建甌萬木林省級自然保護區苦櫧種群種內與種間競爭
2020-03-08 06:06:10黃加林
武夷學院學報 2020年12期
黃加林
(光澤縣寨里林業工作站,福建 光澤 354100)
苦櫧(Castanopsis sclerophylla)為殼斗科栲屬常綠闊葉喬木樹種,栲屬植物中分布最北的一個種。苦櫧木材較密致、堅韌,富于彈性,是優良的用材樹種;樹冠濃密,有一定的觀賞價值,是良好的園林綠化樹種;脫澀后的果實(種仁)制作的苦櫧淀粉可加工成苦櫧豆腐、苦櫧粉絲和苦櫧糕等食品,是具有良好發展潛力的栽培樹種[1]。由于長期以來人們對天然闊葉林和次生林的過度采伐和破壞,天然林和次生林面積的日益減少,導致天然苦櫧資源日漸枯竭[2]。目前有關苦櫧的研究報道主要集中在天然林群落結構與植物多樣性[2]、苗木培育[3]、栽培技術[4]、果實的藥用價值[5]等方面,有關苦櫧次生林種間和種內競爭的研究未見報道。
在結構復雜的森林群落中,植物種群的結構和數量的變化受非生物因素(氣候、土壤等)和生物因素(種內和種間競爭)的共同制約,其中后者起著至關重要的作用[6]。由于環境資源是有限的,森林群落中的植物種群在生長發育過程中必然因爭奪有限的資源而發生種內和種間的競爭,導致群落植物組成、種群結構、植物多樣性的動態變化。因此,植物種內和種間競爭是生態學研究的核心問題[7]。競爭指數是反映植物間競爭程度的指標。徐道煒[8]提出的眾多的競爭指數模型可分為距離有關的和與距離無關的兩大類,其中與距離有關的Hegyi單木競爭指數模型能更好地反映種間和種內競爭程度,而被廣泛使用。筆者利用Hegyi單木競爭指數模型對福建萬木林自然保護區苦櫧群落的種間和種內競爭進行研究,揭示苦櫧種群的生態適應機制和動態變化,為苦櫧資源的科學保護和可持續利用提供參考資料。
1 試驗地概況
試驗地位于福建省建甌市房道鎮萬木林自然保護區的以苦櫧為優勢種的次生林群落,北緯27°02.987'~27°03.032',東經117°08.545'~117°08.685',海拔430~460 m左右,屬亞熱帶海洋性季風氣候,年均氣溫18.7℃,年均降水量1 673 mm,年均相對濕度約81%,年均無霜期290 d[2]。
2 研究方法
2.1 外業調查方法
在苦櫧次生林群落內連續設立20個10 m×10 m樣方,在每塊樣方中選取喬木層(樹高5 m及以上)苦櫧為對象木,測定其胸徑、樹高、冠幅、GPS坐標等指標。根據群落內喬木層大樹的冠幅確定競爭木為距離對象木8 m以內的所有喬木樹種[8]。以每株對象木為圓心,8 m為半徑設置樣圓,記錄樣圓內所有高度5 m及以上的喬灌木樹種(競爭木)的樹種名稱、胸徑、樹高、冠幅、GPS坐標、與對象木的水平距離等指標。
2.2 競爭指數模型的選擇
單木競爭指數模型種類較多,其中Hegyi提出的競爭指數模型使用簡便,數據容易獲得,在研究中得到較多的應用,本文采用該模型計算競爭指數[7-14]:

式中:CIi為第i株對象木(苦櫧)的競爭指數;Di為第i株對象木的胸徑;Dj為第j株競爭木(伴生樹種或苦櫧)的胸徑;Lij為對象木i與競爭木j之間的水平距離;n為第i株對象木的競爭木株數;CI為所有對象木(苦櫧種群)與競爭木的競爭強度;m為對象木的總株數。CI值越大,表明樹種間的競爭越激烈,對象木受到的競爭壓力越大。當競爭木全為苦櫧時,CIi為第i株對象木的種內競爭指數,CI為苦櫧種群的種內競爭強度;當競爭木全為伴生樹種時,CIi為第i株對象木的種間競爭指數,CI為苦櫧種群的種間競爭強度。
2.3 競爭指數與對象木胸徑的關系
樹種的競爭能力除了與其生物學、生態學特性和立地條件有關外,還與對象木個體大小有關。將每株對象木的平均競爭指數與其胸徑分別用倒數模型和冪函數模型等2個數學模型進行擬合,找出最佳的平均競爭指數與對象木胸徑的數學模型:

式中:CI為某對象木與所有競爭木的競爭指數的平均值,D為某對象木胸徑,A、B為模型參數。
3 結果與分析
3.1 對象木與競爭木概況
所有樣方的對象木(苦櫧)有21株,最小胸徑16.2 cm,最大胸徑83.2 cm,老樹較多,沒有小徑級的小樹和幼樹,種群處于老化和衰退階段。對象木30~35 cm和40~45 cm徑級的株數最多,均占總株數的19.05%,其他徑級的株數占比從大到小依次是:20~25 cm(14.29%)、35~40 cm(9.52%)、45~50 cm(9.52%),15~20、25~30、50~55、55~60、65~70和80~85 cm徑級的株數最少,均占總株數的4.76%見表1。

表1 苦櫧(對象木)和競爭木的徑階分布Tab.1 Diameter distribution of Castanopsis sclerophylla trees(objective trees)and their competitive trees
競爭木共294株,最小徑級平均胸徑4.6 cm,最大胸徑90.0 cm,由苦櫧、羅浮栲、木荷、栲樹、薄葉山礬、桂北木姜子、山礬、楓香、閩楠、檵木等38個樹種組成。競爭木的徑級分布與對象木不同,5~10 cm徑級的株數所占的比例最大,高達51.70%,其次是10~15 cm徑級,占競爭木總株數的14.97%,該徑級以上的競爭木株數總體上隨徑級的增大而減少。胸徑50 cm以上大徑級競爭木主要由楓香、苦櫧、馬尾松、木荷、羅浮栲、栲樹等樹種組成。
3.2 苦櫧種內與種間競爭
苦櫧種內競爭強度為8.712,種間競爭強度為27.038,占總競爭強度的75.63%,說明苦櫧的競爭主要來自種間,這主要與苦櫧徑級較大、株數較少、分布較分散有關。
不同徑級對象木受到的種內競爭強度均低于種間競爭強度,但種內單株平均競爭指數除了55~60 cm徑級外均高于種間競爭指數,這主要是由于苦櫧大徑級的比例遠高于伴生樹種。最小徑級(15~20 cm)對象木受到的競爭強度最大,種內和種間單株平均競爭指數分別為0.558和0.191,而最大徑級(80~85 cm)對象木受到的種內和種間單株平均競爭指數僅分別為0.158和0.020,。苦櫧種內和種間競爭強度總體上均有隨著徑級的增大而減小的趨勢,但也有一定的波動,尤其是種間競爭強度有較大幅度的波動(表2)。

表2 苦櫧的種內和種間競爭強度Tab.2 Intraspecific and interspecific competition intensity of Castanopsis sclerophylla
雖然苦櫧受到種內競爭強度(27.038)低于總的種間競爭強度,但其數值高于其他任何一種伴生樹種,見表3。在伴生樹種中,苦櫧與羅浮栲的競爭指數最大(5.431),其次是木荷(3.487)、栲樹(2.267)、薄葉山礬(1.829)、桂北木姜子 (1.744)、山礬 (1.696)、楓香(1.478)、閩楠 (1.109)、檵木 (0.857)、福建山礬(0.813)、欏木石楠 (0.649)、山黃皮 (0.566)、野柿(0.510)、浙江桂(0.453)、米櫧(0.409)、樹參(0.334)、華杜英(0.334)、栓葉安息香(0.323)、秀麗錐(0.318)、臺灣冬青(0.275)、虎皮楠(0.263)、青岡(0.236)、秀麗四照花(0.205)、榕葉冬青(0.178)等。
3.3 苦櫧對象木胸徑與競爭指數的關系
樹種間的競爭強度除了受樹種的生態習性、生活幅度和立地條件等多種因素制約外,對象木的個體大小對競爭強度有較大影響。采用倒數模型和冪函數模型擬合苦櫧對象木胸徑與平均單株競爭指數的關系,結果表明,苦櫧種內單株平均競爭強度與對象木胸徑有顯著的冪函數關系(P=0.039<0.05),與對象木胸徑倒數的關系不顯著(P=0.152>0.05);苦櫧與伴生樹種種間單株平均競爭強度與對象木胸徑有極顯著的冪函數關系(P=0.004<0.01),與對象木胸徑的倒數也有極顯著的線性相關關系(P=0.008<0.01),但冪函數關系更為密切;苦櫧與所有樹種的單株平均競爭強度與對象木胸徑的倒數有極顯著的線性相關關系(P=0.000<0.01),與對象木胸徑也有極顯著的冪函數關系(P=0.001<0.01),但與對象木胸徑的倒數線性相關關系更緊密,見表4、圖1。

表3 競爭木的種類組成、株數和競爭強度Tab.3 Composition of competitive trees and their competitive indices
從苦櫧對象木胸徑與競爭指數的關系數學模型可以看出,隨著對象木胸徑的增加,競爭指數急劇減小,對象木胸徑小于40 cm受到的競爭壓力較大,胸徑40 cm以上的對象木競爭指數較低。
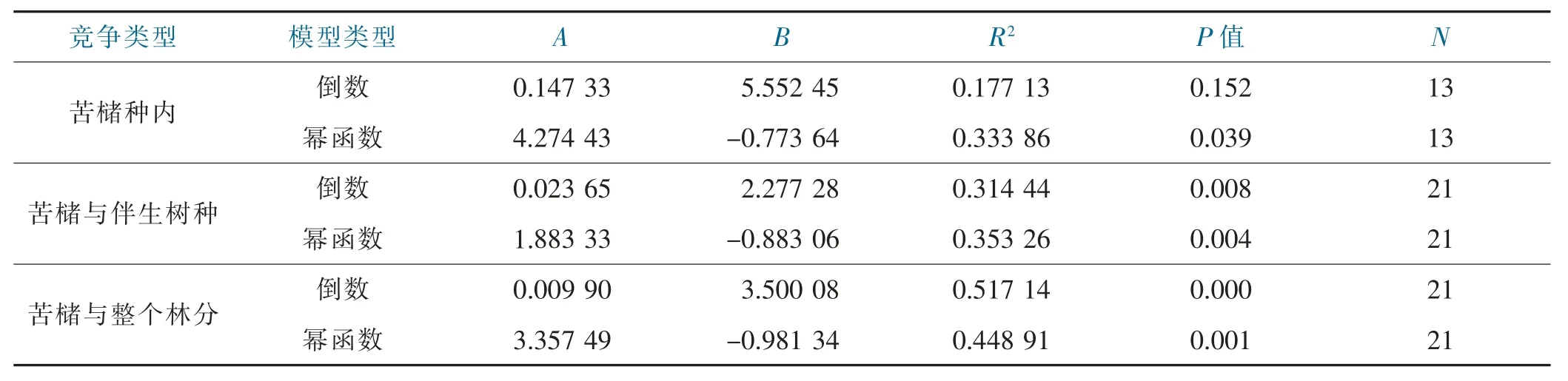
表4 對象木競爭強度與胸徑的回歸模型參數Tab.4 Parameters of CI-DBH models of object trees

圖1 對象木胸徑與種內、種間競爭指數與的關系Fig.1 Relationship between DBH of objective trees and intra-and interspecific competitions
4 小結與討論
眾多的研究表明,使用與距離有關的Hegyi單木競爭指數模型對森林群落中林木個體間的競爭進行定量研究,可更準確地度量林木個體間的競爭程度,進而描述和預測種群的動態變化[7-8]。
通過對苦櫧群落喬木層21株苦櫧對象木和294株競爭木的研究表明:喬木層苦櫧種內競爭強度為8.712,高于其受到的任何一種其他伴生樹種的種間競爭強度,但低于種間競爭指數總和(27.038),喬木層對象木的競爭壓力主要來自種間,這與項小燕等[10]對大別山五針松、曾思齊等[11]對湖南平江青岡櫟和李帥鋒等[12]對滇西北云南紅豆杉的種內和種間關系的研究結果相似,而與黃小波等[13]對云南云龍云南松和王曉霞等[14]對山西太岳山脫皮榆等優勢度較大的樹種的種內和種間關系的研究結果相反。苦櫧在群落中的株數雖少,但分布較為集中,徑階較大,這是苦櫧的種內競爭指數高于其受到的任何一種其他伴生樹種的種間競爭強度的最大的原因。
在伴生樹種中,喬木層羅浮栲的競爭指數最大(5.431),其次是木荷(3.487)。苦櫧對象木受到的種內和種間競爭強度的大小順序為:苦櫧>羅浮栲>木荷>栲樹>薄葉山礬>桂北木姜子>山礬>楓香>閩楠>檵木>福建山礬>欏木石楠>山黃皮>野柿>浙江桂>米櫧>樹參>華杜英>栓葉安息香>秀麗錐>臺灣冬青>虎皮楠>青岡>秀麗四照花>榕葉冬青等。
樹種間的競爭強度與各樹種的生物、生態學特性和生態幅度有關,對象木的種間和種內競爭強度還受到自身和競爭木胸徑、密度、所處生境等因素的共同影響。福建萬木林保護區的次生林的形成已有幾百年的歷史,在演替初期林下光照充足,喜光的馬尾松、楓香等先鋒樹種競爭力強而成為優勢樹種,但隨著群落的演替,林下光照強度逐漸減弱,先鋒樹種的幼苗幼樹耐陰性比常綠闊葉樹差,競爭力低,逐漸被淘汰。苦櫧次生林是次生林演替過程中最早出現的常綠闊葉林類型之一[15],苦櫧種群缺乏幼苗幼樹,主要原因是其幼苗幼樹較為喜光,目前的森林環境中林下光照不足,幼苗幼樹難以生長發育成小樹,再加上且其種子容易被鳥類和嚙齒目動物采食,導致能夠順利發育成幼苗的種子數量極少。
苦櫧種內和種間單株平均競爭強度隨對象木胸徑的增大而急劇減小,呈顯著的冪函數關系。對象木胸徑小于40 cm時受到的競爭壓力較大,對象木胸徑大于40 cm時受到的競爭強度較低且趨于穩定。一些處于林緣的苦櫧由于胸徑和冠幅較大,競爭壓力小,結果量大,因此,為了解決苦櫧天然更新能力差的問題,保護和開發現有的苦櫧資源,建議選擇生長性狀較好、結果量大的苦櫧優樹進行采果育苗繁殖,大力營造苦櫧人工林,加強苦櫧天然林資源的保護、開發和合理利用。
