IGF-1介導的豐富環境對成年弱視小鼠視皮層可塑性的影響
2020-02-08 05:07:26羅瑜琳羅詩詩劉政海王曦瑯陶利娟吳小影涂艷瓊
國際眼科雜志 2020年2期
羅瑜琳,羅詩詩,劉政海,王曦瑯,陶利娟,吳小影,涂艷瓊
0引言
弱視是一種嚴重影響視覺功能的常見兒童眼病。在視覺發育關鍵期內,患者如果得不到及時治療,不僅影響視力,還可能導致雙眼視覺以及立體視功能損害,嚴重危害家庭及社會。傳統觀念認為,超過視覺發育關鍵期的大齡兒童及成年弱視患者視功能及視皮層可塑性難以逆轉,治療無望。但近年來對弱視的研究使該觀點受到新的挑戰[1]。我們的前期研究已經證實,豐富環境可重塑視皮層突觸可塑性,改善成年弱視小鼠的視覺功能,然而其潛在的分子機制尚不十分清楚。胰島素樣生長因子-1(IGF-1)作為一種神經生長因子,具有神經保護及調控神經系統可塑性的作用,可作為研究視皮層可塑性的指標。本研究在建立成年單眼剝奪弱視模型小鼠的基礎上,觀察不同飼養環境對弱視小鼠視功能及視皮層可塑性的影響,重點探討IGF-1是否作為可能的關鍵因子參與弱視的發病,為臨床上弱視治療提供新的可行性思路。
1材料和方法
1.1材料
1.1.1實驗動物及分組選取正常新生21日齡昆明小鼠80只。根據隨機數字表法分為4組:正常組(Nor),單眼剝奪+標準環境組(MD+SE),單眼剝奪+豐富環境組(MD+EE),單眼剝奪+氟西汀組(MD+FLX)。所有小鼠采購于湖南省斯萊克景達實驗動物有限公司(No.43004700018257),許可證號:SCXK(湘)2016-0001。實驗得到湖南省兒童醫院倫理委員會的批準(審批號HCHLL-2015-35),實驗動物的處理按照《湖南省兒童醫院實驗動物管理與使用指南》實施。
1.1.2主要試劑及儀器全自動視覺電生理檢查系統(MetroVision,法國);2.5%戊二醛,1%鋨酸,日立H7700投射電鏡;IGF-1兔多克隆抗體(ab9572,Abcam,英國),IGF1-R兔多克隆抗體(ab39675,Abcam,英國),IGFBP5兔多克隆抗體(ab125696,Abcam,英國),TUBULIN小鼠單克隆抗體(sc-137179,Santa Cruz美國);乙酸纖維素薄膜(浙江路橋四青生化材料廠);電泳儀(北京六一,中國)。
1.2方法
1.2.1動物模型構建及處理各模型組(MD+SE、MD+EE、MD+FLX)小鼠于出生后21d縫合右側眼瞼,建立單眼形覺剝奪弱視模型[2],在小鼠麻醉狀態下,剪除右側上下眼瞼緣組織,然后行褥式縫合。在標準環境(standard enrichment, SE)中飼養2wk后剪開縫合的眼瞼,并對剝奪眼行閃光視覺誘發電位檢測正常眼及剝奪眼的P100波,剝奪眼的P100波幅較正常眼明顯下降,潛伏期明顯延長,確定建模成功。建模過程中眼瞼裂開或閃光視覺誘發電位檢測弱視模型失敗的小鼠予以剔除,每組保留18只小鼠進行后續實驗。剪開模型小鼠眼瞼后,按預先分組:MD+SE組在標準環境中飼養,MD+EE組在豐富環境中飼養,MD+FLX組在標準環境中飼養,但飲用水中添加氟西汀,4wk后行視敏度檢測及閃光視覺誘發電位檢測(F-VEP)后處死動物,取左側視皮層組織為后續檢測準備。
1.2.2環境干預方法豐富環境條件參照Baroncelli等[3]方法設置,采用1m3的木箱,其墊料上擺放隧道、轉輪、小木屋、各色積木及木梯等玩具,玩具隔天更換或交換位置,并定期清洗。MD+EE組小鼠每天置于該環境裝置中6h以上,持續4wk;標準環境則按國家標準實驗動物環境及設施規定的小鼠籠具原則要求飼養。
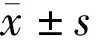
注:Nor:正常組;MD+SE:單眼剝奪+標準環境組;MD+EE:單眼剝奪+豐富環境組;MD+FLX:單眼剝奪+氟西汀組。
1.2.3小鼠視敏度檢測小鼠視敏度檢測方法參照Drapeau等[4]的描述,即前爪觸地反射實驗。操作方法[5]:用黑色的布遮擋小鼠的正常眼,將其從距離平臺大約20cm處緩慢下降,若小鼠在胡須或鼻子觸碰到桌面之前前爪出現探爪動作,則視為探爪成功,重復3次,以小鼠探爪成功率作為計量指標。該部分實驗檢測均由同一實驗人員完成,觀察與記錄均采取盲法。
1.2.4小鼠剝奪眼F-VEP檢測視敏度檢測完成后,參照ISCEV標準,對各組小鼠的右眼行F-VEP檢測,操作方法[6]:小鼠首先置于暗室適應30min,分別在枕骨粗隆、兩耳連線與正中線交點及同側耳處放置銀針電極。用閃爍光(刺激頻率1.0Hz,通頻帶寬0.5~85.0Hz)作為刺激光,疊加60次。檢查時左眼用不透光眼罩完全遮擋,記錄每次測量結果,包括P2波潛伏期及波幅,潛伏期為起點至P2波峰的距離,波幅為N2波谷至P2波峰的距離,連續測量3次,取其平均值。
1.2.5電鏡檢測小鼠視皮層神經元突觸超微結構處死小鼠取左側視皮層雙眼區組織,經2.5%戊二醛、磷酸鹽緩沖液固定2h,1%鋨酸固定液固定3h,體積分數50%、70%、90%乙醇脫水15min后,90%乙醇、體積分數90%丙酮等體積混合脫水15min,90%丙酮脫水15min,包埋液包埋,60℃烤箱固化,超薄切片機50nm厚切片,質量分數3%醋酸鈾-枸櫞酸鉛雙染色。然后將制備的切片于透射電子顯微鏡下觀察,每只小鼠隨機觀察記錄10個突觸,每組共記錄80個突觸進行圖像分析。參照多點平均法[7]及Güldner等[8]的方法測量并記錄各組小鼠視皮層神經元的突觸間隙寬度、突觸活性區長度及突觸后致密物厚度。
1.2.6 Western Blot法檢測小鼠視皮層中IGF-1、IGF-1R及IGFBP5蛋白表達分別收集各組小鼠左側視皮層雙眼區組織總蛋白,蛋白質變性后電泳,轉移至PVDF膜上,用含5%牛血清白蛋白的TBST封閉,用一抗IGF-1兔多克隆抗體(1∶500)、IGF-1R兔多克隆抗體(1∶500)、IGFBP5兔多克隆抗體(1∶1000)、TUBULIN鼠單克隆抗體(1∶1000),4℃孵育過夜,TBST洗3次,每次5min,二抗(HRP羊抗兔IgG,1∶1000,Vector)孵育1h,TBST洗3次,每次5min,化學發光法檢測蛋白質印跡結果。
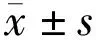
2結果
2.1豐富環境對弱視小鼠視敏度的影響通過檢測前爪觸地成功率判斷小鼠視敏度,各組(n=18)間小鼠前爪觸地成功率比較差異有統計學意義(F=114.632,P<0.001)。MD+SE組小鼠前爪觸地成功率(26.55%±4.66%)明顯低于Nor組(91.73%±2.98%),差異有統計學意義(t=15.480,P<0.001);MD+EE組(90.00%±7.86%)及MD+FLX組(87.43%±5.46%)小鼠前爪觸地成功率明顯高于MD+SE組,差異有統計學意義(t=15.071,P<0.001;t=14.841,P<0.001);與MD+FLX組比較,MD+EE組小鼠前爪觸地成功率差異無統計學意義(t=0.238,P=0.816)。
2.2豐富環境對弱視小鼠F-VEP的影響各組小鼠右眼(模型組小鼠剝奪眼)閃光視覺誘發電位檢測結果見圖1,表1。各組間小鼠F-VEP的P2波的潛伏期比較差異有統計學意義(F=36.510,P<0.01)。MD+SE組小鼠剝奪眼F-VEP的P2波的潛伏期較Nor組比較明顯延長,差異有統計學意義(t=10.220,P<0.01);MD+EE組及MD+FLX組小鼠剝奪眼F-VEP的P2波的潛伏期較MD+SE組比較均明顯縮短,差異有統計學意義(tMD+EE組 vs MD+SE組=7.051,PMD+EE組 vs MD+SE組=0.003;tMD+FLX組 vs MD+SE組=6.049,PMD+FLX組 vs MD+SE組=0.002);與MD+FLX組比較,MD+EE組小鼠剝奪眼F-VEP的P2波的潛伏期差異無統計學意義(t=1.002,P=0.322);各組間小鼠F-VEP的P2波的波幅總體比較差異有統計學意義(F=34.140,P<0.01)。MD+SE組小鼠剝奪眼F-VEP的P2波的波幅明顯低于Nor組,差異有統計學意義(t=10.090,P<0.01);MD+EE組及MD+FLX組小鼠剝奪眼F-VEP的P2波的波幅均明顯高于MD+SE組,差異有統計學意義(tMD+EE組 vs MD+SE組=5.030,PMD+EE組 vs MD+SE組<0.01;tMD+FLX組 vs MD+SE組=5.650,PMD+FLX組 vs MD+SE組P<0.01);與MD+FLX組比較,MD+EE組小鼠剝奪眼F-VEP 的P2波的波幅差異無統計學意義(t=0.618,P=0.539)。
2.3豐富環境對弱視小鼠視皮層神經細胞突觸超微結構的影響透射電鏡下觀察各組小鼠左側(模型組小鼠剝奪眼對側)視皮層雙眼區神經細胞突觸超微結構的結果見表1,圖2。各組間小鼠視皮層神經細胞突觸間隙寬度總體差異有統計學意義(F=8.75,P=0.004)。MD+SE組小鼠視皮層神經細胞突觸間隙明顯寬于Nor組,差異有統計學意義(t=5.018,P<0.01);MD+EE組及MD+FLX組小鼠視皮層神經細胞突觸間隙較MD+SE組明顯縮窄,差異有統計學意義(tMD+EE組 vs MD+SE組=3.24,PMD+EE組 vs MD+SE組=0.0035;tMD+FLX組 vs MD+SE組=2.712,PMD+FLX組 vs MD+SE組=0.0075)。各組間小鼠視皮層神經細胞突觸活性區長度總體差異有統計學意義(F=17.03,P<0.01)。MD+SE組小鼠視皮層神經細胞突觸活性區長度與Nor組比較明顯縮短,差異有統計學意義(t=5.772,P<0.01);MD+EE組及MD+FLX組小鼠視皮層神經細胞突觸活性區長度與MD+SE組比較均增加,差異有統計學意義(tMD+EE組 vs MD+SE組=4.805,PMD+EE組 vs MD+SE組<0.01;tMD+FLX組 vs MD+SE組=6.545,PMD+FLX組 vs MD+SE組<0.01)。各組間小鼠視皮層神經細胞突觸后致密物厚度總體差異有統計學意義(F=29.08,P<0.01)。MD+SE組小鼠視皮層神經細胞突觸后致密物厚度與Nor組比較明顯變薄,差異有統計學意義(t=8.609,P<0.01)。MD+EE組及MD+FLX組小鼠視皮層神經細胞突觸后致密物厚度較MD+SE組均增厚,差異有統計學意義(tMD+EE組 vs MD+SE組=7.181,PMD+EE組 vs MD+SE組<0.01;tMD+FLX組 vs MD+SE組=6.448,PMD+FLX組 vs MD+SE組<0.01)。MD+EE組與MD+FLX組小鼠比較,視皮層神經細胞突觸間隙寬度,突觸活性區長度,突觸后致密物厚度差異均無統計學意義(P>0.05)。
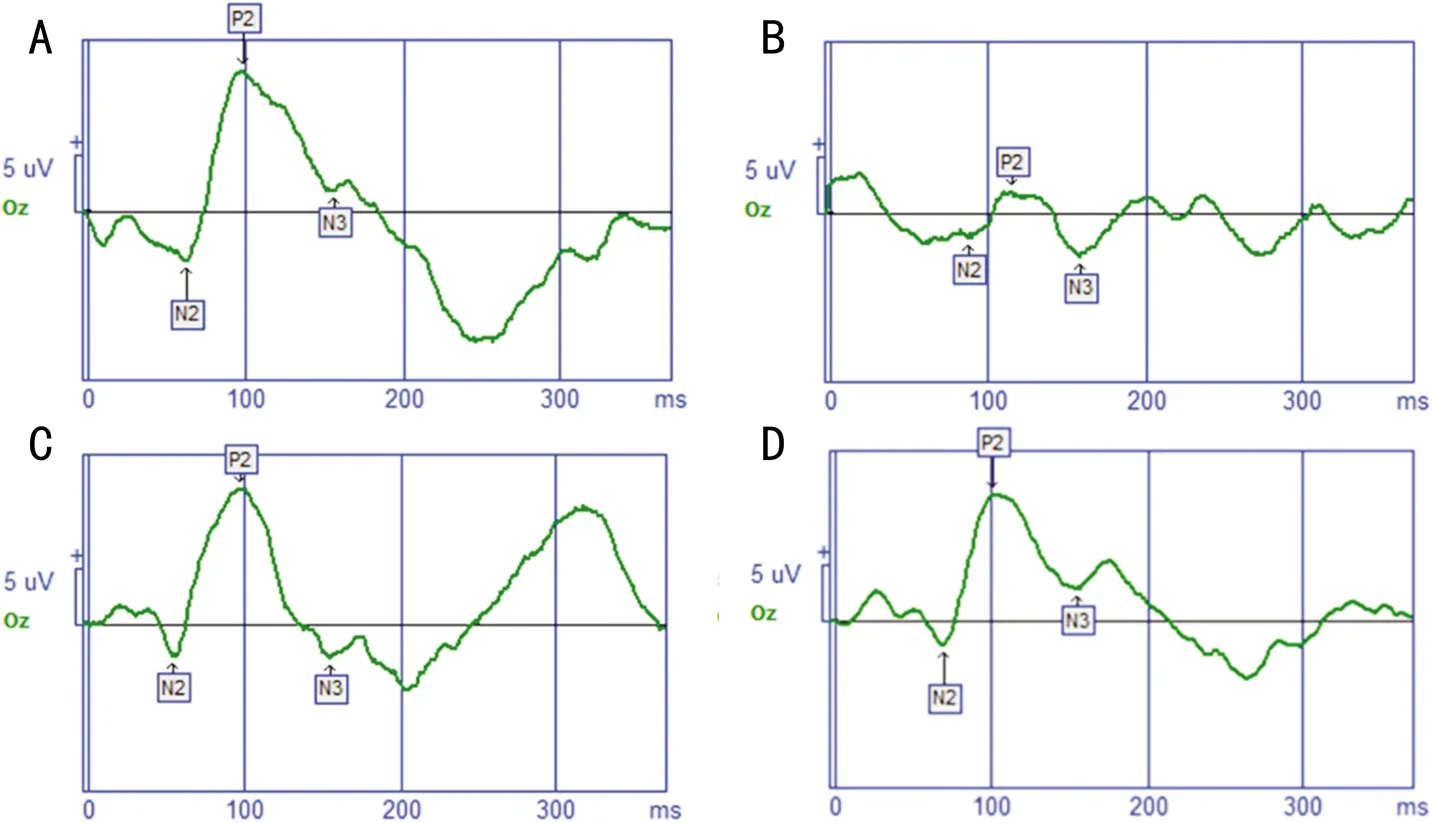
圖1各組小鼠閃光視覺誘發電位P2波潛伏期及波幅比較A: Nor組;B:MD+SE組;C:MD+EE組;D:MD+FLX組。
圖2電鏡觀察各組小鼠視皮層神經細胞突觸超微結構A:Nor組;B:MD+SE組;C:MD+EE組;D:MD+FLX組。
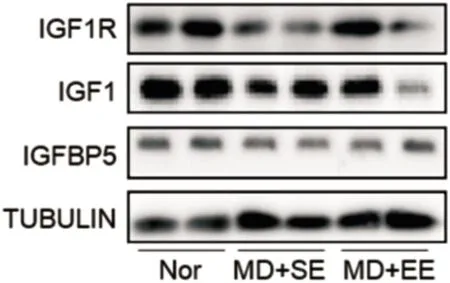
圖3 Western-blot檢測各組小鼠視皮層IGF-1、IGF-1R及IGFBP5蛋白表達。
2.4豐富環境對弱視小鼠視皮層IGF-1、IGF-1R及IGFBP5蛋白表達的影響Western Blot檢測各組小鼠左眼(模型組小鼠剝奪眼對側)視皮層雙眼區組織的IGF-1及IGF-1R蛋白表達見表2,圖3,各組間小鼠視皮層雙眼區組織IGF-1蛋白表達光密度值總體差異有統計學意義(F=22.00,P<0.01)。MD+SE組小鼠IGF-1蛋白表達明顯低于Nor組,差異有統計學意義(t=6.598,P<0.01); MD+EE組小鼠IGF-1蛋白的表達明顯高于MD+SE組,差異有統計學意義(t=2.712,P= 0.016),但仍顯著低于Nor組,差異有統計學意義(t=3.887,P=0.001)。各組間小鼠視皮層雙眼區組織IGF-1R蛋白表達光密度值總體差異有統計學意義(F=67.31,P<0.01)。MD+SE組小鼠IGF-1R蛋白表達明顯低于Nor組,差異有統計學意義(t=11.4,P<0.01); MD+EE組小鼠IGF-1R蛋白表達明顯高于MD+SE組,差異有統計學意義(t=3.815,P=0.041),但仍顯著低于Nor組,差異有統計學意義(t=7.582,P=0.001)。各組間小鼠視皮層雙眼區組織IGFBP5蛋白表達光密度值總體差異無統計學意義(F=0.793,P=0.470)。
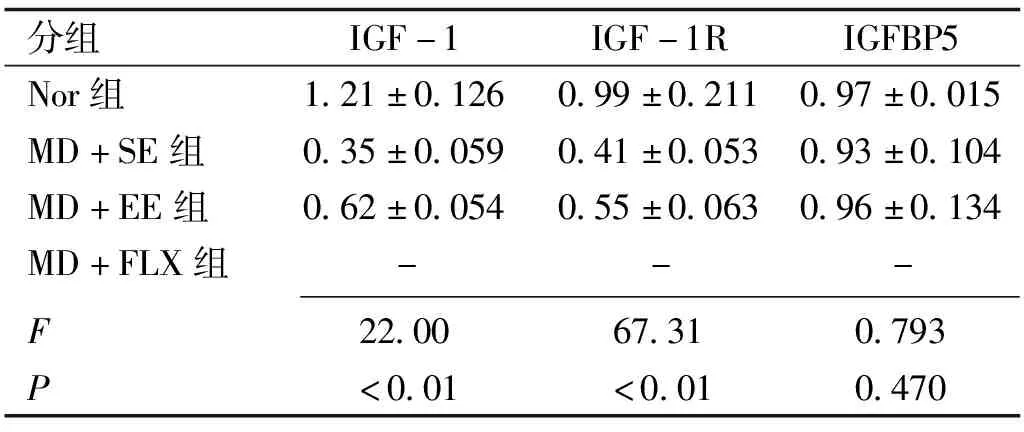
分組IGF-1IGF-1RIGFBP5Nor組1.21±0.1260.99±0.2110.97±0.015MD+SE組0.35±0.0590.41±0.0530.93±0.104MD+EE組0.62±0.0540.55±0.0630.96±0.134MD+FLX組---F22.0067.310.793P<0.01<0.010.470
注:Nor:正常組;MD+SE:單眼剝奪+標準環境組;MD+EE:單眼剝奪+豐富環境組;MD+FLX:單眼剝奪+氟西汀組。
3討論
人類和哺乳動物出生以后,視覺系統能夠根據周圍視覺環境的刺激調整和改變視皮層神經元的結構和突觸間聯系,稱為視皮層的可塑性,發生這一改變的時期稱為視覺發育的關鍵期[9]。隨著年齡的增長,視皮層神經網絡結構逐漸發育成熟,限制了神經元突觸軸突及樹突的生長,可塑性逐漸被抑制。臨床上弱視治療具有顯著的時限性,視皮層可塑性關鍵期內療效明顯,超過關鍵期的患者采用傳統的遮蓋療法效果不佳。因此如何重新激活超越關鍵期“被抑制”的視皮層可塑性是目前研究的熱點和難點。
單眼形覺剝奪弱視模型是目前研究視皮層可塑性的經典模型[10]。本實驗對出生后21d的小鼠行右側眼瞼縫合阻斷該側視覺信息輸入,建立單眼剝奪弱視模型。單眼縫合2wk后,我們觀察模型小鼠前爪觸底反射成功率顯著降低,剝奪眼F-VEP P2波的潛伏期延長、波幅下降,證實單眼剝奪弱視小鼠模型建立成功。
豐富環境是研究感覺經驗對大腦和行為影響的一個廣泛應用的模型,它被定義為“復雜的無生命物與社會刺激的復合體”[11]。豐富環境不僅提供了多感官刺激和運動的機會,而且賦予了相互間社會性情感體驗的可能。近年來,豐富環境逐漸被廣泛應用于中樞神經系統以及視皮層可塑性機制的研究中[12]。將正常新生小鼠暴露于豐富環境中讓其生長,出生后25d視敏度較標準環境中生長的小鼠增加18%,其視敏度基本接近出生后28d的水平,說明豐富環境顯著影響著視覺系統的發育,可加速小鼠視功能的成熟[13-14]。我們前期研究證實,豐富環境可通過上調成年單眼剝奪弱視小鼠視皮層中MAP2的表達,調控突觸相關蛋白PSD-95及SYP重塑視皮層可塑性,從而促進弱視模型小鼠視覺功能的恢復。本實驗以氟西汀干預組作為陽性對照,進一步探討豐富環境重新激活成年視皮層可塑性的分子機制。結果顯示,在單眼剝奪弱視小鼠模型建立成功以后,將其飼養于豐富環境中,4wk后應用前爪觸地反射實驗觀察各組小鼠的主觀視敏度,顯示豐富環境飼養組模型小鼠前爪觸地成功率明顯高于標準環境飼養組,與正常小鼠及氟西汀干預的弱視模型小鼠比較無明顯差異,提示豐富環境明顯改善弱視模型小鼠的主觀視敏度。我們同時使用F-VEP檢測各組小鼠的客觀視功能,與標準環境飼養比較,豐富環境中飼養的弱視模型小鼠剝奪眼F-VEP的P2波潛伏期明顯縮短,波幅升高,小鼠客觀視功能恢復并接近氟西汀治療組。近來的多項研究結果證實,氟西汀對包括海馬[15]、軀體感覺皮層椎體神經元[16]、視皮層[17-18]在內的成年皮層神經元可塑性具有顯著的再激活作用,從而為成年弱視治療提供新的靶點,因此在我們的研究中將氟西汀干預組作為陽性對照。以上實驗結果均提示豐富環境可以重新激活成年弱視小鼠的功能可塑性,顯著改善視覺功能,其機制可能是通過重塑視皮層神經元突觸結構實現的。
以往研究證實,豐富環境可引起正常成年或衰老動物中樞神經系統結構可塑性的變化,包括皮層重量和厚度、神經細胞數量、樹突分支及密度、突觸數量及突觸接觸面積等的變化[19-20]。我們的研究顯示,與標準環境相比較,豐富環境飼養條件下的成年弱視模型小鼠,剝奪眼對側視皮層突觸間隙縮小、突觸活性區長度增加、突觸后致密物厚度增加;突觸超微結構參數與正常小鼠及氟西汀干預的弱視模型小鼠比較無明顯差異,提示豐富環境可重新激活成年弱視小鼠的結構可塑性,改變突觸結構,促進視皮層視覺電沖動的傳導。突觸是神經元間信息傳遞的結構基礎,豐富環境干預以后,視皮層神經元突觸結構發生適應性變化,突觸間隙縮小,加快神經遞質到達突觸后膜的速度,神經遞質失活的可能性降低,增強神經元與效應細胞之間的信息溝通,加強突觸傳遞效能。突觸界面活性區長度在一定程度上反映神經元功能活動狀態,活性區長度的增加使突觸前后膜接觸面增加,減少遞質向周圍間隙擴散,有利于遞質與受體更有效結合。PSD是突觸后膜上的致密物質,主要含有微管蛋白、肌動蛋白、神經絲蛋白及各種酶,豐富環境飼養后的弱視小鼠視皮層神經元突觸后致密物厚度顯著增加能反映出突觸后膜上受體與離子通道的變化,從而在功能上增強突觸的傳遞效率,改善小鼠視覺功能。
胰島素樣生長因子-1(IGF-1)是胰島素樣生長因子家族中的一員,是由70個氨基酸組成的單鏈堿性多肽,主要由肝臟合成及分泌,具有廣泛的生物學效應。IGF-1的生物學活性受IGF-1受體(IGF-1R)和胰島素樣生長因子結合蛋白(IGFBP)的調節。血漿中75%的IGF-1都和IGFBP結合形成無活性的復合物,構成IGF-1的儲備庫。組織中的IGFBP通過阻止IGF-1與其受體結合,抑制IGF-1發揮生物學作用。此外,IGF-1還可穿過血-腦屏障,與廣泛分布于腦組織中的IGFBP5結合形成無活性的復合物,游離的IGF-1則分布于腦組織中與IGF-1R結合而發揮作用[21]。經典的理論認為,IGF-1與哺乳動物中樞神經系統的發生密切相關,包括調控神經細胞增殖及發生,神經元存活、分化,突觸發生,髓鞘形成等[22]。近年來研究報道,IGF-1不僅能對神經細胞起到保護作用[23],而且可介導并調控海馬以及視皮層可塑性[24-25],參與學習、記憶及弱視的發病機制。本實驗結果證實,IGF-1、IGF-1R及IGFBP5在正常小鼠視皮層組織中均有表達,單眼剝奪后剝奪眼對側視皮層中IGF-1及IGF-1R表達明顯下調,IGFBP5表達無明顯變化。豐富環境干預后較標準環境飼養小鼠視皮層中IGF-1,IGF-1R表達明顯上調,但并不影響IGFBP5的表達。以上結果均提示豐富環境可能通過直接上調IGF-1及受體IGF-1R表達,介導對成年弱視小鼠視皮層可塑性進行調控,重新激活“被抑制”的視皮層神經環路,改變突觸結構,增強突觸傳遞效能,在一定程度上改善弱視小鼠的視覺功能。我們推測IGF-1調控視皮層可塑性機制包括以下幾種:(1) IGF-1通過調控葡萄糖代謝及體內血糖水平,調控視皮層組織、細胞及突觸對外界感覺刺激的反應;(2) IGF-1作為視皮層中突觸及神經細胞活動的重要調質,發揮神經營養因子的作用調控視皮層可塑性;(3) IGF-1通過影響PI3K的表達,調控神經細胞傳導及突觸蛋白表達,重新激活視皮層可塑性。因此,在未來的研究中,我們將對以上可能機制進行深入探究,力求尋找能適合臨床大齡及成年弱視患者治療的有效性、無創性新方法。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
新少年(2022年9期)2022-09-17 07:10:54
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
北極光(2014年8期)2015-03-30 02:50:51
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
