長鏈非編碼RNA DRAIC調控miR-181通過STAT信號通路對卵巢癌遷移和侵襲的影響
2020-04-27 06:25:32李晗宇
轉化醫學雜志 2020年2期
李晗宇,陳 飛
卵巢癌是全世界女性中發病率較高且死亡率最高的一類婦科腫瘤性疾病,也是女性惡性腫瘤中相關死亡率最常見的原因[1]。盡管目前手術和化療治療方式都取得了一定的進展,但卵巢癌患者的總體存活率仍然很低[2]。最近的研究表明,許多長鏈非編碼RNA(long non-coding RNA,lncRNA)在多種細胞過程中都具有重要作用,如增殖,分化,凋亡,細胞生物行為以及疾病發病機制,導致對細胞基因調控網絡的影響,干擾其進程[3]。LncRNA DRAIC是一種廣泛表達的lncRNA,首次被發現是非小細胞肺癌中高轉移潛能和預后不良的因子[4]。最近,lncRNA DRAIC與多種人類腫瘤相關,包括肺癌、肝癌、腎癌、結直腸癌、胃癌和乳腺癌[5]。生物信息學分析表明lncRNA DRAIC與miR-181(一種有效的腫瘤抑制因子)結合,然后靶向miR-181的表達[6]。本研究探討lncRNA DRAIC調控miR-181通過STAT信號通路對卵巢癌遷移和侵襲的影響。
1 材料和方法
1.1 細胞株 人卵巢癌細胞株SKOV3、COC1、CAOV3和A2780購買于中國科學院上海分院。卵巢癌細胞株均培養在含10%胎牛血清和1%雙抗的RPMI-1640培養基中,所有細胞均在5% CO2,37 ℃條件下培養,每隔3 d換液1次,待細胞鋪滿培養瓶底進行傳代,以下所有試驗均取處于對數生長期的細胞。
1.2 材料和試劑 RPMI-1640培養基和0.25胰酶(0.25 Trypsin EDTA)購買于美國Gibco公司;胎牛血清(fetal bovine serum,FBS)和雙抗(Penicillin-Streptomycin Solution)購買于美國海克隆公司;PBS磷酸鹽緩沖液購買于中國碧云天生物公司。DRAIC-siRNA購買于上海吉瑪有限公司。氯仿、乙醇、異丙醇購買于重慶川東化工公司。Trizol裂解液、Premix ExTaqVersion2.0和SYBR Premix Ex Taq購買于日本Takara公司。PCR引物由上海Sangon Biotech公司合成。Transwell基質膠購買于美國BD公司;Transwell小室購買于美國康寧公司。
1.3 方法
1.3.1 細胞轉染 PBS緩沖液將細胞清洗干凈,胰酶消化細胞2 min,轉移至無菌15 mL離心管,離心并計數細胞,以每孔4×105個細胞接種于96孔板中,掌握過夜細胞融合率達70%左右,細胞分為DRAIC-siRNA組和NC組2組,無血清培養基將Lipofectamine 2000按3 μL/L進行稀釋,37 ℃孵育20 min,用無血清培養基分別將DRAIC-siRNA和NC按50 μM濃度稀釋,常溫下孵育5 min,最后與Lipofectamine 2000等體積分別混勻,分別加入2組細胞中,37 ℃孵箱繼續培養;12 h后,觀察轉染細胞狀態,并將無血清培養基更換為完全培養基,繼續培養,48h后,提取細胞RNA,驗證轉染效率。
1.3.2 實時熒光定量PCR分析 提取卵巢癌細胞總RNA,并逆轉錄合成相應cDNA。DRAIC上游引物序列為:5′-ATCTTTAGCTGGAGTTCGA-3′,下游引物序列為:5′-AAGGCTTGAGGGCGAGCGAT-3′;miR-181上游引物序列為:5′-ATCGTAGGTCGAGCGGAGC-3′,下游引物序列為:5′-GGGATGCGATTTGCGAGCT-3′。內參照GAPDH的上游引物的序列為5′-TTGGTATCGTGGAAGGACTCA-3′,下游引物序列為5′-TGTCATCATATTTGGCAG-GTT-3′。反應條件為:95 ℃預變性10 min,95 ℃15 s,60 ℃15 s,45個循環,獲取熒光信號溫度為60 ℃。普通cDNA實時熒光定量PCR檢測,采用2-△△Ct方法進行統計。
1.3.3 平板克隆細胞實驗 首先消化并計數細胞,以5 000/孔細胞數接種于96孔板中,每組3個復孔,37 ℃孵箱培養;待細胞貼壁,觀察細胞狀態,吸出培養基,避光條件下,分別加入不同慢病毒轉染的細胞中,37 ℃孵育24 h;檢測不同組細胞的克隆形成數目,并統計分析。
1.3.4 Transwell侵襲實驗 將5×104個細胞置于上室,并加入Matrigel基質膠。將具有10%FBS(20 ng/mL)的培養基置于下室中。在37 ℃下孵育24 h后,小心地去除上膜表面上的細胞。用95%乙醇固定20 min后,用0.5%結晶紫溶液染色10 min,自來水沖洗干凈過后,于倒置顯微鏡下計數。
1.3.5 熒光素酶報告基因測定 根據Clément等[7]研究方法,構建含DRAIC基因野生型3′UTR、突變型3′UTR片段,兩端均設計XbaⅠ酶切位點,插入熒光素酶報告基因質粒(pGL3-promoter,Promega)構建pGL3-3′UTR WT、pGL3-3′UTR MUT。取生長狀態良好的293T細胞,質粒轉染后48 h據Promega公司操作說明,裂解細胞,加入熒光素酶底物,檢測熒光素酶活性。
1.3.6 裸鼠體內成瘤實驗 取10只4~6周齡裸鼠,分為DRAIC-siRNA組和NC組,每組5只。將1×106濃度的轉染NC序列和DRAIC-siRNA的卵巢細胞分別注射到2組裸鼠的左側腋下。飼養8周后檢測裸鼠腫瘤異種移植物的體積和質量生長情況。異體移植物大小根據以下公式測量:體積=1/2(最短直徑)2×(最長直徑)。
1.3.7 Western Blot實驗 細胞分為NC組、DRAIC-siRNA組和miR-181-mimic組,從卵巢癌細胞SKOV3中提取總蛋白,將總蛋白的等分試樣行10% SDS-PAGE,然后電轉至PVDF膜上。分別使用抗JAK1、STAT2、p-JAK1、p-STAT2和GAPDH抗體進行孵育,4 ℃過夜,次日PBS洗滌后,加入Ⅱ抗,室溫孵育PBS充分洗滌后進行ECL顯影,拍照并統計相對表達量。

2 結果
2.1 LncRNA DRAIC和miR-181在卵巢癌細胞株中的表達情況 通過實時熒光定量PCR檢測lncRNA DRAIC在卵巢癌細胞株中的表達,結果顯示SKOV3中lncRNA DRAIC的表達(2.51±0.26)高于其他COC1(2.31±0.21),CAOV3(1.03±0.13)和A2780(1.93±0.12)細胞株(P<0.05),同時對miR-181在卵巢癌細胞株中的表達進行檢測,SKOV3中miR-181的表達(2.55±0.31)高于CAOV3(0.76±0.11)和A2780(1.62±0.13)細胞株(P<0.05),稍低于COC1細胞株(2.72±0.24)。圖1。后續實驗選取卵巢癌SKOV3細胞株作為研究對象。
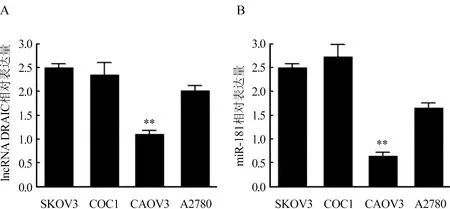
圖1 lncRNA DRAIC和miR-181在卵巢癌細胞株中的表達
2.2 雙熒光素酶實驗檢測lncRNA DRAIC和miR-181之間的關系 通過生物信息學檢測lncRNA DRAIC與miR-181之間的相互關系,結果顯示(圖2A),lncRNA DRAIC和miR-181在卵巢癌細胞中可能有相似結合的位點,表明lncRNA DRAIC與miR-181之間可能有調控關系。通過使用DRAIC-siRNA轉染卵巢癌細胞后,miR-181-WT的表達水平明顯降低[(1.02±0.11)vs(0.38±0.05),P<0.05],而miR-181-MUT的表達水平變化不大[(1.02±0.11)vs(0.96±0.08),P>0.05],圖2B。
2.3 LncRNA DRAIC對卵巢癌細胞增殖行為的影響 通過平板克隆細胞實驗檢測lncRNA DRAIC對卵巢癌細胞株SKOV3增殖能力的影響。首先使用特異的干擾RNA(DRAIC-siRNA)抑制卵巢癌細胞中lncRNA DRAIC的表達,在轉染DRAIC-siRNA 48 h和60 h后,SOKV3細胞的增殖能力明顯低于對照組(*P<0.05,**P<0.01),圖3。
2.4 LncRNA DRAIC對卵巢癌細胞侵襲行為的影響 Transwell實驗結果如圖4所示,抑制lncRNA DRAIC后,和NC組相比,DRAIC-siRNA組的侵襲細胞數目[(186.4±12.4) vs (73.6±8.6),P<0.05]顯著降低。
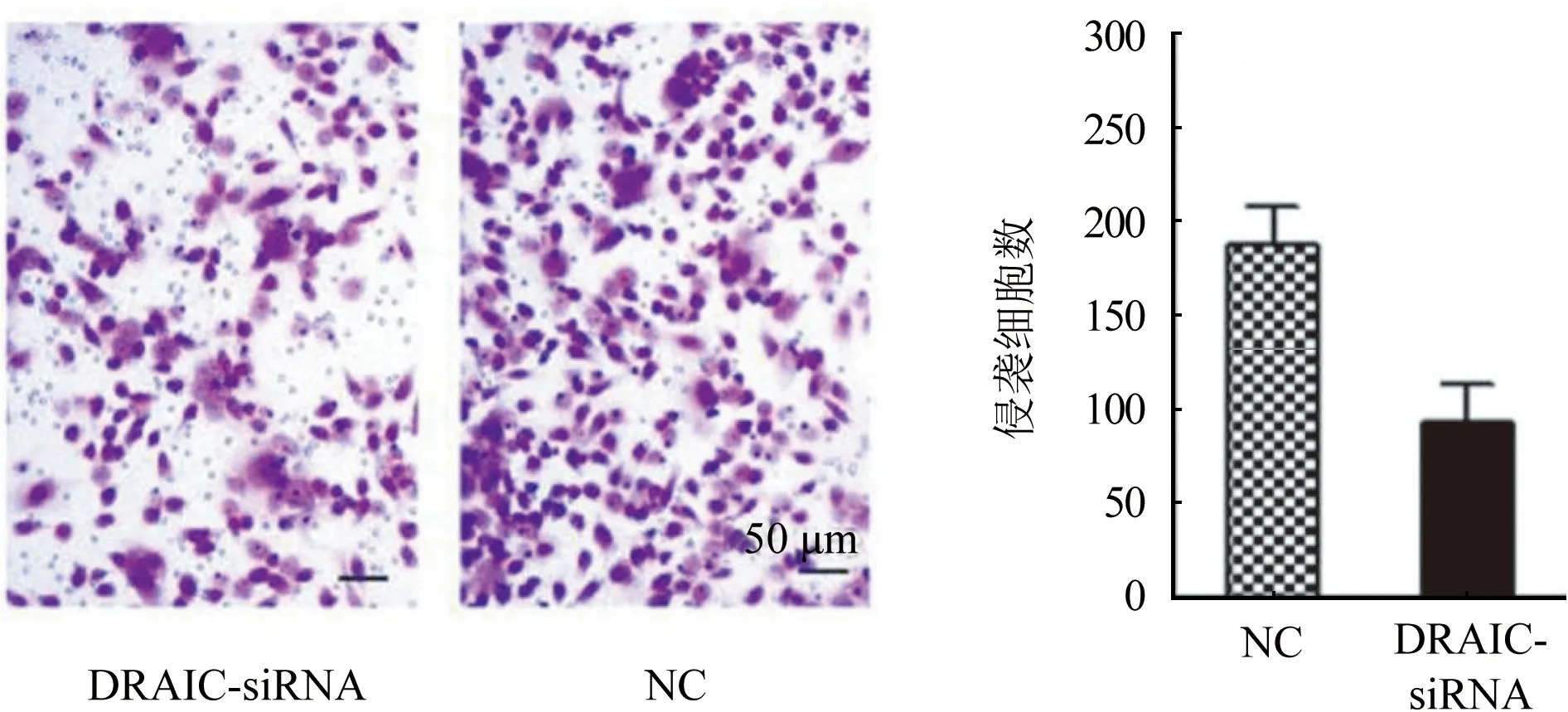
圖4 lncRNA DRAIC對卵巢癌細胞侵襲行為的影響
2.5 LncRNA DRAIC對卵巢癌細胞遷移行為的影響 劃痕愈合實驗結果如圖5所示,抑制lncRNA DRAIC后,和NC組相比,DRAIC-siRNA組的細胞遷移距離比例[(2.53±0.24) vs (1.21±0.09),P<0.05]顯著降低。
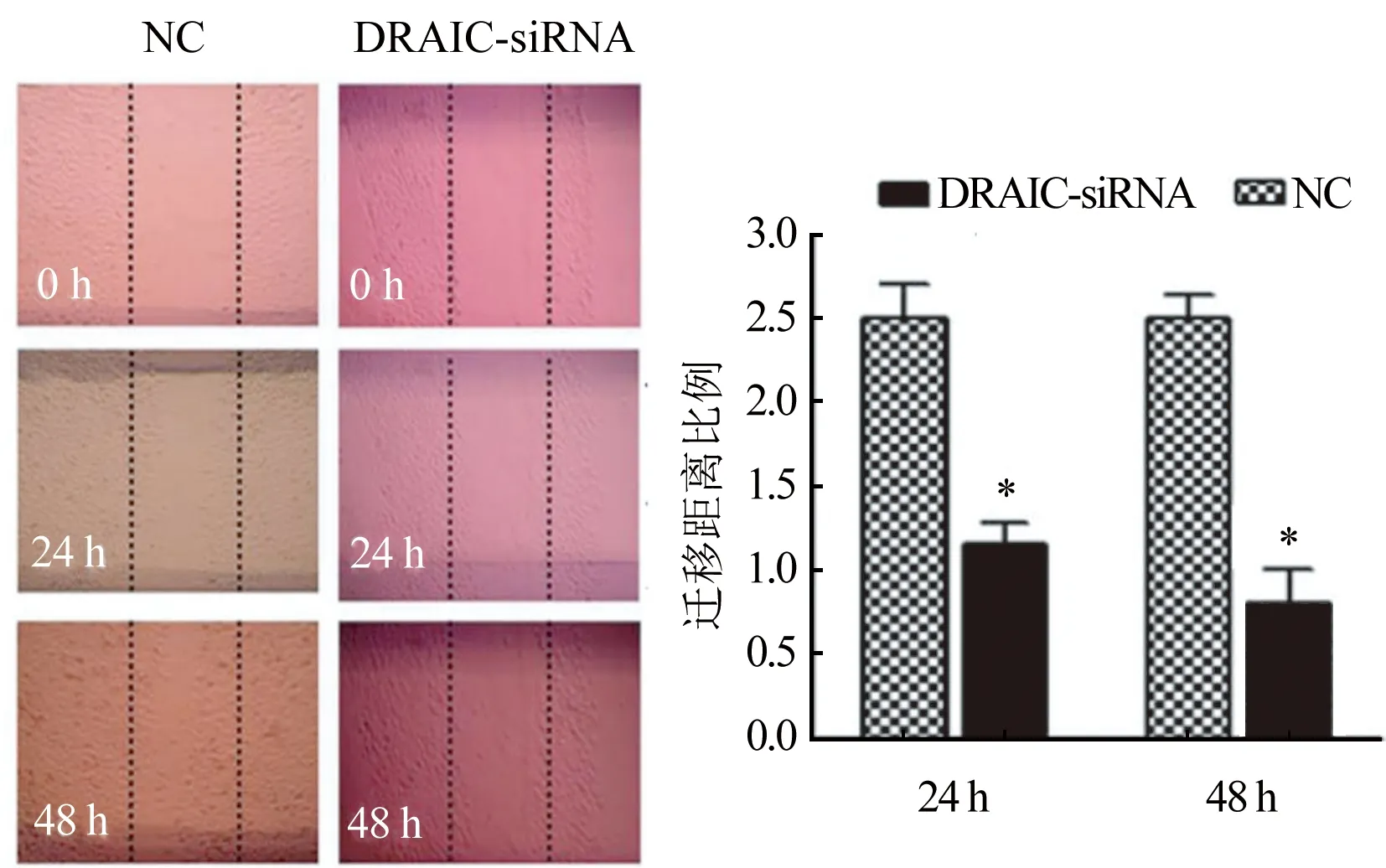
圖5 lncRNA DRAIC對卵巢癌細胞遷移行為的影響
2.6 LncRNA DRAIC對卵巢癌細胞裸鼠成瘤能力的影響 通過裸鼠異種移植腫瘤生長測定,DRAIC-siRNA組平均腫瘤體積明顯小于NC組[(1.135±0.09)cm3vs (2.13±0.13)cm3,P<0.05],而且DRAIC-siRNA組平均腫瘤重量也顯著低于對照組[(1.52±0.12)g vs (1.89±0.21)g,P<0.05]。圖6。
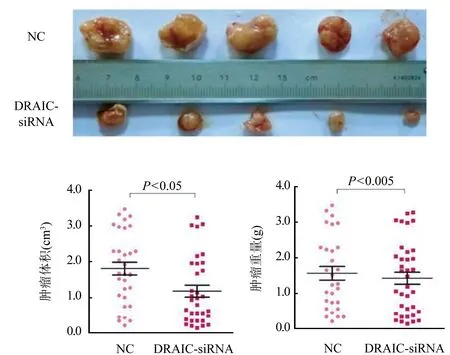
圖6 lncRNA DRAIC對卵巢癌細胞裸鼠成瘤能力的檢測
2.7 LncRNA對JAK1/STAT2信號通路激活的影響 Western Blot實驗結果顯示,與NC組相比,DRAIC-siRNA組的磷酸化p-JAK1和p-STAT2蛋白表達水平相應下調,而同時過表達miR-181-mimic后,磷酸化p-JAK1和p-STAT2蛋白表達水平相應的恢復,表明抑制DRAIC可以在一定程度上抑制JAK1/STAT2信號通路的激活,而同時增加miR-181的表達后對JAK1/STAT2的抑制效果相對緩解,同時非磷酸化的JAK1和STAT2蛋白表達未受明顯的影響。圖7。
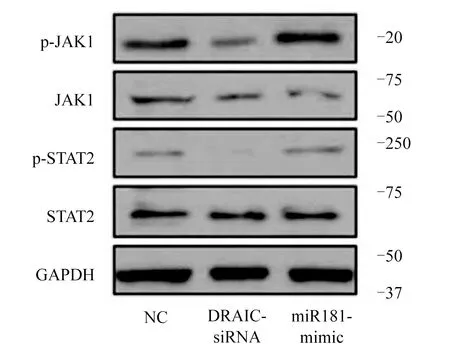
圖7 lncRNA對JAK1/STAT2信號通路激活的影響
3 討論
LncRNA DRAIC可以以不同細胞類型特異性方式進行動態變化。在成年小鼠組織中,lncRNA DRAIC在有限的細胞類型中表達,例如消化組織的上皮層。相比之下,lncRNA DRAIC在人類更廣泛的細胞類型中表達[8]。最近研究發現,lncRNA DRAIC的生物學作用主要與缺氧、病毒感染以及人胚胎干細胞分化有關[9]。據報道lncRNA DRAIC的水平在不同的癌癥類型中呈現動態調節。例如,血漿lncRNA DRAIC水平可作為非小細胞肺癌的探索指標[10];lncRNA DRAIC在結直腸癌中的表達上調,并與腫瘤的分化、浸潤、轉移和TNM分期有關[11];此外,高表達lncRNA DRAIC是結直腸癌預后不良的獨立預后指標[12];lncRNA DRAIC還參與了肝細胞癌,慢性淋巴細胞白血病和前列腺癌的疾病進展[13]。然而,它在卵巢腫瘤發生中的作用尚未明確。
本研究證實lncRNA DRAIC是人類卵巢癌細胞的致癌調節劑,針對卵巢癌lncRNA DRAIC的治療靶點研究具有潛在應用前景。本研究發現由lncRNA DRAIC介導的生物網絡對卵巢癌致癌作用貢獻很大,在這個復雜的網絡中,上游lncRNA DRAIC影響轉錄miR-181,進而激活JAK1/STAT2信號通路,JAK1/STAT2在卵巢癌細胞樣本中參與程度沒有明顯差異,而磷酸化的JAK1/STAT2的表達水平受lncRNA DRAIC和miR-181影響明顯。本研究結果表明miR-181受lncRNA DRAIC的調控表達活性降低,抑制miR-181有助于調控JAK1/STAT2靶標信號通路的激活,同時包括許多致癌基因的表達失調。本研究證實lncRNA DRAIC在卵巢癌細胞系中表達較高,這與之前相關研究的結果數據一致[14]。本研究結果表明lncRNA DRAIC可能在體外和體內控制卵巢癌細胞生長中發揮作用。更重要的是,在體內移植實驗中,lncRNA DRAIC水平的降低顯著地抑制了腫瘤生長。結果證實可能通過下調這種潛在的生物標志物來提供有效抑制腫瘤的治療方法,這將為卵巢癌患者的臨床治療和預后提供更有效途徑。
本研究存在一定的不足,只在lncRNA DRAIC序列中鑒定了一個miR-181結合位點,但當在不同條件下使用報告基因和內源基因作為檢測源時,它顯示出明顯的熒光活性水平不同,目前已經證明lncRNA DRAIC的作用位點和含有功能性miR-181的結合位點相近,但仍未證實通過生物信息學分析預測的更準確的結合位點,有必要進一步進行生物信息學分析以揭示可能具有強堿基對的miR-181的位點與目標基因的結合。
綜上所述,我們的研究表明卵巢癌細胞中特異性lncRNA DRAIC作為一種miRNA分子的調節劑,可以調控miR-181以干擾卵巢癌的基因的表達并進一步保持卵巢癌細胞的生物學功能,為卵巢癌的分子診斷和治療提供了新的研究途徑和方向。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
