二苯乙烯苷抗氧化和抗炎作用的機制研究
2020-04-09 14:33:20陳子卓徐宇航趙九洲朱敬樸鄭義鵬李文麗吳浩芝海春旭于衛華
癌變·畸變·突變 2020年2期
陳子卓,徐宇航,#,趙九洲,#,朱敬樸,鄭義鵬,李文麗,吳浩芝,海春旭,*,于衛華,*
(1.空軍軍醫大學基礎醫學院,陜西 西安710032;2.空軍軍醫大學軍事預防醫學院軍事毒理學與防化醫學教研室,陜西省自由基生物學與醫學重點實驗室,特殊作業環境危害評估與防治教育部重點實驗室.陜西 西安 710032)
巨噬細胞是機體內重要的免疫細胞,可通過細胞吞噬和釋放炎癥因子發揮免疫調節作用[1-2]。脂多糖(lipopolysaccharide,LPS)是革蘭氏陰性細菌細胞壁的主要成分,是一種內毒素和重要特異性抗原。巨噬細胞在LPS 的刺激下,會發生M1 型極化,并釋放TNFα和IL-6等多種促炎因子[3]。大量證據表明,炎癥反應與氧化應激關系密切,炎癥過程會生成大量的活性氧(reactive oxygen species,ROS),而氧化應激也可促進炎癥的發生發展,二者交互作用,形成惡性循環[4-5]。由于ROS與炎癥信號的相互依賴性,單純抗炎或單純抗氧化往往效果不佳,尋找兼具抗炎和抗氧化作用的藥物可能是解決問題的關鍵[6]。核因子E2 相關因子2(nuclear factor erythroid-2-related factor 2,Nrf2)可通過調控血紅素加氧酶1(heme oxygenase 1,HO-1)、錳超氧化物歧化酶(manganese superoxide dismutase,SOD2)、過氧化氫酶(catalase,CAT)和谷胱甘肽過氧化物酶1(glutathione peroxidase 1,GPX-1)等一系列抗氧化酶表達,從而清除細胞內過量ROS,減輕細胞的氧化損傷[7]。正常條件下Nrf2 存在于細胞質中且表達水平較低,但在氧化應激和抗氧化劑作用下Nrf2迅速激活,并轉位至細胞核發揮轉錄活性[8]。二苯乙烯苷(2,3,5,4’-tetrahydroxystilbene-2-O-β-D-glucoside,TSG)是一種多羥基酚類化合物,是中藥何首烏中提取的關鍵活性單體。研究表明,二苯乙烯苷具有較好的抗炎作用,在腸道炎癥、關節炎和脂肪性肝炎防治中具有良好效果[9-10]。研究表明,二苯乙烯苷具有抗氧化活性,可抑制ROS生成,并阻斷血管緊張素誘導的心臟成纖維細胞增殖[11]。此外,二苯乙烯苷可下調NADPH氧化酶介導的氧化應激,減輕小鼠缺血再灌注損傷[12-13]。查閱國內外文獻發現,二苯乙烯苷發揮抗炎和抗氧化作用的內在機制并不清楚,特別是二苯乙烯苷與Nrf2抗氧化調控的研究較少。因此,本研究借助LPS 誘導的RAW264.7 巨噬細胞促炎分化模型,重點研究二苯乙烯苷對Nrf2相關抗氧化酶系統調控作用,進而探討其抗炎抗作用機制,使人們正確認識二苯乙烯苷和中藥何首烏的藥理學作用,對于相關藥物研發意義重大。
1 材料與方法
1.1 材料
1.1.1 主要試劑小鼠巨噬細胞系(RAW264.7)由第四軍醫大學毒理學教研室細胞庫提供;TSG 由成都曼思特生物有限公司購買,純度98%;LPS 和地塞米松試劑購自Sigma公司;H2DCFH-DA和Mito-SOXTM熒光探針購自Invitrogen 公司;小鼠TNF-α和IL-6 酶聯免疫試劑盒購自武漢華美生物工程有限公司;寒鴉子酸(Brusatol)標準品自上海同田生物技術股份有限公司;Nrf2 抗體購自Santa Cruz 生物公司;細胞核蛋白提取試劑盒購自碧云天生物技術公司;DMEM 高糖培養液、胎牛血清、0.25%胰蛋白酶、PBS 購自上海吉諾醫藥生物技術有限公司。
1.1.2 儀器垂直流超凈臺購自ESCO,型號SCV-4A1;CO2培養箱購自Thermo 公司;全波段酶標儀購自infinite 公司,型號M200 PRO;激光共聚焦顯微鏡(Olympus fluo,FV10i);流式細胞儀購自BD 公司,型號Accuri C6,倒置顯微鏡購自Olympus 公司,型號IX7。
1.2 方法
1.2.1 RAW264.7細胞培養及實驗分組小鼠巨噬細胞系(RAW264.7)培養條件為:DMEM 高糖培養基(含10%胎牛血清),置于37 ℃、CO2體積分數為5%的孵箱中。1 μg/mL LPS 處理細胞12 h,建立炎癥模型[14]。藥物干預組分別給予30、60和120 μmol/L二苯乙烯苷預處理4 h。同時,100 nmol/L 的地塞米松(dexamethasone,Dex)作為陽性對照藥物預處理4 h。此外,為闡明Nrf2在二苯乙烯苷抗炎和抗氧化中關鍵作用,我們使用Nrf2 抑制劑寒鴉子酸(brusatol,50 μmol/L)與二苯乙烯苷(120 μmol/L)共同預處理4 h。本研究實驗分組如表1所示。
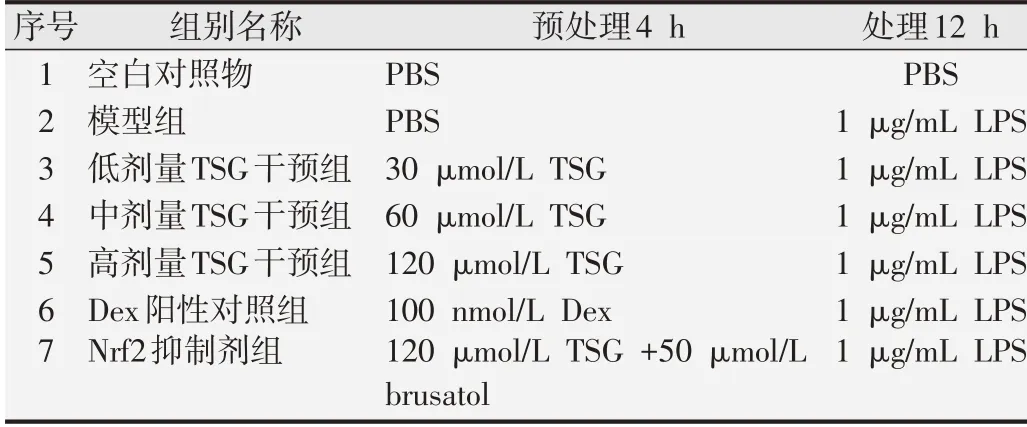
表1 實驗分組及處理
1.2.2 光鏡觀察巨噬細胞形態并計算活化比例對照組和LPS 處理后巨噬細胞,倒置顯微鏡觀察巨噬細胞形態。靜息態巨噬細胞呈圓形,體積較小;而LPS 刺激激活的巨噬細胞胞體明顯增大,且出現明顯觸角。因此,我們統計兩組中不同形態細胞的數目,每組統計50個細胞,并計算細胞活化比例。
1.2.3 細胞炎癥因子TNF-α和IL-6分泌水平檢測采用酶聯免疫法檢測細胞培養基中TNF-α和IL-6 含量。按照實驗分組1~7接種24孔板,每組重復3個樣,處理完成后,采用無菌管收集細胞上清,2 000 r/min離心10 min作為待測樣品。TNF-α和IL-6標準品按說明書稀釋液為0、1.56、3.12、6.25、12.5、25、50、100 pg/mL,按照試劑盒說明書加入待測樣品和標準品,37 ℃條件下孵育2 h,清洗后加入檢測工作液,37 ℃條件下孵育30 min,酶標儀450 nm 處檢測吸光度,根據標準曲線計算樣品中TNF-α和IL-6濃度。
1.2.4 細胞ROS水平檢測DCFH-DA是一種檢測細胞內ROS總體水平的熒光探針,自身被氧化后發出綠色熒光,因此熒光越強,表示ROS 水平越高。接種RAW264.7細胞于6孔板,按實驗分組1~5處理完成之后,加入終濃度為10 μmol/L的DCFH-DA,37℃條件下孵育30 min。PBS 清洗后,吹懸細胞,流式細胞儀檢測綠色熒光水平。因DCFH 熒光激發波長488 nm,發射波長525 nm,選擇FL1-A通道分析。
1.2.5 線粒體ROS 水平檢測MitoSOXTM是一種新型線粒體靶向性熒光探針,可選擇性檢測線粒體內的超氧化物水平。探針可透過活細胞膜,選擇性進入線粒體,繼而被超氧化物特異性的氧化,發出紅色熒光。接種RAW264.7細胞于6孔板,按實驗分組1~5處理完成之后,加入終濃度為5 μmol/L 的MitoSOXTM,37 ℃條件下孵育30 min。PBS 清洗后,吹懸細胞,流式細胞儀檢測紅色熒光水平。因DCFH 熒光激發波長510 nm,發射波長580 nm,選擇FL2-A通道分析。
1.2.6 免疫熒光法檢測Nrf2亞細胞定位通過免疫熒光方法檢測細胞內Nrf2亞細胞定位。激光共聚焦培養皿中RAW264.7 細胞生長至匯合度為70%時,給予120 μmol/L 二苯乙烯苷處理4 h。4%的多聚甲醛室溫固定細胞30 min,加入0.1% TritonX-100室溫條件下處理10 min。然后用5%的BSA 溶液封閉,37 ℃條件下1 h。培養皿中加入1∶100 稀釋的兔來源Nrf2 一抗,37 ℃濕盒中孵育過夜。PBS 洗細胞3 次,每次5 min。用1∶100 稀釋的FITC 標記羊抗兔二抗,37 ℃孵育1 h。最后加入終濃度為10 μg/mL 的DAPI,37 ℃染色30 min。PBS 洗細胞3 次,每次5 min。采用激光共聚焦顯微鏡檢測熒光強度,FITC(綠色)選擇Ex/Em:495 nm/530 nm;DAPI(藍 色)選 擇Ex/Em:340 nm/488 nm。
1.2.7 細胞核蛋白提取接種RAW264.7 細胞于6 孔板,分別給予0,30,60,120 μmol/L 二苯乙烯苷處理4 h。采用凱基細胞核提取試劑盒,按說明書進行操作:①收集2×106個細胞,加入1 mL 冰冷PBS 均漿,4 ℃條件下,500 g轉速離心3 min,棄上清;②每20 μL 體積沉淀,加入200 μL 預冷的Buffer A(Buffer A∶DTT∶PMSF∶蛋白酶抑制劑=1 000∶1∶5∶5),劇烈渦旋振蕩15 s,放置冰上10~15 min;③加入11 μL 冷Buffer B,最大轉速渦旋劇烈振蕩5 s,放置冰上1 min;④劇烈渦旋振蕩5 s 后,4 ℃離心,16 000 g 轉速離心5 min;⑤在沉淀(細胞核)中加100 μL 預冷的Buffer C (Buffer C∶DTT∶PMSF∶蛋白酶抑制劑=1 000∶1∶5∶5),劇烈渦旋振蕩15 s,放置冰上40 min;⑥4 ℃、16 000 g轉速離心10 min,即得核蛋白;⑦對提取的核蛋白進行蛋白定量(BCA法),分裝并保存于-80 ℃。
1.2.8 蛋白凝膠電泳測定細胞核中Nrf2蛋白表達采用BCA法測定提取的核蛋白樣品中蛋白含量,根據標準曲線計算蛋白濃度。蛋白凝膠電泳實驗檢測細胞核中Nrf2 蛋白表達,取定量處理后蛋白每組上樣20 μg,選用10%凝膠。PCNA 作為核蛋白內參,計算標準化處理后的Nrf2相對表達水平。
1.2.9 PCR 檢測細胞中抗氧化酶譜的mRNA 表達接種RAW264.7 細胞于6 孔板,給予120 μmol/L 二苯乙烯苷處理4 h。使用Trizol法提取細胞中總RNA,采用核酸定量儀檢測RNA 濃度。采用TIANGEN 公司反轉錄試劑盒,按照說明書操作獲得cDNA。小鼠GAPDH、 HO-1、 SOD2、 SOD1、 CAT、 NQO-1 和GPX-1對應的RNA引物由北京奧科鼎盛生物合成,序列如表2 所示。以cDNA 為模板,使用Tiangen 公司熒光定量PCR 試劑盒進行擴增,以GAPDH 為內參標準化,計算各組抗氧化酶譜的mRNA相對表達量。

表2 引物序列
1.3 統計學方法
實驗數據以均數±標準差表示,采用方差分析和LSD t 檢驗進行各組間數據比較,以α=0.05 為檢驗水準,數據統計分析采用SPSS 13.0軟件。
2 結 果
2.1 TSG抑制LPS誘導的RAW264.7細胞激活
二苯乙烯苷和脂多糖處理后RAW264.7 細胞形態學變化見圖1A。RAW264.7 細胞是小鼠巨噬細胞系,正常狀態下貼壁生長,形態為圓形或橢圓形,體積較小。在暴露于內毒素或不利生長環條件下,細胞會被激活并伴有形態和功能改變,主要表現為胞體增大,伸出大量偽足,形態不規則。我們統計了激活態細胞的比例,見圖1B。1 μg/mL LPS 的處理12 h 后,RAW264.7 細胞激活比例達到80%,而120 μmol/L TSG 干預組僅有約20%的細胞發生形態改變。說明二苯乙烯苷可阻斷LPS介導的RAW264.7細胞激活。
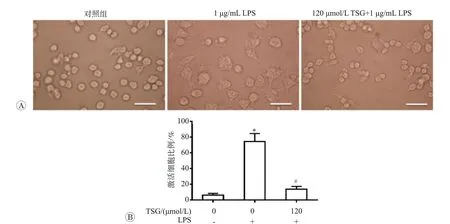
圖1 二苯乙烯苷和脂多糖干預對RAW264.7細胞激活狀態的影響
2.2 TSG 阻斷LPS 誘導的RAW264.7 細胞炎癥因子釋放
巨噬細胞在內毒素等不利因素刺激下,可以釋放大量促炎因子如TNF-α、IL-8、IL-6、IL-1等。我們采用ELISA法檢測培養基中炎癥因子IL-6和TNF-α水平,如果如圖2 所示。1 μg/mL 的LPS 處理后IL-6 和TNF-α分別增加了17 倍和16 倍(P<0.05),而60 和120 μmol/L 的TSG 干預有效抑制了LPS 介導的炎癥因子釋放(P<0.05)。同時,我們使用100 nmol/L的Dex作為抗炎的陽性對照藥物。結果發現120 μmol/L的TSG處理后抗炎效果優于地塞米松干預組(P<0.05)。上述結果表明,TSG可以有效抑制LPS誘導的炎癥反應。
2.3 TSG抑制LPS誘導的RAW264.7細胞ROS升高
炎癥與氧化應激密不可分炎癥過程會釋放大量的ROS殺傷入侵的細菌病毒。同時ROS的釋放又可以進一步誘導炎癥因子釋放,加重炎癥反應。我們采用DCFH-DA 和MitoSOXTM染色檢測細胞和線粒體水平ROS 水平,結果見圖3。1 μg/mL LPS 處理后細胞內DCFH-DA 和MitoSOXTM熒光強度均明顯增強(P<0.05),表明細胞處于氧化應激狀態。給予30、60 和120 μmol/L TSG干預后細胞和線粒體ROS均明顯下調(P<0.05)。表明二苯乙烯苷具有抗氧化作用,可有效抑制LPS誘導的氧化應激。
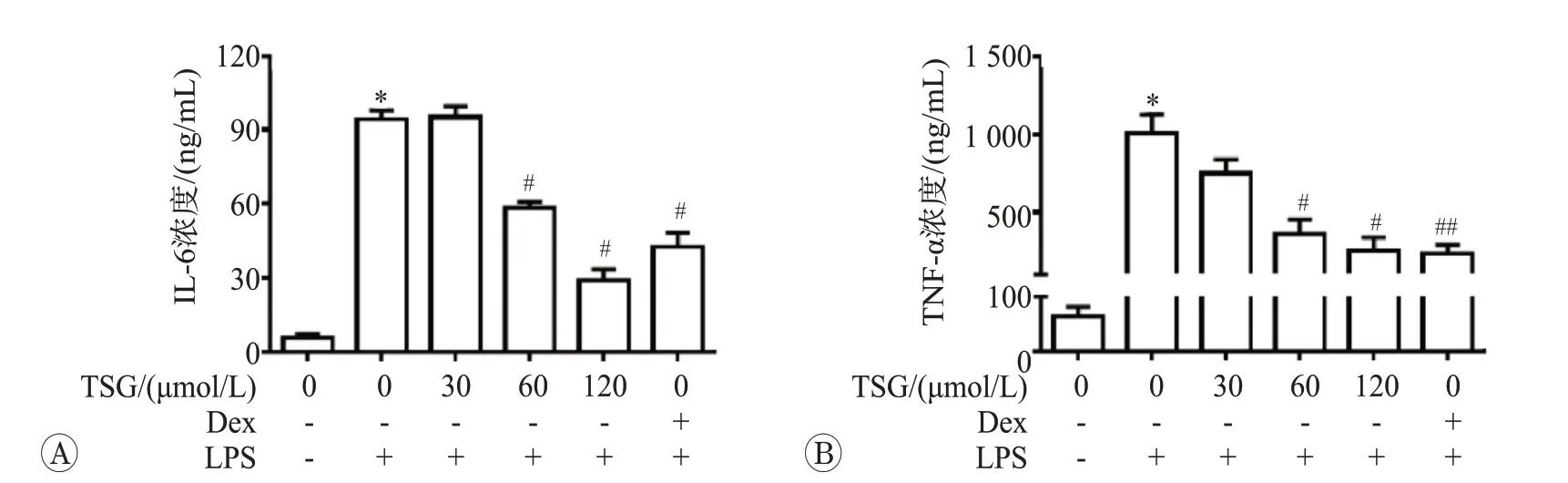
圖2 二苯乙烯苷對脂多糖誘導RAW264.7細胞釋放IL-6和TNF-α水平影響
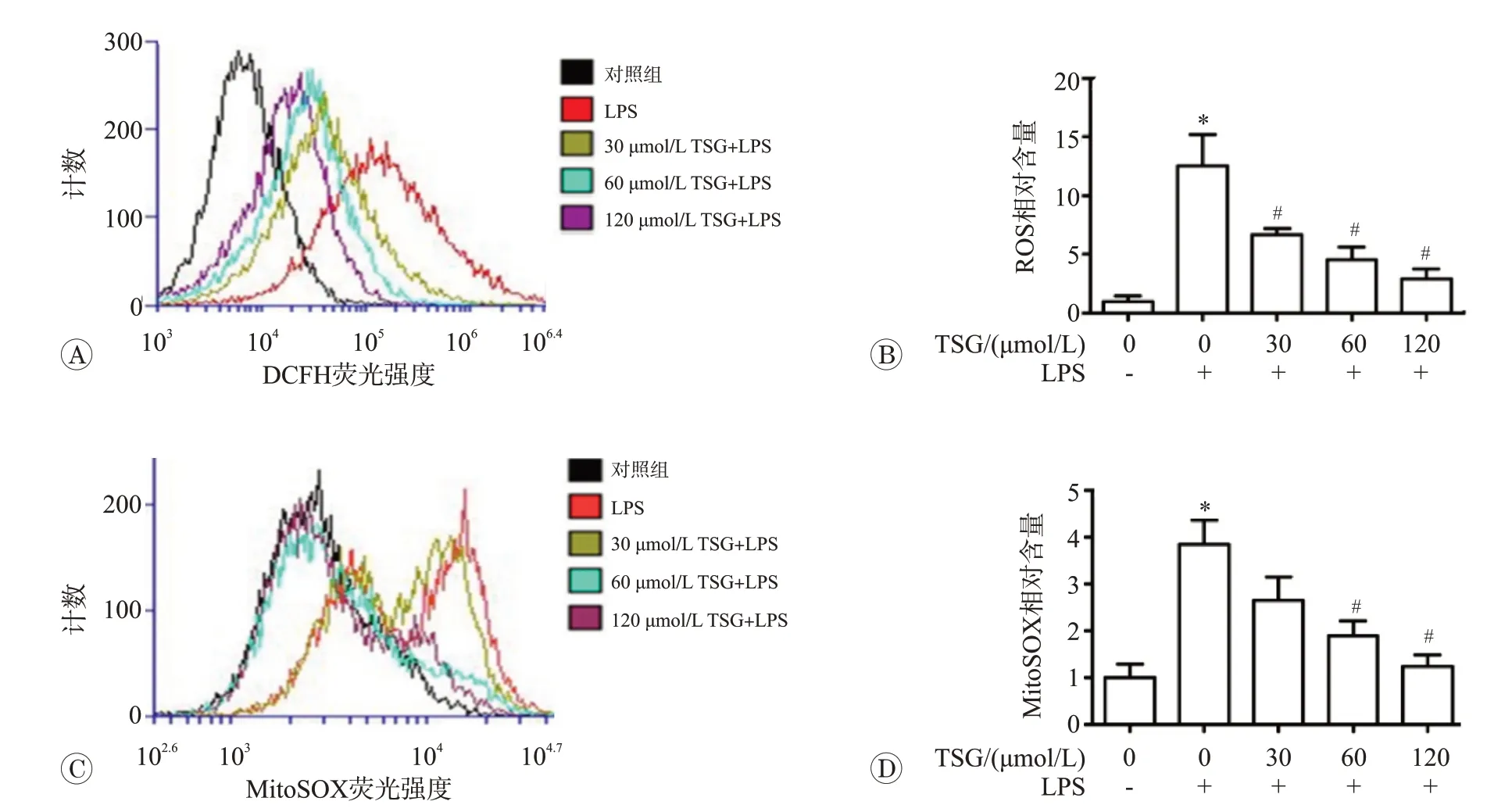
圖3 二苯乙烯苷對脂多糖誘導RAW264.7細胞ROS生成的影響
2.4 TSG促進Nrf2為核心的抗氧化酶系統表達
二苯乙烯苷對RAW264.7 細胞Nrf2 及其下游抗氧化酶系統影響見圖4。Nrf2 是重要的氧化還原調控因子,在正常情況下表達量較低且存在于細胞質中,在ROS和抗氧化劑刺激下便可迅速轉位至細胞核,通過與抗氧化原件(ARE)結合,激活下游抗氧化酶系統。免疫熒光結果表明,給予120 μmol/L TSG處理后Nrf2綠色熒光增強,且出現大量核內聚集現象(圖4A)。為進一步證實TSG 是否導致了Nrf2 核轉位,我們使用30、60和120 μmol/L TSG處理RAW264.7細胞,提取細胞核蛋白。如圖4B 所示,Western blot 結果表明,與對照組相比,TSG 處理組細胞核中Nrf2 水平明顯升高(P<0.05)。檢測Nrf2 下游抗氧化酶表達發現,120 μmol/L TSG明顯促進抗氧化酶HO-1、SOD2和CAT表達(P<0.05)。特別指出的是HO-1 表達增強尤為明顯,是對照組的26 倍(圖4C)。這些結果證明,TSG 激活了Nrf2為核心的抗氧化酶系統。
2.5 TSG的抗炎效應依賴于調控Nrf2抗氧化系統
為進一步驗證Nrf2 在TSG 介導的抗炎效應中作用,我們使用Nrf2 特異性抑制劑寒鴉子酸(Brusatol)干預,并檢測細胞培養基中IL-6和TNF-α的水平,結果見圖5。結果表明,120 μmol/L TSG 明顯抑制了LPS介導的TNF-α和IL-6釋放,而給予50 μmol/L Brusatol處理,可顯著阻斷TSG 發揮抗炎作用。這些結果證實,TSG 介導的抗炎反應依賴于Nrf2 抗氧化系統的激活。
3 討 論
炎癥反應是機體重要的自我防御機制,當機體遭遇病原體侵襲或創傷時,會招募免疫細胞到感染位點并釋放炎癥因子,清除病原菌。但是炎癥失控也會引起一系列的病理反應,如嚴重的感染、敗血癥、阿爾茨海默氏癥、動脈粥樣硬化、2 型糖尿病和腫瘤等。巨噬細胞是機體內重要的炎癥效應細胞,可以通過吞噬作用和釋放炎癥因子發揮免疫效應。RAW264.7 細胞是小鼠來源巨噬細胞系,正常情況下為圓形,體積較小。脂多糖刺激后胞體明顯增大,形成大量偽足,形態不規則[15]。我們的研究也證實,脂多糖刺激后RAW264.7 細胞發生顯著的形態學改變,激活形態的細胞比率增加。TNF-α和IL-6 既是重要的炎性介質,也可激活其他細胞因子的合成釋放。LPS 刺激后培養基中TNF-α和IL-6水平明顯升高,這表明LPS促進了巨噬細胞的活化和炎癥反應過程。
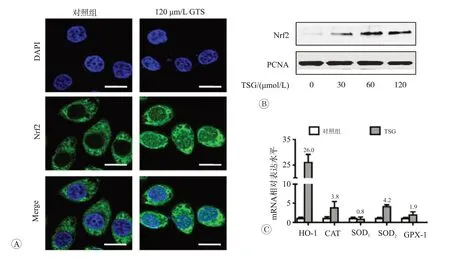
圖4 二苯乙烯苷對RAW264.7細胞Nrf2及其下游抗氧化酶系統影響
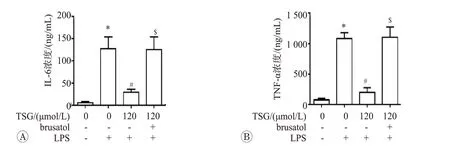
圖5 二苯乙烯苷調控的抗炎效應依賴于Nrf2通路的激活
二苯乙烯苷是一種多羥基酚類化合物,是中藥何首烏的主要活性成分。大量研究表明,二苯乙烯苷具有廣泛的生物學作用,具有神經保護、抗衰老、抗血脂、抗動脈粥樣硬化以及抗腫瘤的功能[16-18]。炎癥在上述疾病的發生和發展過程中均發揮關鍵作用,但二苯乙烯苷介導抗炎作用及相關機制并不清楚。我們前期研究發現,二苯乙烯苷可抑制亞硝胺誘導的肝毒性,減輕肝臟炎癥和DNA損傷[19]。而在本研究中,二苯乙烯苷預處理可抑制LPS 誘導的小鼠巨噬細胞活化,降低培養基中TNF-α和IL-6水平,且在一定劑量范圍內呈現劑量效應關系。與地塞米松陽性對照藥物相比,二苯乙烯苷表現出了良好的抗炎效果,有望作為一種抗炎藥物使用。
炎癥和氧化應激密不可分,炎癥可以導致ROS的大量生成,ROS也會促進NFκB和COX2等炎癥轉錄因子的激活,二者形成級聯反應[20]。因此,氧化應激是巨噬細胞激活和炎癥反應中的關鍵事件,而抑制ROS生成或者給予抗氧化劑可阻斷LPS 等誘導的巨噬細胞炎癥因子增多[21-22]。本研究中我們也發現,LPS刺激后RAW264.7 細胞和線粒體水平ROS 生成顯著增多,而二苯乙烯苷干預后細胞中的ROS明顯降低,提示二苯乙烯苷的抗炎效應可能與其清除細胞內ROS 能力相關。Nrf2 為核心的抗氧化系統,可通過激活HO-1、SOD2、SOD1、CAT、NQO-1 和GPX-1。等抗氧化酶譜表達,有效保護細胞應對多種不利性刺激造成的氧化損傷[23]。Nrf2 缺失可加劇血管緊張素II 誘導的炎癥反應和心肌損傷[24]。Nrf2 信號通路激活在葉黃素、烏司他丁和天青素等介導的抗炎反應中發揮關鍵作用[25-26]。我們發現二苯乙烯苷可誘導Nrf2 蛋白水平的上調和核轉位,并激活了下游HO-1、SOD2和CAT表達。其中,HO-1 升高最為顯著,約為對照組的26倍。研究表明,HO-1 及其代謝產物膽紅素和一氧化碳具有顯著的抗氧化和抗炎作用,過表達HO-1 可抑制TNF-α誘導的氣道炎癥[27]。安石榴苷也可通過激活Nrf2/HO-1 表達抑制LPS 誘導的巨噬細胞促炎因子升高[28]。為進一步證明二苯乙烯苷是通過激活Nrf2 通路發揮抗炎效應,我們使用Nrf2 活性抑制劑Brusatol 與TSG共處理,發現Brusatol可逆轉二苯乙烯苷介導的抗炎作用。這表明二苯乙烯苷可顯著增強Nrf2及其下游抗氧化酶活性,可清除LPS 誘導巨噬細胞生成的ROS,降低炎癥因子表達。
氧化應激是炎癥反應的重要組成部分,因此在治療全身炎癥反應綜合征及慢性炎癥性疾病時不可忽視抗氧化應激的治療[6,22]。聯合使用抗氧化類和抗炎類藥物是治療炎性疾病的關鍵,這種觀點受到越來越多基礎研究和臨床應用的支持。作為何首烏的主要活性成分,二苯乙烯苷刺激通過Nrf2表達及核轉位,激活下游HO-1、SOD2和CAT等抗氧化酶表達,最終發揮了較好的抗氧化和抗炎作用。本研究明確了二苯乙烯苷的抗炎和抗氧化分子機制,指出了炎癥相關疾病的預防和治療新的思路,也為何首烏相關中藥開發提供了理論支撐。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
海峽科技與產業(2016年3期)2016-05-17 04:32:12
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
