春季低溫對小麥穎果發育的影響
2020-04-01 07:23:42余徐潤顧清欽倪愛淳夏敏潔
麥類作物學報 2020年7期
余徐潤,郝 朵,顧清欽,倪愛淳,夏敏潔,臧 勇,熊 飛
(江蘇省作物遺傳生理國家重點實驗室培育點/糧食作物現代產業技術協同創新中心/揚州大學教育部農業與農場品安全國際合作聯合實驗室,江蘇揚州 225009)
初春氣溫回升較快,而在春季后期氣溫偏低的天氣現象稱為倒春寒,此時,小麥的穗正處于對低溫極其敏感的雌雄蕊分化期至藥隔期之間。低溫會導致小麥營養生長、生殖生長均受到抑制,在一定程度上使小麥減產。研究表明,小麥籽粒的品質與產量是由基因型和環境因素共同決定的[1],但小麥籽粒的品質受到環境因素的影響更大[2]。小麥生長發育過程中,低溫使小麥的穗生長量減少、千粒重下降[3],同時還會減緩小麥的發芽速度,阻礙幼苗的生長[4]。晉鵬宇等[5]研究表明,春季低溫陰雨,會導致小麥在穗分化中后期可孕小花數及籽粒結實數減少,最終造成產量下降。而這些產量下降的原因一方面可能在于碳水化合物的代謝過程受到了低溫的影響[6];另一方面,在生理水平上,低溫脅迫降低了能量利用的能力,從而抑制了光合作用的進程[7]。而小麥為適應低溫環境的影響,其內源激素含量也會發生相應的變化[8],在低溫脅迫下,氮同化能力也會發生一定的改變,從而使籽粒中蛋白質的積累發生變化[9]。
前人研究都集中在探究低溫脅迫對小麥籽粒品質、產量以及物質能量代謝等方面的影響,有關春季低溫條件下小麥籽粒形態結構發育的研究較少。本研究以揚麥15為材料,在雌雄蕊分化期至藥隔期之間設置-3 ℃低溫處理,從顯微和超微結構水平觀察低溫處理下小麥穎果果皮消亡、胚乳儲藏物質積累以及養分運輸組織微觀結構的變化,以期解析小麥穎果發育過程中的結構變化對春季低溫的響應機制,為培育抗寒小麥品種提供理論參考。
1 材料與方法
1.1 試驗材料
本研究所選用的小麥品種為揚麥15,由江蘇里下河地區農業科學研究所提供,于2018-2019年種植于揚州大學江蘇省作物栽培重點實驗室試驗棚內。使用盆栽方式種植,每盆播種20粒種子,通過體式顯微鏡剝離幼穗,確定小麥幼穗處于雌雄蕊分化期至藥隔期時,于揚州市農科所低溫室進行低溫處理。濕度設定為75%RH,CO2設置為500 ppm。對照組溫度為0 ℃,低溫處理組白天0 ℃處理14 h,夜間-3 ℃處理10 h,處理 3 d后搬回試驗棚內繼續進行正常溫度生長,后續管理均一致。低溫處理組與對照組均選取3盆盆栽作為試驗材料。在小麥開花時,用記號標注穗中部小穗基部的第二朵小花并掛記號牌準確記錄開花時間。
1.2 穎果組織結構的半薄切片觀察
將不同發育時期的穎果中部進行橫切,厚約2 mm,參照熊 飛等[10]的方法制作樹脂樣品塊,采用超薄切片機將樣品切為1 μm厚的薄片,用 0.5%甲基紫以及甲苯胺藍溶液對切片進行染色,然后在光學顯微鏡下觀察并拍照。每組隨機挑選3個穎果作為重復,每個穎果均來自不同麥穗。
1.3 細胞數目和果皮厚度觀測
使用Image-Pro Plus圖像分析軟件對拍攝的顯微圖像進行處理,將標尺設定后利用軟件中的長度和距離快捷工具測量穎果長度、寬度等。利用軟件中計數功能,對果皮細胞層數等進行統計。每組樣品均選取3粒穎果作為重復,并將每粒穎果拍攝3張顯微圖片,后續進行數據統計。
1.4 細胞及細胞內含物面積統計
參考余徐潤等[11]的方法,用Image-Pro Plus和Photoshop軟件對腹部胚乳中淀粉體和蛋白體的相對面積、養分運輸途徑中韌皮部及珠心突起傳遞細胞面積。每組樣品均選取3粒穎果作為重復,并將每粒穎果拍攝3張顯微圖片,后續進行數據統計。
1.5 數據統計與分析
使用Excel處理數據,采用SPSS 19.0進行t檢驗。圖表制作采用Photoshop CS6圖像處理軟件和Origin 8.5函數繪圖軟件。
2 結果與分析
2.1 春季低溫對穎果外觀形態的影響
穎果發育早期呈乳白色,發育至花后15 d左右時,其縱向生長基本固定,其后進行橫向生長,與對照組相比,低溫處理組穎果果皮顏色更綠(圖1A)。花后20 d,穎果形態基本成型,果皮顏色從綠色向黃色轉變。花后30 d,與對照組相比,低溫處理組果皮顏色更深。通過對穎果的長度和寬度進行統計,發現花后5~20 d,低溫處理組的穎果長度均大于對照組,而在花后25~30 d時,對照組的穎果長度均大于低溫處理組(圖1B)。而在發育過程中穎果的寬度,對照組均寬于低溫處理組(圖1C)。

A圖下方的數字表示花后天數(d);*表示對照和處理組間差異顯著(P<0.05);CK為對照組;LTT為低溫處理。Numbers in figure A indicate the days(d) after flowering. * indicates significant difference between control and treatment groups (P<0.05). CK is the control; LTT is the low temperature treatment.圖1 低溫處理下穎果形態發育過程(A)及其長度(B)和寬度(C)的變化Fig.1 Changes of caryopsis morphological development(A) and its length(B) and width(C) under low temperature treatment
以上結果表明,在穎果發育早期,低溫促進了穎果的長度生長、抑制了寬度生長;與對照相比,低溫處理組小麥穎果發育時期縮短。
2.2 春季低溫對穎果果皮發育及消亡的影響
花后5 d,中果皮細胞開始凋亡,大量游離淀粉粒充斥在中果皮和內果皮的空隙間(圖2A、D、A1、D1),對照組的果皮細胞層數多于低溫處理組。花后10 d,可以觀察到低溫處理組小麥穎果果皮細胞進一步降解,其果皮細胞層數少于對照組,且出現凋亡空腔(圖2B、E)。花后15 d,中果皮細胞基本降解完全,此后果皮細胞的層數不發生變化,兩者之間無明顯差異(圖2C、F)。花后20~30 d之間,果皮中淀粉粒完全降解,果皮細胞層數保持不變(圖2G~L)。

A、B、C、G、H、I 分別為正常溫度下小麥花后5、10、15、20、25、30 d的穎果果皮顯微結構;D、E、F、J、K、L 分別為低溫處理下小麥花后5、 10、15、20、25、30 d的穎果果皮顯微結構;A1、B1、D1、E1均為相應時期完整結構圖;SG:淀粉體;Ex:外果皮; Me:中果皮。A,B,C,G,H and I are the pericarp microstructures of wheat caryopsis at normal temperature at 5,10,15,20,25 and 30 d after flowering; D,E,F,J,K and L are the microstructure of the pericarp in wheat caryopsis at 5,10,15,20,25 and 30 d under low temperature treatment; A1,B1,D1 and E1 are complete structural diagrams of the corresponding periods; SG:Starch granule; Ex:Exocarp ; Me:Mesocarp.圖2 低溫處理對小麥果皮發育的影響Fig.2 Effect of low temperature treatment on the development of wheat pericarp
上述結果表明,在穎果發育5~10 d時,對照組果皮細胞層數均多于低溫處理組,且果皮中含有較多淀粉,說明對照組發育較遲緩,且果皮降解時間晚。但隨著生長發育的進行,果皮淀粉粒發生降解,發育后期,中果皮細胞逐步凋亡。
2.3 春季低溫對穎果胚乳發育的影響
從圖4可以看出,花后5 d,腹部胚乳細胞剛開始分化(圖3A、D,圖4A、D),只有少量的物質積累。花后10 d,細胞開始生長分化,此時,腹部胚乳細胞中出現淀粉體,且大多聚集在細胞壁周圍,同時細胞中也開始積累蛋白體。通過顯微和超微結構觀察及統計分析(圖3B、E,圖4B、E,表1),發現該階段中低溫處理組淀粉體含量高于對照組,但蛋白體含量少于對照組。花后15 d,蛋白體的積累量持續增加,淀粉體的數量也不斷增多,此時對照組蛋白體的含量多于低溫處理組,而淀粉體數量低溫處理組較多(圖3C、F,圖4C、F,表1)。花后20 d,蛋白體之間相互靠近,使得蛋白體體積增大,且細胞中聚集了一些小淀粉體(圖3G、J,圖4G、J)。花后25 d,蛋白體相互聚集,此時,對照組蛋白體數量持續多于低溫處理組,淀粉體含量仍然較少(圖3H、K,圖4H、K,表1),但對照組腹部充實度高于低溫處理組。花后30 d,大量淀粉體和蛋白體充實于細胞內,且在對照組中較多蛋白體聚集在一起,低溫處理組蛋白體數量低于對照組,且胚乳充實度仍然小于對照組(圖3I、L,表1)。
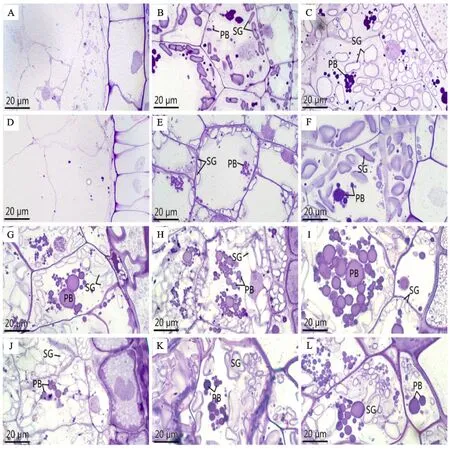
A、B、C、G、H、I 分別為正常溫度下小麥穎果花后5、10、15、20、25、30 d的腹部胚乳細胞;D、E、F、J、K、L 分別為低溫處理下小麥穎果花后5、10、15、20、25、30 d的腹部胚乳細胞;SG:淀粉體;PB:蛋白體。A,B,C,G,H and I are abdominal endosperm cells at 5,10,15,20,25 and 30 d after flowering of wheat caryopsis at normal temperature; D,E,F,J,K and L are the abdominal endosperm cells at 5,10,15,20,25 and 30 d after anthesis of wheat caryopsis under low temperature treatment; SG:Starch granule; PB:Protein body.圖3 低溫處理對小麥穎果腹部胚乳細胞發育的影響Fig.3 Effect of low temperature treatment on the development of abdominal endosperm cells in wheat caryopsis

A、B、C、G、H、I 分別為正常溫度下小麥穎果花后5、10、15、20、25、30 d的腹部胚乳細胞(×3 000);D、E、F、J、K、L 分別為低溫處理下小麥穎果花后5、10、15、20、25、30 d的腹部胚乳細胞(×3 000);SG:淀粉體。A,B,C,G,H,I are abdominal endosperm cells(× 3 000) at 5,10,15,20,25 and 30 d after flowering of wheat caryopsis at normal temperature; D,E,F,J,K and L are abdominal endosperm cells(× 3 000) at 5,10,15,20,25 and 30 d after flowering of wheat caryopsis under low temperature treatment,respectively. SG:Starch granule.圖4 低溫處理下小麥穎果腹部胚乳細胞的掃描電鏡圖片Fig.4 Scanning electron microscopy images of abdominal endosperm cells in wheat caryopsis under low temperature treatment
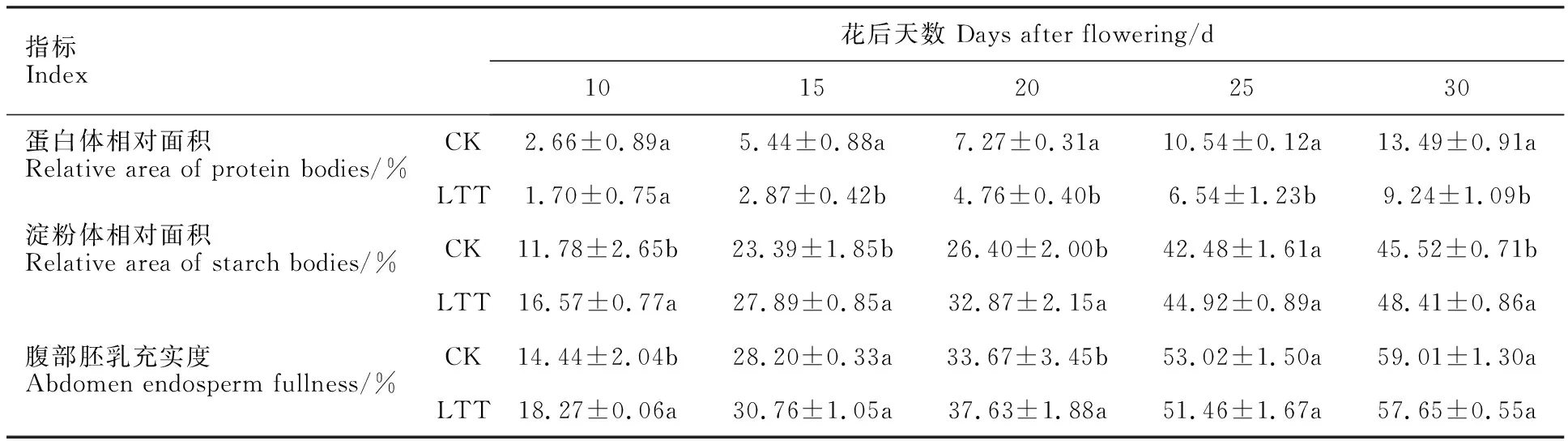
表1 低溫處理對小麥穎果腹部胚乳蛋白體和淀粉體面積以及充實度的影響Table 1 Effect of low temperature treatment on the area of protein body and starch granule and the plumpness of abdominal endosperm in wheat caryopsis
以上結果表明,穎果發育早期低溫處理組淀粉體發育較快,腹部胚乳充實度顯著高于對照組,因此,低溫處理在穎果發育的早期促進了腹部胚乳的分化,加速了胚乳的發育。但隨著發育進行,腹部胚乳發育漸緩,蛋白體含量以及腹部胚乳充實度均低于對照組。
2.4 低溫處理對穎果養分運輸組織發育的影響
通常情況下,小麥韌皮部是用來運輸光合產物等有機物質,在小麥維管束的發育過程中,主要依賴于韌皮部的發育,從而進行同化物的運輸。花后5 d,韌皮部中的薄壁細胞出現,具有明顯的細胞核(圖5A、D),此時低溫處理組的韌皮部面積大于對照組(表2)。花后10 d,維管薄壁中細胞核數量均減少(圖5B、E),但對照組韌皮部面積小于低溫處理組。花后15 d,篩管細胞數均明顯增加,維管薄壁細胞中細胞核數量均有所減少(圖5C、F)。花后20 d,維管薄壁細胞變得不規則,且開始凋亡(圖5G、J)。花后25~30 d,韌皮部面積持續增加且低溫處理組面積較大(圖5H、I、K、L,表2)。
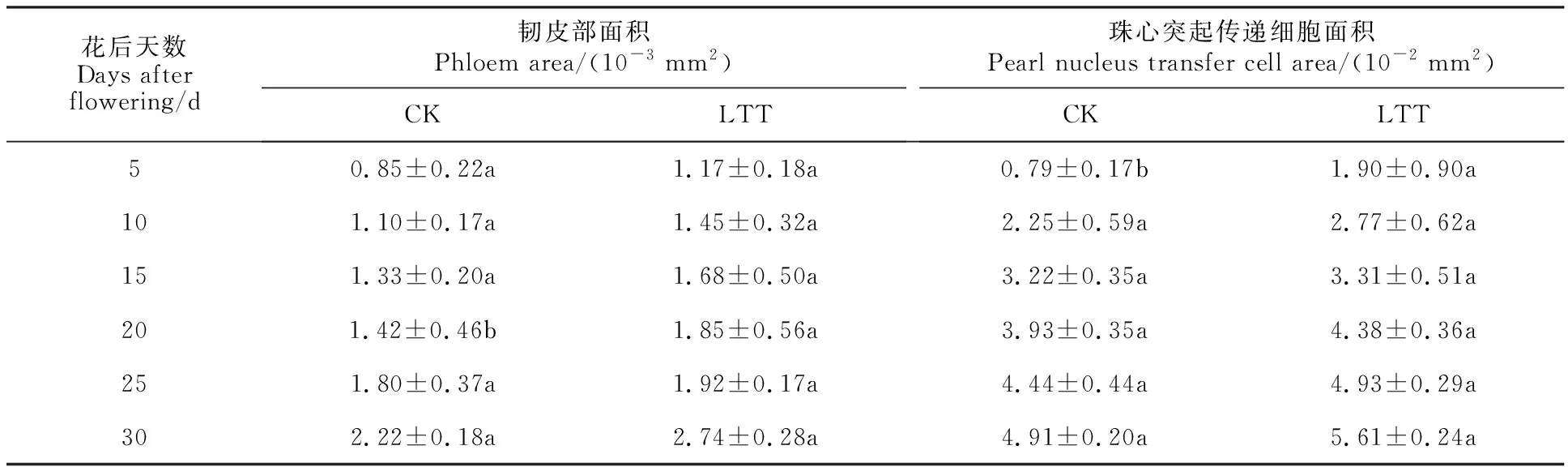
表2 低溫處理對穎果維管束中韌皮部面積及珠心突起傳遞細胞面積的影響Table 2 Effect of low temperature treatment on phloem area and the area of nucellar projection transfer cells in caryopsis vascular bundle

A、B、C、G、H、I 分別為正常溫度下小麥穎果花后5、10、15、20、25、30 d的韌皮部細胞;D、E、F、J、K、L 分別為低溫處理下小麥穎果花后5、10、15、20、25、30 d的韌皮部細胞;Nu:細胞核; ST:篩管;SG:淀粉體。A,B,C,G,H and I are phloem cells at 5,10,15,20,25 and 30 d after flowering of wheat caryopsis under normal temperature; D,E,F,J,K and L are the phloem cells at 5,10,15,20,25 and 30 d after flowering in wheat caryopsis under low temperature treatment; Nu:Nucleus; ST:Sieve tube; SG:Starch granule.圖5 低溫處理對穎果韌皮部發育的影響Fig.5 Effect of low temperature treatment on the development of phloem in caryopsis
以上結果表明,低溫處理使韌皮部面積增大,物質的運輸效率更高,更有利于養分的輸送。
花后5 d,小麥穎果具有較大的珠心突起細胞,細胞內有大的細胞核,且最外層的細胞出現加厚(圖6A、D),低溫處理組珠心突起細胞厚度大于對照組。花后10 d,珠心突起細胞加厚轉變為珠心突起傳遞細胞(圖6B、E)。花后15~20 d,絕大部分珠心突起細胞分化為珠心突起傳遞細胞,細胞壁進一步加厚(圖6C、F、G、J),此時低溫處理組珠心突起傳遞細胞面積大于對照組(表2)。花后25~30 d,珠心突起傳遞細胞已經具有較厚的細胞壁,此時部分外層細胞出現皺縮,可以清楚地觀察到細胞殘體(圖6H、I、K、L),此時低溫處理組珠心突起傳遞細胞面積大于對照組。

A、B、C、G、H、I 分別為正常溫度下小麥穎果花后5、10、15、20、25、30 d的珠心突起傳遞細胞;D、E、F、J、K、L 分別為低溫處理下小麥穎果花后5、10、15、20、25、30 d的珠心突起傳遞細胞;NP:珠心突起;NPTC:珠心突起傳遞細胞。A,B,C,G,H,and I are the nucellar projection transfer cells at 5,10,15,20,25 and 30 d after flowering of wheat caryopsis under normal temperature,respectively; D,E,F,J,K and L are the nucellar projection transfer cells at 5,10,15,20,25 and 30 d after flowering in wheat caryopsis under low temperature treatment; NP:Nucellar projection; NPTC:Nucellar projection transfer cells.圖6 低溫處理對小麥穎果珠心突起傳遞細胞發育的影響Fig.6 Effect of low temperature treatment on the development of nucellar projection transfer cells in wheat caryopsis
以上結果表明,低溫處理組珠心突起傳遞細胞細胞壁較厚于對照組,且珠心突起傳遞細胞面積也較對照組更大。因此,低溫處理組具有較高效的養分卸載能力。
3 討 論
3.1 春季低溫對穎果形態發育的影響
在小麥生長發育過程中一旦出現春季低溫脅迫,小麥的單株產量會明顯下降,同時籽粒的品質也會發生改變[2]。此時小麥正處于拔節期至孕穗期,低溫會使由莖稈傳輸至葉片的同化物含量減少,從而導致干物質的積累量減少,最終降低產量[12]。路玉彥[13]發現,大麥經低溫處理后,其花器官生長受到負面影響,花藥變小,可育小花數下降,不育花粉數增加,穗部結實率降低;低溫處理在穎果發育后期也具有影響,當營養組織中的物質大量消耗之后,營養物質的運輸速率開始減緩,最終導致籽粒粒重低于正常處理。李玉剛等[14]研究發現,孕穗期低溫會導致冬小麥籽粒看起來干癟,粒寬減少,千粒重降低,這也與本研究結果相一致。
3.2 春季低溫對果皮消亡及胚乳發育的影響
果皮中淀粉主要貯藏在中果皮,隨著小麥穎果的生長發育,中果皮發生細胞程序性死亡,大量的細胞出現變形和破裂現象,同時釋放出淀粉粒[15]。在小麥發育過程中如遭遇到低溫脅迫等環境因素均會對果皮的發育造成一定的影響。本研究發現,在穎果發育前期,低溫處理組小麥穎果果皮細胞層數少于對照組,這可能是由于早期小麥為應對低溫環境,加速其生長發育,從而使得果皮凋亡速度提前,加快了果皮的發育進程。陳思思等[16]研究表明,低溫脅迫下葉片的凈光合速率、氣孔導度、蒸騰速率均下降,此時小麥果皮中的葉綠素合成也會受到影響,從而抑制果皮的發育進程。
胚乳的發育包含細胞數目的增加、細胞體積的擴大、淀粉體和蛋白體的充實等過程。淀粉體和蛋白體的發育狀況直接影響小麥產量和品質[17]。韋存虛等[18]研究表明,小麥胚乳中蛋白體大概于花后第8~9天開始出現。隨著胚乳細胞的發育,蛋白體不斷充實與積累,多個小蛋白融合形成一些較大的蛋白體。熊 飛等[19]認為,花后7 d,小麥胚乳中大淀粉的數量增長減慢,大約花后14 d開始出現小淀粉粒,且在籽粒成熟之前,小淀粉粒的數量以及大小都不斷增加。在本研究中,遭受低溫脅迫的小麥胚乳在花后10 d淀粉體數量顯著多于對照組,而腹部胚乳中的蛋白體數量低于對照組。但小麥胚乳細胞充實是一個動態變化過程,因此低溫脅迫對胚乳發育的影響可以用胚乳細胞中胚乳充實度所衡量。本研究結果表明,在發育早期,低溫對胚乳發育具有促進作用,但隨著對胚乳發育的減緩,低溫處理組的胚乳充實度低于對照組。
3.3 春季低溫對穎果養分運輸組織的影響
養分的輸送主要依靠腹部維管組織進行,葉片中合成的光合產物,需通過篩分子輸送到貯藏庫[20]。劉大同等[21]發現,維管束越發達,篩管面積就越大,韌皮部的卸載能力就越強。養分通過合點輸送至珠心突起細胞,珠心突起細胞間隙較大,有利于養分質外體的輸送[20],且珠心突起傳遞細胞質膜表面積的增加對穎果干物質的積累具有促進作用[22]。本研究通過切片觀察發現,在穎果早期發育過程中,低溫處理組珠心突起傳遞細胞壁較厚,細胞層數多且面積較大,因此能夠更高效地將來自維管束的養分運輸至胚乳腔中,最終進入籽粒。
