云南松3種切梢小蠹梢轉干期競爭共存的時空生態位*
2020-04-28 07:10:16武承旭孔祥波張蘇芳
林業科學 2020年3期
關鍵詞:生態
武承旭 劉 福 孔祥波 張蘇芳 張 真
(1. 中國林業科學研究院森林生態環境與保護研究所 國家林業和草原局森林保護學重點實驗室 北京 100091; 2. 貴州大學林學院 貴陽 550025)
橫坑切梢小蠹(Tomicusminor)、云南切梢小蠹(T.yunnanensis)和短毛切梢小蠹(T.brevipilosus)屬鞘翅目(Coleoptera)小蠹科(Scolytidae)切梢小蠹屬(Tomicus),三者混合發生,協同危害,已成為云南松(Pinusyunnanensis)的重要害蟲(L?ngstr?m, 1983; Ye, 1991; Yeetal., 1997; Kirkendalletal., 2008)。3種切梢小蠹對云南松的危害主要包括蛀梢和蛀干(葉輝, 1996):每年5—11月為蛀梢期,樹干中新羽化的成蟲通過蛀食枝梢補充營養(Yeetal., 1999; L?ngstr?m, 1983; Lüetal., 2014),危害周期長,造成云南松樹梢枯黃易折,樹勢衰弱(Ye, 1991; Lüetal., 2014); 11月至翌年4月為蛀干期,初期成熟切梢小蠹首先大量聚梢(L?ngstr?metal., 2002),然后轉移到樹干產卵繁殖,在云南松韌皮部蛀食形成母坑道,子代取食韌皮部形成子坑道,是導致寄主死亡的直接原因(L?ngstr?m, 1983; Lieutieretal., 2003; Lüetal., 2014)。
在小蠹對寄主樹木的選擇和入侵危害過程中,其種內或種間常常由于有限的空間和營養資源存在競爭和共存關系(蒲曉娟等, 2007),并在林間通過競爭相互適應(Yeetal., 1999; 葉輝, 1997)。在空間資源利用方面,橫坑切梢小蠹主要蛀食云南松樹干中、下部,而云南切梢小蠹主要蛀食云南松樹干中、上部(Yeetal., 1999; Luetal., 2015)。在時間資源利用方面,云南切梢小蠹一般11月開始由梢部轉移到樹干部位危害,即蛀梢末期或蛀干初期,至翌年4月(葉輝, 1996),而橫坑切梢小蠹梢轉干期從每年11月開始至翌年3月,比云南切梢小蠹為害期晚約1~2周(葉輝等, 2004); 也有研究表明,云南切梢小蠹、橫坑切梢小蠹和短毛切梢小蠹的蛀梢時間分別起于4、5和6月,至翌年2、4和5月(賈平等, 2014; Lüetal., 2014)。這說明云南切梢小蠹為初期性種類(先鋒種),首先大量聚梢危害,為其他次生性小蠹(如橫坑切梢小蠹)開辟資源利用空間。此外,3種切梢小蠹中橫坑切梢小蠹適應云南切梢小蠹的分布規律而分布(葉輝, 1997; Yeetal., 1999),但其中競爭和共存機制還不清楚。
生態位理論反映物種對生態資源的占據和利用情況以及物種間的競爭關系(孫儒泳, 2001; 徐汝梅, 1987),目前通常利用時間和空間生態位的寬度、重疊關系以及相似性比例和種間競爭系數來研究某一群落中物種間的競爭共存機制(陳輝等, 1999; 劉麗等, 2007; 路紀芳等, 2017)。研究發現,小蠹利用種群密度大小、入侵時間以及空間資源的多樣性差異來平衡種間和種內的競爭和共存(陳輝等, 1999; 劉麗等, 2007);蛀干后期小蠹密度大時,其幼蟲生態位重疊與樹脂分布體積關系密切(Kimberlyfetal., 2008); 小蠹家族種間也利用異種信息素的相互作用增加或減少生態位重疊(Ayresetal., 2001)。Paine等(1981) 研究北美西南部同域危害火炬松(Pinustaeda)的南部松大小蠹(Dendroctonusfrontalis)和其他3種齒小蠹屬(Ips)昆蟲發現,物種多樣性與生態位重疊值存在正相關關系。關于其他蛀干害蟲的生態位也有零星報道,如興安落葉松(Larixgmelinii)中落葉松八齒小蠹(I.subelongatus)與3種優勢天牛因取食分化而實現共存(袁菲等, 2011);宗世祥等(2005)從時空生態位角度定量描述了沙棘木蠹蛾(Holcocerushippophaecolus)與紅緣天牛(Asiashalodendri)種群之間的聯系和競爭共存機制。
研究物種時空生態位及其分布規律,既可了解種間競爭關系,還可為綜合防治提供科學依據(宗世祥等, 2005)。截至目前,從種群生態位角度探討同域、同寄主的3種切梢小蠹種間競爭共存關系未見報道。鑒此,本研究采用抽樣調查法,根據生態位理論,從時間和空間資源利用上對3種切梢小蠹在云南松樹冠和樹干中的分布規律及三者雌雄之間的競爭共存關系進行研究,為種群監測綜合防治提供理論基礎。
1 試驗地概況與研究方法
1.1 試驗地概況
試驗地位于云南省大理市祥云縣普棚鎮(25°20′25.96″N,100°54′31.43″E),海拔2 250 m,為人工營造的云南松純林,約20 hm2,樹齡20年左右,郁閉度0.5~0.6。氣候屬干、濕季節分明的北亞熱帶高原季風型中溫帶氣候,5—9月為雨季,降水集中; 10月至翌年4月為干季,光照充足。常年主要風向為西南風。調查期間(2016年10月至次年5月)平均氣溫13.6 ℃,降水較少,試驗期間試驗地未噴灑農藥。該地常年受橫坑切梢小蠹、云南切梢小蠹和短毛切梢小蠹混合危害,其中短毛切梢小蠹數量占前兩者的1%。
1.2 種群空間分布及生態位調查
2016年10月下旬至次年4月上旬,每隔1個月左右,在1 hm2切梢小蠹危害的云南松標準地內,采用五點取樣法隨機抽取15株云南松,每株云南松按照上、中、下3個層次隨機抽取25枝樹梢,詳細記錄其在樹冠中不同分布區間的總梢數、有蟲梢數。將有蟲梢裝入5 mL帶孔離心管,帶回實驗室,在解剖鏡下根據形態特征鑒定切梢小蠹的種類和性別(李霞等, 2012; 王平彥等, 2015),并統計數量。共調查5次。同時,每隔1個月左右,在標準地內選取樹干上有切梢小蠹新蛀孔的云南松4株,砍成1 m小段,逐段解剖,查看切梢小蠹的數量,蛀干后期根據坑道類型判斷切梢小蠹的種類,并將蛀干切梢小蠹帶回實驗室鑒定種類和性別。調查地云南松樹高為4.3~11 m,胸徑為7~15 cm。
1.3 數據分析方法
1.3.1 切梢小蠹種群生態位資源劃分 根據時間和空間資源序列,對梢轉干期樹梢和轉移到樹干入侵的3種切梢小蠹數量進行生態位資源劃分。時間資源生態位劃分: 2016年10月底至次年4月初,蛀梢和蛀干切梢小蠹時間生態位均以月為單位劃分資源單元。 空間資源生態位劃分: 2016年11月至次年4月,蛀梢切梢小蠹空間生態位以樹冠上、中、下3層作為空間資源序列進行劃分; 蛀干切梢小蠹空間生態位依據樹干高度1 m劃分。
1.3.2 計算公式 1) 生態位寬度和生態位重疊 采用Levins(1968)和徐汝梅(1987)的測定公式計算3種切梢小蠹的時空生態位寬度和生態位重疊,以表明某種生物資源在不同數量單元資源序列上的分布情況及對資源的利用情況。
生態位寬度公式如下:
式中:B為物種的生態位寬度;S為每個資源集合中的總單元數;Pi為一個物種在資源序列第i單元[i分別為橫坑切梢小蠹(雌、雄)、云南切梢小蠹(雌、雄)和短毛切梢小蠹(雌、雄),下同]中所占的比例。
本研究在梢轉干期樹冠中調查持續5個月,其時間資源集合S為5; 空間資源上分上、中、下3層,資源集合S為3。蛀干期連續調查4次,時間資源集合S為4;將樹干截成5~7段,空間資源集合S為5~7。
生態位重疊公式如下:
式中:aij為第i個物種對物種j的生態位重疊;Pih、Pjh分別為第i和第j個物種在資源集第h單元中的比例;Bi為第i個物種的生態位寬度。
2) 生態位相似性系數 采用Levins(1968)和Colwell等(1977)的測定公式計算切梢小蠹的生態位相似性系數,反映2個物種利用資源的相似程度:
式中: PS為物種i和物種j的生態位相似性系數;Pij、Pjh分別為第i個物種和第j個物種在資源集第h單元中的數量占總資源的比例,即在資源單元h中第i和j個物種所占的比例。
當2個物種間的生態位相似系數小于0.7時,則認為在某種資源利用上物種隔離(Hutchinson, 1959)。
3) 種間競爭系數 采用May (1975)的測定公式計算切梢小蠹之間的種間競爭系數,反映2個物種對資源的競爭激烈程度:
式中:α為種間競爭系數;Pi、Pj分別為第i個物種和第j個物種在各資源序列中的比例。
1.3.3 數據分析 試驗所得數據在Excel 2010中進行整理,采用SPSS 19.0數據處理系統進行分析。梢轉干期樹冠和樹干中3種切梢小蠹雌、雄蟲在不同調查時間和樹冠上、中、下3層的分布比例采用費舍爾精準檢驗(Fisher exact test)方法。不同調查期橫坑切梢小蠹和云南切梢小蠹在樹干不同高度分布比例以及2種切梢小蠹在不同調查期分布比例的差異顯著性采用卡方檢驗(Chi-square test)進行分析。橫坑切梢小蠹和云南切梢小蠹種群蟲口密度在樹干不同高度的差異顯著性經單因素方差分析(one-way ANOVA)后采用Turkey’s HSD檢驗進行多重比較。3種切梢小蠹雌、雄蟲梢轉干期在樹冠和樹干中的時空生態位寬度、生態位重疊、生態位相似性系數和生態位種間競爭系數采用Excel 2010整理計算。
2 結果與分析
2.1 梢轉干期樹冠中3種切梢小蠹成蟲時間生態位
梢轉干期,樹冠中橫坑切梢小蠹和云南切梢小蠹雌、雄蟲所占比例隨著時間推移均表現出先增加后減少的規律;橫坑切梢小蠹雌、雄高峰期均晚于云南切梢小蠹,短毛切梢小蠹無明顯高峰期;調查初期,云南松樹冠中橫坑切梢小蠹和云南切梢小蠹雌蟲蟲口數量明顯多于雄蟲,01-07后則相反,短毛切梢小蠹沒有顯示明顯隨時間變化的規律;梢轉干期3種切梢小蠹雌、雄蟲在不同時間段差異顯著(P< 0.001,n=225)(圖1)。
橫坑切梢小蠹雌、雄蟲的時間生態位寬度高于云南切梢小蠹小蠹和短毛切梢小蠹,且云南切梢小蠹雌、雄蟲的時間生態位寬度低于短毛切梢小蠹,3種切梢小蠹雌蟲的時間生態位寬度均低于雄蟲(表1)。
3種切梢小蠹在樹冠中相互競爭共存,關系緊密,其中橫坑切梢小蠹雌蟲對云南切梢小蠹雌蟲的時間生態位重疊最大(0.203 3),相互間對時間資源的共享最多,但是橫坑切梢小蠹雌蟲與云南切梢小蠹雄蟲的生態位相似性系數最大(0.945 2),相互間競爭激烈; 橫坑切梢小蠹雄蟲對云南切梢小蠹雌、雄蟲的時間生態位重疊分別為0.201 1和0.199 0,均高于短毛切梢小蠹雌、雄蟲;橫坑切梢小蠹和云南切梢小蠹之間的時間生態位相似性系數均高于二者與短毛切梢小蠹之間的時間生態位相似性系數,且短毛切梢小蠹雌、雄蟲與云南切梢小蠹雌蟲的時間生態位相似性系數低于0.7,趨于分離,各自不構成時間資源上的競爭,短毛切梢小蠹雄蟲與橫坑切梢小蠹雌蟲在時間資源上也趨向分離(表1)。
橫坑切梢小蠹雌蟲對云南切梢小蠹雄蟲的種間競爭系數最大(0.993 1),表明者之間競爭最激烈,但對短毛切梢小蠹雌雄蟲的生態位種間競爭系數相對較小(表2)。

圖1 梢轉干期云南松每25梢切梢小蠹蟲口密度隨時間變化情況對比Fig.1 Comparison of population densities of Tomicus species in the crowns each 25 shoots on P. yunnanensis with time
2.2 梢轉干期樹冠中3種切梢小蠹成蟲空間生態位
梢轉干期,樹冠中未蛀干橫坑切梢小蠹雌、雄蟲所占比例在樹冠不同層次由上到下依次增多,而云南切梢小蠹和短毛切梢小蠹相反,在樹冠上層分布最多。3種切梢小蠹雌、雄蟲在樹冠不同層次差異不顯著(P=1.000,n=18)。
橫坑切梢小蠹雌、雄蟲的空間生態位寬度均小于云南切梢小蠹,但大于短毛切梢小蠹;云南切梢小蠹和短毛切梢小蠹雌蟲的生態位寬度均小于雄蟲,但橫坑切梢小蠹兩性的空間生態位寬度相反(表1)。
橫坑切梢小蠹雌蟲對其雄蟲的空間生態重疊相對于云南切梢小蠹和短毛切梢小蠹雌、雄蟲生態位重疊最大,其中橫坑切梢小蠹雌蟲對短毛切梢小蠹雄蟲的空間生態位重疊最小;橫坑切梢小蠹雄蟲對云南切梢小蠹雌、雄蟲的空間生態位重疊分別為0.326 5和0.326 4,二者對空間資源的利用相對接近,而云南切梢小蠹雌蟲對短毛切梢小蠹雌蟲的生態位重疊最大(表1)。
2種切梢小蠹性別間的空間生態位相似性系數均大于0.7,各自構成對空間資源的競爭(表1)。其中云南切梢小蠹雌蟲對其雄蟲的生態位相似性系數最大為0.985 7,其次是橫坑切梢小蠹雌蟲對其雄蟲的空間生態位相似性系數,而橫坑切梢小蠹對短毛切梢小蠹雌蟲的生態位相似性系數最小。云南切梢小蠹雌蟲和其雄蟲的生態位種間競爭系數最大,橫坑切梢小蠹雌蟲和其雄蟲之間的種間競爭系數次之,云南切梢小蠹雌蟲和短毛切梢小蠹雌蟲之間的種間競爭系數最小(表2)。
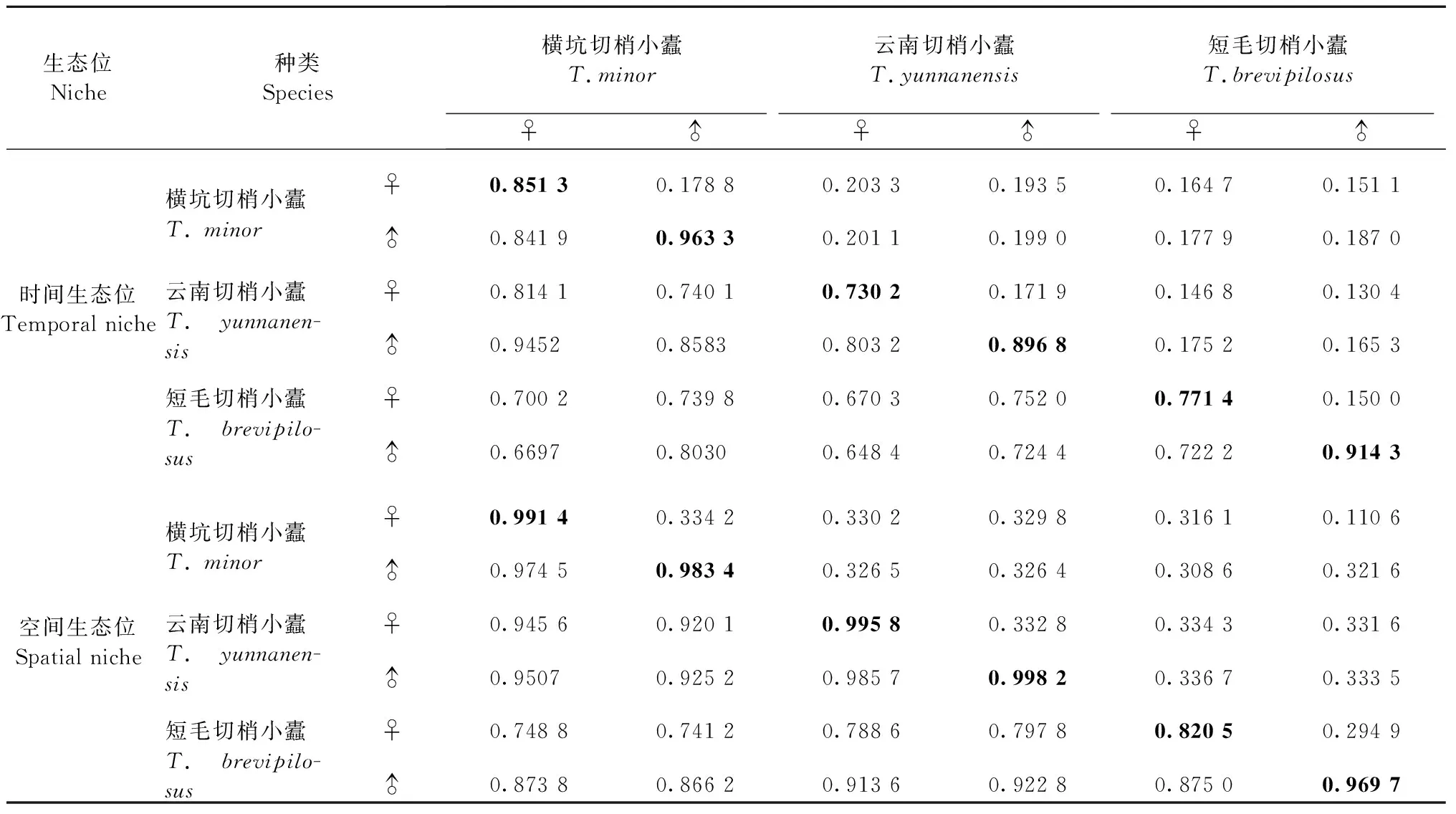
表1 蛀干期樹冠中3種切梢小蠹的時空生態位寬度、生態位重疊和生態位相似性系數①Tab.1 Temporal- and spatial-niche breadth, overlap and proportion similarity of three Tomicus species in the crown of P. yunnanensis during the trunk-feeding phase
① 主對角線加粗數據為蛀干期云南松樹梢中3種切梢小蠹的生態位寬度,右上方為生態位重疊,左下方為生態位相似性系數。The data with bold in diagonal line are temporal- or spatial-niche breadth of threeTomicusspecies in the shoots ofP.yunnanensisduring the trunk-feeding phase, while which in upper-right and lower-left are respectively temporal niche overlap and proportion similarity.

表2 蛀干期樹冠中3種切梢小蠹的種間競爭系數Tab.2 Interspecific competition coefficient of three Tomicus species in the crown of P. yunnanensis during the trunk-feeding phase
2.3 梢轉干期樹干中2種切梢小蠹的分布及其時空生態位
梢轉干期,在云南松樹干中未發現短毛切梢小蠹侵入,因此蛀干期樹干中僅研究橫坑切梢小蠹和云南切梢小蠹的分布及共時空生態位。切梢小蠹蛀干期以母坑道數量為依據,不考慮性別差異。
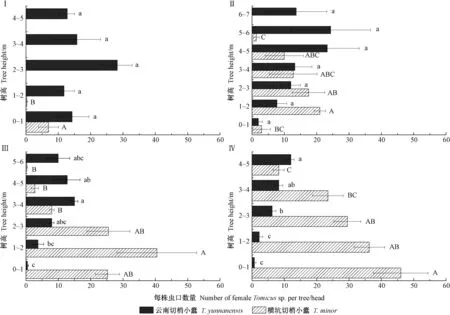
圖2 蛀干期2種切梢小蠹在不同調查時期不同樹干高度的蟲口密度情況對比Fig.2 Comparsion of population densities of T. minor and T. yunnanensis infested in the stem ofP. yunnanensis during the trunk-feeding phaseI—IV:蛀干期調查的日期分別為12-06、01-07,02-23和04-05 Represent the survey date 12-06, 01-07, 02-23 and 04-05,respectively. 不同大、小寫字母分別表示橫坑切梢小蠹和云南切梢小蠹種群蟲口密度在樹干不同高度經單因素方差分析后Turkey 檢驗在0.05水平上差異顯著。The capital and lowercase letter in the figure respectively represent the significant difference at 0.05 level of population densities of T. minor and T. yunnanensis in the different heights of stem from P. yunnanensis.
12月初,橫坑切梢小蠹主要分布在云南松樹干2 m以下,隨著時間推移,橫坑切梢小蠹逐步向樹干上部入侵并蛀食母坑道,母坑道數量隨著樹干高度增加逐漸減少(圖2)。不同調查時期,橫坑切梢小蠹在樹干不同高度差異顯著(12-06: df=4,F=92.953,P=0.01; 01-07: df =6,F=3.083,P=0.028; 02-23: df =5,F=7.104,P=0.001; 04-05: df =4,F=7.393,P=0.002)。云南切梢小蠹在入侵初期可攻擊樹干任何部位,形成母坑道;蛀干初期入侵云南松樹干中部所占比例最大,為34.24%; 隨著時間推移,攻擊樹干上部形成的母坑道逐漸增多,如12-06,上部占40%; 12-06和01-07在樹干不同高度差異不顯著(df =4,F=1.918,P=0.160; df =6,F=1.394,P=0.271)(圖2Ⅰ、Ⅱ)。02-23和04-05,云南切梢小蠹種群密度在樹干不同高度差異顯著(df =5,F=5.531,P=0.003; df =4,F=18.515,P<0.001)(圖2Ⅲ、Ⅳ)。在不同調查時期,橫坑切梢小蠹和云南切梢小蠹在樹干不同位置均差異顯著:12-06(df =4,χ2=92.953,P<0.001); 01-01(df =6,χ2=144.000,P<0.001); 02-23(df =5,χ2=287.400,P<0.001)和04-05(df =4,χ2=154.400,P<0.001);且2種切梢小蠹在調查期間不同月份差異顯著(df =3,χ2=600.500,P<0.001)。
橫坑切梢小蠹對云南切梢小蠹的時間生態位重疊小于云南切梢小蠹對橫坑切梢小蠹,二者的時間生態位相似性系數小于0.7(0.556 1),存在空間生態位隔離; 且二者的種間競爭系數為0.237 9(圖3)。橫坑切梢小蠹梢轉干期的時間生態位寬度隨著時間推移逐漸增加,而云南切梢小蠹相反(圖4)。
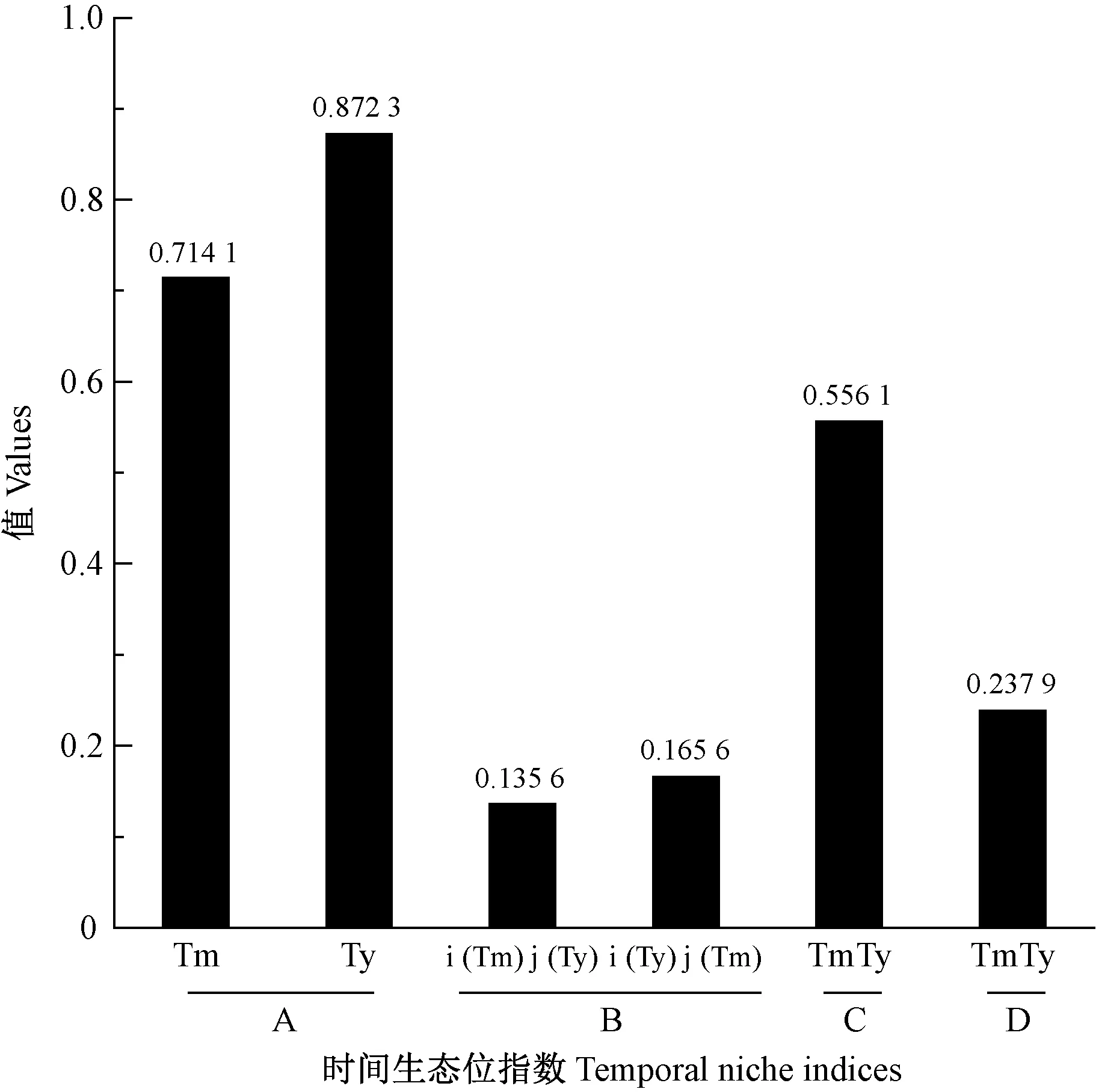
圖3 蛀干期切梢小蠹在云南松樹干中的時間生態位指數Fig.3 The values of temporal niche indices for Tomicus species on the stems of P. yunnanensis during the trunk-feeding phaseTm:橫坑切梢小蠹T. minor;Ty:云南切梢小蠹T. yunnanensis; TmTy:二者相互作用Interaction relationship between these two species; i (Tm) j (Ty):橫坑切梢小蠹對云南切梢小蠹的生態位重疊The niche overlap of species i to species j.下同The same below.A—D:分別代表生態位寬度、生態位重疊、生態位相似性系數和種間競爭系數Represent niche breadth, niche overlap, niche proportion similarity and inlerspecific competition coefficient,respectively.
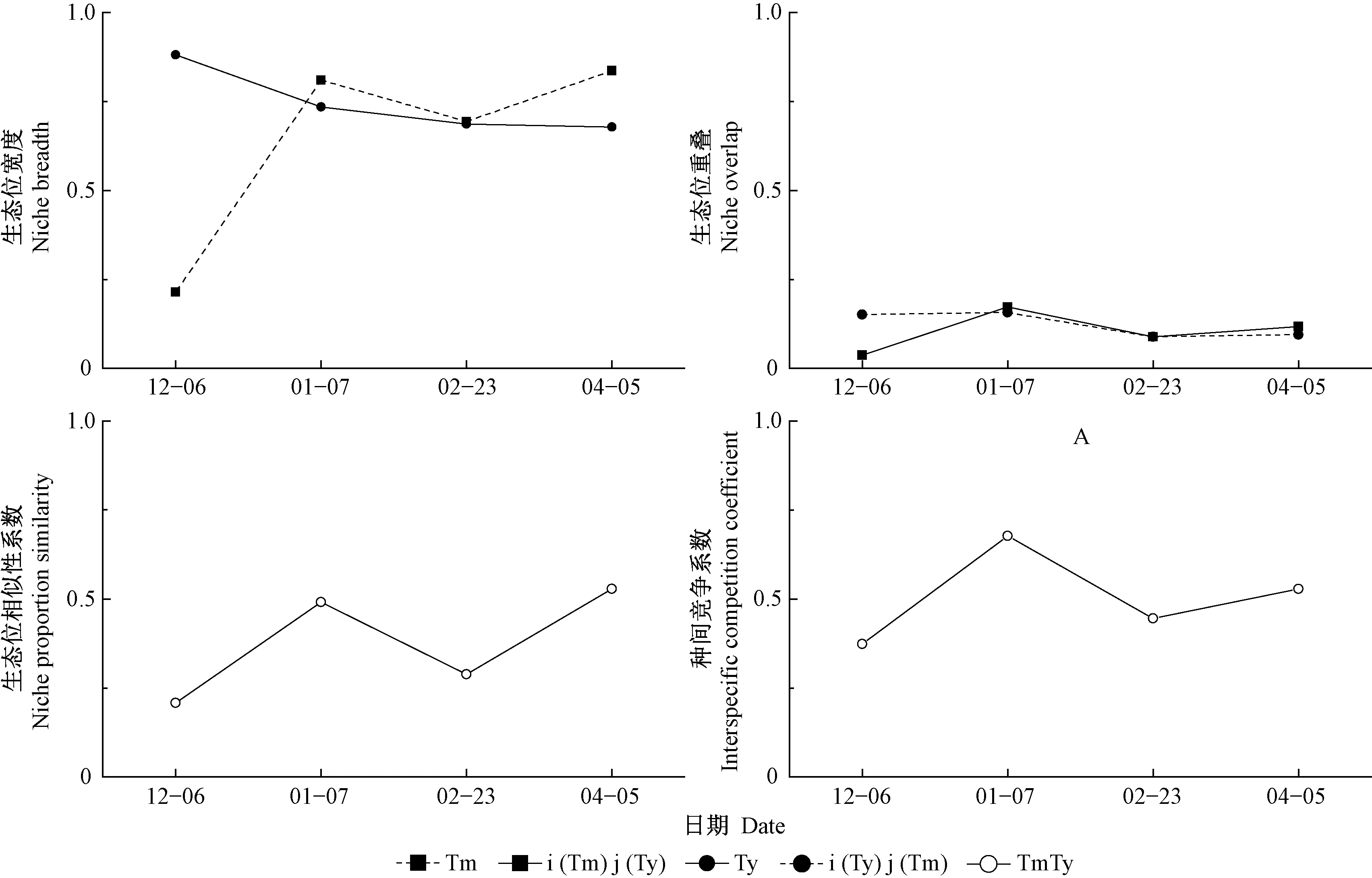
圖4 蛀干期切梢小蠹在云南松樹干中的空間生態位指數Fig.4 The values of spatial niche indices of Tomicus species on the stems of P. yunnanensis during the trunk-feeding phase
云南切梢小蠹的空間生態位寬度隨著時間推移逐漸降低,而橫坑切梢小蠹則增加,在蛀干后期增加到最大,為0.837 0(圖3)。在01-10,二者的空間生態位重疊最大(0.172 9),空間生態位相似性系數和種間競爭系數均最大。
3 討論
3.1 分布規律
綜合切梢小蠹在云南松樹冠和樹干中的分布比例和時空生態位可以發現,云南切梢小蠹作為初期性害蟲在梢轉干初期首先聚梢危害(Lieutieretal., 2003),然后入侵整個樹干,為其他次生性切梢小蠹(如橫坑切梢小蠹)開辟資源利用空間。橫坑切梢小蠹晚于云南切梢小蠹入侵后,通過空間上的重疊和競爭逐漸擴張利用資源,與Lü等(2014)研究結果相似,類同于青海云杉(Piceacrassifolia)天然林中云杉大小蠹(Dendroctonusmicans)為先鋒種入侵健康木,東方擬齒小蠹(Pseudipsorientalis)和光臀八齒小蠹(I.nitidus)為次生性小蠹入侵剩余資源(劉麗等, 2007)。這些結果說明,時序模式是2種切梢小蠹重要的影響因素,不同小蠹需根據自身進化和生物學特征,判斷恰當的時間入侵適合的寄主。本研究中短毛切梢小蠹種群數量較小,僅在樹梢中發現,且在不同調查時期種群數量無差異。云南松樹冠中橫坑切梢小蠹和云南切梢小蠹雌性蟲口數量隨著時間推移先增加后減少,且高峰期明顯早于雄蟲,這與切梢小蠹屬昆蟲雌性首先攻擊蛀干形成母坑道的行為有關(L?ngstr?m, 1983)。在樹冠不同層次,橫坑切梢小蠹大量聚集在樹冠下層,而云南切梢小蠹和短毛切梢小蠹則在樹冠上層分布最多,與前人的報道相同(葉輝等, 1994; 葉輝, 1996)。在云南松樹干上,橫坑切梢小蠹蛀干初期主要分布在樹干中、下層(2 m以下),與前人的研究結果接近(Yeetal., 1999; Luetal., 2015)。云南切梢小蠹蛀干初期可分布于整個樹干,并隨著橫坑切梢小蠹競爭呈現出向樹干上部轉移的動態變化情況,與Ye等(1999)和Lu等(2015)報道的云南切梢小蠹主要分布在樹干中上部基本相同,但與其他小蠹屬昆蟲不同,如南部松大小蠹、I.avulsus和I.calligraphus可能短期內先入侵樹干中部,再向兩側擴散(Svihraetal., 1980),這均反映出同一生態系統中小蠹選擇寄主入侵部位時存在一定的競爭與共存策略。調查期間,在寄主樹干中未曾發現入侵短毛切梢小蠹,可能是種群數量較小、蛀干木太少的緣故,因此目前結果無法評價短毛切梢小蠹與其他2種切梢小蠹的競爭與共存關系; 然而Chen等(2015)發現,在云南松上的短毛切梢小蠹存在2種侵入模式,初期侵入橫坑切梢小蠹和云南切梢小蠹未蛀食空間,后期在沒有前二者存在競爭的情況下從樹干下部向上侵入。
3.2 時空生態位寬度
分布在樹冠中的3種切梢小蠹,短毛切梢小蠹的時空生態位寬度最小,橫坑切梢小蠹和云南切梢小蠹通過對時間和空間資源的分配和彌補達到種群共存和同源競爭,與陳輝等(1999)研究秦嶺華山松(Pinusarmandi)11種小蠹的生態位相似,其發現11種小蠹利用不同營養和空間需求、種群密度大小以及入侵時間差異來平衡種間和種內的競爭與共存; 而與青海云杉天然林中6種小蠹的生態位系統不同,其是利用空間資源多樣性來達到種間競爭的平衡和共存(劉麗等, 2007)。這說明,不同生態系統中小蠹之間的競爭與共存機制并不存在共性,小蠹對生存環境的資源競爭、適應與其生物學、寄主等因素有很大關系,諸多因素決定了小蠹種間競爭與共存策略的多樣性和復雜性。
樹冠中3種切梢小蠹兩性間在時間和空間生態位寬度上也存在差異,3種切梢小蠹雌蟲的時間生態位寬度以及云南切梢小蠹和短毛切梢小蠹雌蟲的空間生態位寬度均低于其雄蟲,這可能是因為切梢小蠹雌蟲短期內大量聚梢危害,之后快速轉移到樹干蛀食坑道(L?ngstr?metal., 1993; Lieutieretal., 2003; 葉輝等, 2004),進而降低了寄主云南松的抗性(Lüetal., 2014),也印證了云南松樹冠中橫坑切梢小蠹和云南切梢小蠹雌蟲的高峰期明顯早于雄蟲這一結論。3種切梢小蠹中橫坑切梢小蠹的時間生態位寬度最大,但其空間生態位寬度卻小于云南切梢小蠹,這說明梢轉干期云南切梢小蠹占據更大空間資源,但時間資源上卻少于橫坑切梢小蠹,也印證了橫坑切梢小蠹梢轉干高峰略晚于云南切梢小蠹的結論,基于時間和空間資源的分配實現了該生態系統中3種切梢小蠹的競爭與共存。此外,橫坑切梢小蠹雄蟲的空間生態位寬度均低于雌蟲,與其他2種切梢小蠹的空間生態位寬度剛好相反,但雌蟲的時間生態位卻小于雄蟲,這說明較其他2種切梢小蠹雌雄成蟲而言,橫坑切梢小蠹雌成蟲對空間資源的需求大于對時間資源的需求。從3種切梢小蠹的空間生態位、時間生態位在種間、種內雌雄個體的差異來看,該生態系統的資源分配機制較為復雜,特別是性別間的差異還需要進一步研究探討。
從切梢小蠹在樹干中的空間生態位來看,云南切梢小蠹的空間生態位寬度隨著時間推移逐漸降低,與橫坑切梢小蠹正好相反,反映出云南切梢小蠹占據云南松樹干的空間隨著時間推移受橫坑切梢小蠹入侵的競爭縮小,說明2種切梢小蠹在樹干空間資源上相互競爭和抑制并維持共存,進一步證明了云南切梢小蠹較橫坑切梢小蠹是一個先鋒種,橫坑切梢小蠹屬于次生性害蟲(葉輝等, 2004)。
3.3 時空生態位重疊
生態位重疊與昆蟲間潛在的競爭有關(Levins, 1968),能夠反映同域種競爭的情況。本研究中橫坑切梢小蠹雌蟲與云南切梢小蠹雌蟲的時間生態位重疊最大,且對云南切梢小蠹雌雄蟲的生態位重疊均高于對短毛切梢小蠹雌雄的生態位重疊,說明橫坑切梢小蠹和云南切梢小蠹對時間資源的利用更高。這可能是調查樣地中橫坑切梢小蠹和云南切梢小蠹種群數量接近,而短毛切梢小蠹種群數量較少的原因。在空間生態位重疊上,橫坑切梢小蠹雌雄間的生態位重疊最大,說明橫坑切梢小蠹雌雄蟲在空間資源上跟隨作用明顯。3種切梢小蠹在寄主樹冠中長期的生態位重疊既可有效降低寄主的抗性,也可促進小蠹對寄主樹干的攻擊(Lüetal., 2014)。
4 結論
通過云南松林間調查,并結合前人研究,本研究明確了云南省祥云縣3種切梢小蠹梢轉干期在樹冠和樹干中的分布規律,同時發現同域3種切梢小蠹在寄主云南松樹冠和樹干中利用時間和空間資源的多層次性達到生態位的重疊和分離,進而實現種內和種間的競爭與共存,維持生態系統中同域切梢小蠹種群的穩定性。本研究結果可為3種切梢小蠹的綜合防治提供理論基礎,然而3種切梢小蠹聚梢和蛀干期競爭能力的強弱還需進一步證實,梢轉干的行為機制尚不清楚,還需從行為學、化學生態學等角度進一步研究三者對時空資源的分配問題。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
貴茶(2019年3期)2019-12-02 01:46:32
鄉村地理(2018年3期)2018-11-06 06:51:02
茶葉通訊(2017年2期)2017-07-18 11:38:40
河北城市研究(2015年4期)2015-08-23 11:53:06
浙江工商大學學報(2015年6期)2015-03-01 02:56:40
