染。
1.2.2慢病毒預(yù)感染實(shí)驗(yàn) 為了確定PC12細(xì)胞合適的感染復(fù)數(shù)(multiple of infection,MOI)及是否需要助感染試劑,按照不同培養(yǎng)條件將實(shí)驗(yàn)分為空白感染組(未加助感染劑)及助感染組(加助感染試劑HitansG),用不同MOI值的病毒感染細(xì)胞;DMEM培養(yǎng)基將shRNA慢病毒顆粒依次稀釋成病毒滴度為1×105TU/L(MOI=100),1×104TU/L (MOI=10)及1×103TU/L (MOI=1),待細(xì)胞生長至20%~30%,換掉舊培養(yǎng)基,加入培養(yǎng)基100 μL/組,對應(yīng)病毒滴度的慢病毒試劑90 μL和助感染試劑10 μL,混勻,繼續(xù)培養(yǎng);感染細(xì)胞10 h時(shí)吸走培養(yǎng)基,加入正常DMEM培養(yǎng)基,繼續(xù)培養(yǎng);細(xì)胞感染48~72 h,熒光顯微鏡觀察病毒感染的熒光強(qiáng)度,當(dāng)熒光強(qiáng)度達(dá)80%、且細(xì)胞生長正常組所對應(yīng)的感染條件和MOI值即為PC12細(xì)胞合適的最佳感染條件,記錄此時(shí)接種的細(xì)胞量、感染時(shí)的總體積、感染后換液的時(shí)間及細(xì)胞的MOI值。
1.2.3Seipin蛋白表達(dá)水平 采用Western blot檢測,依據(jù)上述獲得的感染條件,用陰性對照病毒和Seipin-shRNA目的病毒感染PC12細(xì)胞,分別作為陰性對照組與Seipin敲低組,同時(shí)設(shè)置正常組(正常PC12細(xì)胞);待細(xì)胞密度90%時(shí)用已配好的裂解液提取細(xì)胞總蛋白,與適量5×Buffer混勻,沸水煮沸變性蛋白,12% SDS-PAGE凝膠電泳(40 mA),用PVDF膜作為轉(zhuǎn)膜濾紙冰浴轉(zhuǎn)膜,200 mA、2 h;5%脫脂奶粉封閉1 h,TBST洗滌2次、5 min/次;膜用一抗孵育過夜,TBST洗滌3次、5 min/次;對應(yīng)二抗室溫孵育2 h,TBST洗滌3次、10 min/次;曝光儀獲取圖像,并用Image J計(jì)算蛋白條帶灰度值。
1.2.4細(xì)胞凋亡 采用TUNEL和Hoechst染色試劑盒檢測,將陰性對照組與Seipin敲低組細(xì)胞種于12孔板,待細(xì)胞密度達(dá)到60%~70%時(shí),用預(yù)冷的PBS洗滌細(xì)胞2次,5 min/次;4%多聚甲醛固定細(xì)胞20 min,含0.3%Triton X-100的PBS室溫孵育5 min,PBS洗滌細(xì)胞3次,分別用于TUNEL和Hoechst染色。前者系加入配好的TUNEL檢測液37 ℃孵育60 min,PBS洗滌細(xì)胞,DAPI封片,倒置熒光顯微鏡下觀察細(xì)胞染上紅光的情況;后者系加入0.5 mL Hoechst 33258染色液染色5 min,PBS洗滌細(xì)胞,中性樹脂封片,鏡下觀察細(xì)胞核出現(xiàn)致密濃染的程度。
1.3統(tǒng)計(jì)學(xué)分析
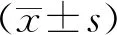
2 結(jié)果
2.1慢病毒預(yù)感染效果
熒光圖結(jié)果顯示,PC12細(xì)胞在MOI=1時(shí)無感染效果,助感染組與空白感染組相比感染效果無明顯差異;MOI=10時(shí)有較低感染效果,助感染組與空白感染組相比感染效果無明顯差異;MOI=100時(shí)感染效果最強(qiáng),助感染組PC12細(xì)胞感染效果高于空白感染組,見圖1。

圖1 2組PC12細(xì)胞的病毒感染效果(200×)Fig.1 Transfection efficiency of PC12 cells in the two groups(200×)
2.2Seipin蛋白敲低效果
Western blot結(jié)果顯示,與正常組相比,Seipin敲低組Seipin蛋白表達(dá)水平降低,差異有統(tǒng)計(jì)學(xué)意義(P<0.05),但陰性對照組Seipin蛋白表達(dá)水平差異無統(tǒng)計(jì)學(xué)意義(P>0.05),見圖2。
2.3TUNEL凋亡染色
TUNEL染色后結(jié)果顯示,與陰性對照組相比,Seipin敲低組紅色熒光的凋亡細(xì)胞明顯增加,見圖3。
2.4Hoechst凋亡染色
Hoechst染色結(jié)果顯示,與陰性對照組相比,Seipin敲低組細(xì)胞的細(xì)胞核致密濃染或碎塊狀致密濃染程度明顯增多,見圖4。

注:A為Western blot目的條帶,B為Seipin蛋白灰度條圖;(1)與正常組比較,P<0.05。圖2 各組PC12細(xì)胞Seipin蛋白表達(dá)Fig.2 The expression of Seipin in PC12 cells of the different groups

圖3 TUNEL法檢測PC12細(xì)胞凋亡(200×)Fig.3 TUNEL assay for apoptosis in PC12 cells(200×)

陰性對照組 Seipin敲低組圖4 Hoechst法檢測PC12細(xì)胞凋亡(200×)Fig.4 Hoechst assay for apoptosis in PC12 cells(200×)
3 討論
PD又叫震顫性麻痹,隨著年齡的增長,發(fā)病率呈逐年上升,給家庭和社會都帶來很大的經(jīng)濟(jì)負(fù)擔(dān),老年人的生活質(zhì)量也受到了影響,在臨床診斷上表現(xiàn)為DA能神經(jīng)元的退行性丟失以及神經(jīng)元內(nèi)出現(xiàn)的LB,其治療局限于預(yù)防對癥治療,不能有效地阻斷疾病的進(jìn)展[2]。PD的病因和發(fā)病機(jī)制十分復(fù)雜,至今未明,可能是氧化應(yīng)激過度、線粒體功能障礙、蛋白質(zhì)錯誤折疊和聚集等多種機(jī)制,最終通過多種級聯(lián)反應(yīng)導(dǎo)致DA能神經(jīng)元變性死亡;而歷年來的研究表明內(nèi)質(zhì)網(wǎng)應(yīng)激在PD發(fā)病過程起著重要作用[3]。Seipin/BSCL2于2001年首次被確定為BSCL2的致病基因[4],存在于內(nèi)質(zhì)網(wǎng)的完整膜上,目前預(yù)測其有2個跨膜結(jié)構(gòu)域,一個是腔內(nèi)環(huán),另一個是胞質(zhì)內(nèi)氨基和羧基末端[5];Seipin對脂肪形成起著重要作用,其功能缺陷可以嚴(yán)重?fù)p害脂肪細(xì)胞穩(wěn)態(tài),并導(dǎo)致脂肪代謝障礙[13]。現(xiàn)在有研究顯示Seipin缺陷也有潛在的神經(jīng)受累[14-17],如PD、Alzheimer Disease及Celia's Encephalopathy 等疾病中[18-20]。因此,Seipin可能是一種調(diào)節(jié)神經(jīng)元疾病的關(guān)鍵分子,在神經(jīng)退行性疾病中Seipin的生物學(xué)功能研究是該類疾病研究的新方向。
為探討Seipin蛋白表達(dá)降低后對細(xì)胞凋亡的影響,本研究采用神經(jīng)退行性疾病模型細(xì)胞PC12,用基因干擾技術(shù)下調(diào)其Seipin蛋白的表達(dá)。預(yù)感染試驗(yàn)結(jié)果顯示,PC12細(xì)胞在MOI=1時(shí)無感染效果,助感染組與空白感染組相比感染效果無明顯差異;MOI=10時(shí)有較低感染效果,助感染組與空白感染組相比感染效果無明顯差異;MOI=100時(shí)感染效果最強(qiáng),助感染組PC12細(xì)胞感染效果高于空白感染組,提示PC12細(xì)胞用MOI=100計(jì)算病毒量來感染,助感染試劑能增加感染效率。Western blot結(jié)果顯示,與正常組相比,Seipin敲低組Seipin蛋白水平降低,差異有統(tǒng)計(jì)學(xué)意義(P<0.05),陰性對照組Seipin蛋白水平差異無統(tǒng)計(jì)學(xué)意義(P>0.05),提示成功構(gòu)建Seipin敲低細(xì)胞,陰性對照細(xì)胞與正常細(xì)胞無差異。TUNEL染色后熒光圖結(jié)果表明,與陰性對照組相比,Seipin敲低組顯示紅色熒光的凋亡細(xì)胞明顯增加,說明Seipin敲低后細(xì)胞發(fā)生凋亡的水平增加。Hoechst染色結(jié)果顯示,與陰性對照組相比,Seipin敲低組細(xì)胞的細(xì)胞核致密濃染或碎塊狀致密濃染程度明顯增多,正常細(xì)胞的細(xì)胞核經(jīng)染色后會呈淡藍(lán)色,凋亡細(xì)胞的細(xì)胞核會呈致密濃染,或呈碎塊狀致密濃染,顏色有些發(fā)白[21],說明Seipin蛋白敲低后細(xì)胞凋亡增加。以往研究顯示細(xì)胞中Seipin發(fā)生突變會導(dǎo)致內(nèi)質(zhì)網(wǎng)中錯誤折疊的蛋白質(zhì)聚集,從而引發(fā)內(nèi)質(zhì)網(wǎng)應(yīng)激,最終導(dǎo)致錯誤折疊蛋白反應(yīng)和細(xì)胞死亡[22];體內(nèi)實(shí)驗(yàn)結(jié)果也表明,小鼠敲除Seipin后,隨著年齡的增長,運(yùn)動發(fā)生明顯障礙且有運(yùn)動神經(jīng)元的丟失[23]。這些實(shí)驗(yàn)結(jié)果表明Seipin的缺乏可以導(dǎo)致細(xì)胞維持正常的生物學(xué)功能受到影響,從引起細(xì)胞凋亡的發(fā)生。
綜上所述,本研究成功構(gòu)建大鼠PC12 Seipin敲低細(xì)胞,下調(diào)Seipin蛋白會增加細(xì)胞凋亡。
猜你喜歡
中老年保健(2021年12期)2021-11-30 02:58:01
好日子(2021年8期)2021-11-04 09:02:46
小學(xué)生學(xué)習(xí)指導(dǎo)(爆笑校園)(2020年6期)2020-07-03 10:01:10
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
攝影之友(影像視覺)(2018年12期)2019-01-28 09:01:02
攝影之友(影像視覺)(2018年12期)2019-01-28 09:01:02
中華詩詞(2018年11期)2018-03-26 06:41:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年11期)2017-10-23 01:32:36
Coco薇(2016年8期)2016-10-09 02:11:50
中國醫(yī)藥科學(xué)(2015年19期)2015-02-27 12:33:11
