油橄欖果實油脂積累的轉錄組分析
2020-01-16 00:46:22耿樹香寧德魯李勇杰陳海云
中國油脂 2019年12期
耿樹香,寧德魯,李勇杰,陳海云
(云南省林業(yè)科學院,昆明 650201)
油橄欖是世界著名的優(yōu)質木本油料樹種,是木犀科木犀欖屬的小喬木。栽培油橄欖OleaeuropaeaL.是從野生種OleachrysophyllaLam.經(jīng)過OleaoleasterL.或Oleaeuropaeaoleaster進化而來的。橄欖油是油橄欖中最有價值的部分,其富含不飽和脂肪酸及維生素、類胡蘿卜素、多酚類化合物和有機酸等[1-2]。目前,國外對油橄欖基因組方面的研究做了很多工作[3-10]。如對油橄欖栽培種萊星進行測序、組裝,獲得完整基因組;對油橄欖栽培種Farga使用RNA序列數(shù)據(jù)進行注釋,產(chǎn)生了56 000多個蛋白質編碼基因,同時進行測序、組裝,獲得完整基因組;研究發(fā)現(xiàn)結合siRNA中的一個油橄欖OD基因隨其積累可導致FAD2不表達,抑制FAD2基因表達的結果是可導致油橄欖果中油酸含量增高。國內近年來對木本油料油脂合成代謝途徑的轉錄組基因組學研究越來越多[11],如林萍[12]、Xia[13]等對普通油茶種子進行轉錄組測序分析,陳昊等[14]對油桐種子3個不同發(fā)育時期轉錄組進行分析,丁健[15]對沙棘果油脂積累轉錄組進行了系統(tǒng)的分析。目前,對影響油橄欖油脂合成的關鍵基因及途徑尚未清晰,特別是油橄欖果中髙積累油酸的機制未見報道。對油橄欖果油脂和脂肪酸組分的動態(tài)變化規(guī)律,以及發(fā)育過程中相關功能基因的轉錄表達模式進行研究,可為通過現(xiàn)代組學技術探索油橄欖油脂合成積累機理研究奠定基礎,也為進一步提高油橄欖油脂含量提供技術支撐。
1 材料與方法
1.1 實驗材料
以云南永仁引種油橄欖佛奧(FA)及鄂植8號(EZ)品種為實驗對象,取2個油橄欖品種授粉后60 d(5月)、120 d(7月)、180 d(9月),分別在所選油橄欖品種株樣樹冠中部的東、西、南、北4個方位隨機采摘果實10個,立即置于液氮中,于-80℃冰箱保存,備用。
重蒸水;CTAB、水飽和酚、三氯甲烷、異戊醇、無水乙醇,分析純;安捷倫RNA提取試劑盒、TruSeq RNA樣品準備試劑盒,美國Illumina公司;超級II逆轉錄酶,美國Invitrogen公司;AgencourtAMPure XP-Medium,美國Agencourt公司;TaKaRa Ex Taq Hot Start版、HiSeq 4000 SBS工具包、HiSeq 4000 PE Cluster Kit,美國Illumina公司。
熒光定量PCR儀,瑞士Roche;冷凍混合球球磨儀,德國萊馳;日本奧林巴斯顯微成像系統(tǒng);Nano drop 2000超微量紫外可見分光光度計,美國Thermo公司;Illumina genome Analyzer測序儀、Cluster generation、HiSeq System,美國Illumina公司;Agilent 2100 Bioanalyzer,美國安捷倫公司;Eppendorf高速低溫離心機、全套Eppendorf微量移液器。
1.2 實驗方法
1.2.1 油橄欖果實RNA的提取
油橄欖果實RNA的提取方法按照TIANGEN?多糖多酚植物總RNA提取試劑盒說明書進行。
1.2.2 cDNA文庫構建和測序
油橄欖佛奧(FA)及鄂植8號(EZ)的轉錄組測序由成都生命基線科技有限公司完成。提取的樣品總RNA用DNasel消化基因組DNA,經(jīng)質檢合格后,用帶有Oligo(dT)的磁珠富集真核生物mRNA;加入打斷試劑在恒溫混勻儀中將mRNA打斷成150~200 nt的短片段,以短片段mRNA為模板,用6堿基隨機引物合成一鏈cDNA;然后配制二鏈合成反應體系合成二鏈cDNA,并利用試劑盒純化雙鏈cDNA;純化后的cDNA先進行黏性末端修復,隨后在cDNA的3′末端加上堿基A并連接接頭,最后進行片段大小選擇和PCR擴增;構建好的文庫用Agilent 2100 Bioanalyzer和ABI StepOnePlus Real-Time PCR Syst實時熒光定量系統(tǒng)質檢合格后,進行測序。
測序得到的原始數(shù)據(jù)經(jīng)過濾和數(shù)據(jù)質控后得到高質量序列數(shù)據(jù),使用HISAT2軟件將高質量序列數(shù)據(jù)比對到參考基因/基因組序列后,統(tǒng)計其在二者上的分布情況,比對質控合格后才能進行定量分析、功能富集分析、可變剪接、新轉錄組本預測及注釋、SNP/InDel檢測和基因融合等后續(xù)一系列分析。
1.2.3 轉錄組基因功能注釋和表達分析
參考油橄欖基因組數(shù)據(jù)庫進行功能注釋。基因表達量的計算采用FPKM(Fragments per Kilobase Million),即每100萬個比對上的測序序列到外顯子的每1 000個堿基上的可讀序列個數(shù)。在RNA-Seq分析中,可通過對測序序列的計數(shù)來估計基因的表達水平。一個基因表達水平的直接體現(xiàn)就是其兩端不能再延長的序列的豐度情況,兩端不能再延長的序列豐度程度越高,則基因表達水平越高。Reads計數(shù)除了與基因的真實表達水平成正比外,還與基因的長度和測序深度呈正相關。
1.2.4 差異基因表達分析
差異基因表主要用于考察各發(fā)育階段間的基因表達差異,主要參照Audic法。在得到差異檢驗的錯誤發(fā)現(xiàn)率(False discovery rate,F(xiàn)DR)值的同時,根據(jù)基因的表達量(FPKM)計算該基因2個樣本間的差異表達倍數(shù)。FDR值越小,差異倍數(shù)越大,則表示表達差異越顯著。在本研究數(shù)據(jù)分析中,將基因表達矩陣分為隨機組和對照組,使用DEGSeq軟件對差異基因表達進行分析,對差異檢驗的概率作多重假設檢驗校正,通過控制FDR來決定概率的域值,分析中取p不大于0.05為篩選差異基因的閾值。
1.2.5 差異基因表達Pathway及GO分析
以油橄欖基因組數(shù)據(jù)為參考基因進行構建人工比對數(shù)據(jù)庫,以佛奧和鄂植8號測得的具有編碼蛋白序列的289、593、580個兩端不能再延長的序列基因在KEGG數(shù)據(jù)庫中進行功能注釋,并運用人工比對進行功能分類。Pathway分析同時還借助GO和MapMan(3.5.1R2)進行分析。
2 結果與分析
2.1 兩種油橄欖品種的含油率
佛奧(FA)和鄂植8號(EZ)的鮮果在不同的成熟時期果實的形態(tài)差異明顯,參照GB/T 14772—2008中提油方法,對佛奧和鄂植8號的鮮果在果實成熟階段的60 d(5月)、120 d(7月)、180 d(9月)對鮮果含油率進行測定,結果如圖1所示。
從圖1可以看出,佛奧的含油率顯著高于鄂植8號。伴隨著果實成熟,果實顏色逐漸變黑,2個油橄欖品種含油率顯著增加,呈上升趨勢。因此,本研究選取成熟過程中的佛奧和鄂植8號的鮮果進行轉錄組測序,探究參與油橄欖脂肪酸代謝途徑的表達差異基因,通過轉錄組分析解析2個品種間含油率差異的原因。

圖1 佛奧(FA)與鄂植8號(EZ)油橄欖鮮果含油率
2.2 RNA質量及濃度檢測
經(jīng)檢測,提取2個品種3個時期油橄欖鮮果的RNA樣品28S∶18S值均在0.5~1.3間,反應RNA完整性的RIN值大于7.5,初提質量良好,符合進一步檢測并測序的要求。濃度和純度檢測結果顯示大部分樣本RNA提取完整性好,無降解,無蛋白、多糖、多酚和DNA等雜質污染,完全達到文庫構建要求(RIN值>7)。
2.3 Illumina雙末端測序以及denovo組裝(見表1)

表1 油橄欖成熟過程中果實轉錄組基本數(shù)據(jù)
由表1可知,通過Illumina測序,F(xiàn)A5得到46 764 072條高質量序列數(shù)據(jù),F(xiàn)A7得到49 780 566條高質量序列數(shù)據(jù),F(xiàn)A9得到49 689 152條高質量序列數(shù)據(jù),EZ5得到46 646 112條高質量序列數(shù)據(jù),EZ7得到49 435 120條高質量序列數(shù)據(jù),EZ9得到47 278 558條高質量序列數(shù)據(jù),平均長度在327 bp。將2個品種的成熟過程中的高質量序列數(shù)據(jù)比對到油橄欖基因組,F(xiàn)A5、FA7、FA9、EZ5、EZ7、EZ9基因組分別達到55.48%、55.86%、45.43%、55.39%、54.79%、58.22%的匹配,所比對到的表達基因分別為26 231、26 079、25 292、27 255、25 994、25 344個,所比對的測序所得序列與基因序列達到35.29%、37.99%、27.13%、34.41%、35.71%、37.32%的匹配。兩端不能再延長的序列長度在200 bp以上。
2.4 差異基因表達分析(見圖2)
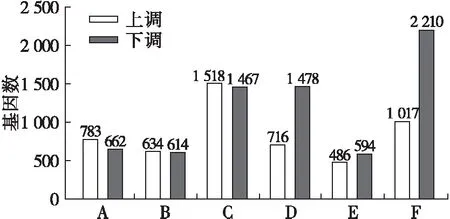
注:A、B、C、D、E、F分別為FA7-vs-FA5、FA9-vs-FA7、FA9-vs-FA5、EZ7-vs-EZ5、EZ9-vs-EZ7、EZ9-vs-EZ5。
圖2 油橄欖成熟過程中果實差異基因統(tǒng)計
從圖2可以看出,佛奧的不同月份發(fā)育時期之間差異基因的表達(上調或下調)變化不明顯,而鄂植8號5月與7月、5月與9月的差異基因的表達差異較大,下調的基因數(shù)量是上調基因數(shù)量的兩倍,推測可能與果實的成熟和發(fā)育相關。顎植8號油橄欖品種在果實成熟初期及末期差異基因數(shù)較多,說明其成熟前后,差異基因表達較多,這與果實成熟規(guī)律趨于一致,即果實成熟初期,細胞分裂旺盛,內含物質新陳代謝旺盛,而發(fā)育后期,油脂累積趨于穩(wěn)定所致。
2.5 油橄欖油脂生物合成途徑中關鍵基因的表達(見表2)

表2 油橄欖油脂生物合成途徑中關鍵基因的表達
從表2可以看出:橄欖油生物合成途徑中第一步的關鍵酶是生物素羧基載體蛋白(Biotin carboxyl carrier proteins,BCCP),EZ7的基因表達最高,F(xiàn)PKM為3 338.30,F(xiàn)A5最低,F(xiàn)PKM為482.96,F(xiàn)A7為FA中表達量最高的,F(xiàn)PKM為2 351.68;第二步反應中的β-酮酰基ACP還原酶(Beta-ketoacyl-ACP reductase,F(xiàn)ABG)基因表達量最高的是EZ7,F(xiàn)PKM為3 063.45,F(xiàn)A5最低,F(xiàn)PKM為811.36,F(xiàn)A7為FA中表達量最高的,F(xiàn)PKM為2 494.31;β-羥酰ACP脫氫酶(Beta-hydroxyacyl-ACP dehydrases,F(xiàn)ABZ)基因表達量最高的是EZ7,F(xiàn)PKM為2 708.94,F(xiàn)A5最低,F(xiàn)PKM為657.02,F(xiàn)A7為FA中表達量最高的,F(xiàn)PKM為1 547.22。在油橄欖的整個成熟期,兩品種油橄欖果實在7月脂肪酸的生物合成中關鍵酶基因表達量最高。
脂肪酸去飽和酶有許多種,可以將其分為兩大類,一類在脂肪酸形成甘油酯之前引入第一個雙鍵時起作用,僅包含一種酶,即硬脂酰ACP去飽和酶(stearoyl-ACP desaturase,SACPD),主要存在于質體中,是唯一的一種可溶性去飽和酶,另外一類是形成甘油酯之后,在脂肪酸基團進一步去飽和過程中起作用,包括油酸去飽和酶(FAD2-4和FAD2-5)和亞油酸去飽和酶(FAD3-3)。
油橄欖不飽和脂肪酸形成相關酶中omega-6油酸去飽和酶(FAD2)基因表達量最高的是FA5,F(xiàn)PKM為415.47,F(xiàn)A7基因表達量最低,為70.12,EZ5為EZ中表達量最高的,F(xiàn)PKM為366.46,兩品種均為5月樣品具有高表達量的FAD2。油橄欖果中的β-酮酰基ACP合成酶(Beta-ketoacyl-ACP synthases,KAS)中的KASI基因是啟動脂肪酸合成的關鍵酶,在鄂植8號果實的3個發(fā)育時期,隨油橄欖成熟度增加,油橄欖果中的KAS基因的表達水平不斷增加,EZ5中FPKM為227.66,EZ9為416.10,KAS基因的表達水平影響橄欖油中脂肪酸的合成。
2.6 油脂合成基因的表達分析(見表3)
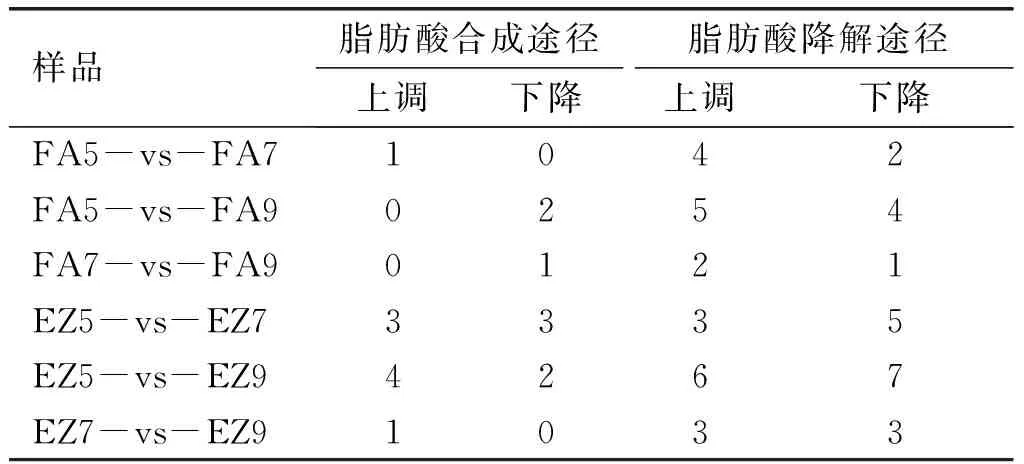
表3 2個品種不同發(fā)育相鄰時期脂肪酸合成與降解差異基因表達
在6個油橄欖樣本中總共檢測出1 886個基因具有表達。由表3可知:佛奧果實FA5-vs-FA7發(fā)育時期,脂肪酸合成路徑乙酰輔酶A合成丙二酰輔酶A,1個基因發(fā)生了上調表達,而在脂肪酸降解途徑中有4個基因發(fā)生了上調表達,2個基因發(fā)生了下調表達;FA7-vs-FA9時期,脂肪酸合成途徑上有1個基因發(fā)生下調表達,沒有注釋到上調表達的候選基因,脂肪酸降解途徑發(fā)生了1個基因的下調表達,2個基因的上調表達;FA5-vs-FA9時期,脂肪酸合成路徑上被注釋到的基因有2個發(fā)生了下調表達,而沒有上調表達的基因,脂肪酸降解途徑被注釋到的基因有5個發(fā)生了上調表達,有4個發(fā)生了下調表達;鄂植8號果實EZ5-vs-EZ7時期,在脂肪酸合成路徑上被注釋到的基因有3個發(fā)生了上調表達,有3個發(fā)生了下調表達,脂肪酸降解途徑上被注釋到的基因有3個發(fā)生了上調表達,有5個發(fā)生了下調表達;EZ7-vs-EZ9時期,脂肪酸合成路徑上注釋到的基因有1個發(fā)生了上調,沒有注釋到下調基因,脂肪酸降解途徑上注釋到的基因有3個發(fā)生了上調表達,有3個發(fā)生了下調表達;EZ5-vs-EZ9時期,脂肪酸合成路徑上注釋到的基因有4個發(fā)生了上調表達,有2個基因發(fā)生了下調表達,脂肪酸降解途徑上注釋到的基因有6個發(fā)生上調表達,有7個發(fā)生下調表達。
2.7 油脂合成基因的聚類分析
根據(jù)差異基因檢測結果,使用gplots軟件包進行層次聚類分析。對1 886個基因進行篩選,將FPKM 值低于200的差異基因去除,對篩選后的208個與油脂合成相關的基因采用Omicshare在線工具(http://www.omicshare.com)作圖并進行聚類分析,結果見圖3。

圖3 油橄欖果與油脂合成相關基因聚類分析
從圖3可以看出,2個品種油橄欖參與油脂合成的差異基因表達模式在5月較為相似,每個品種的7月和9月差異基因的表達模式較為相似,果實成熟后期(7月和9月)與果實生長初期(5月)相比,大多數(shù)基因發(fā)生了表達量上調,可能與果實的成熟有關。
為尋找脂肪合成的相關基因,以脂肪合成初期(FA5)與合成盛期(FA9)的表達譜比對,分析比對脂肪酸合成通路的差異基因,包括脂肪酸合成通路、脂肪酸鏈延伸通路以及不飽和脂肪酸的合成通路等。結果顯示:在脂肪合成盛期上調表達的基因均為與橄欖油脂肪酸合成相關的基因,有β-酮酰基ACP合成酶、硬脂酰ACP去飽和酶(SACPD)等。
3 結 論
(1)以含油率差異顯著的2個品種(高含油率品種佛奧(FA),9月油橄欖果含油率為23.16%和低含油率品種鄂植8號(EZ),9月含油率為15.17%)為材料,轉錄組分析探究了其果實發(fā)育3個關鍵時期(5月、7月和9月)基因表達與油脂積累的關系。研究發(fā)現(xiàn),在FA7-vs-FA5、FA9-vs-FA7、FA9-vs-FA5、EZ7-vs-EZ5、EZ9-vs-EZ7、EZ9-vs-EZ5 6個階段中分別找到783、634、1 518、716、486和1 017個差異基因上調,662、614、1 467、1 478、594和2 210個差異基因下調。
(2)在脂肪酸代謝途徑中富集到的KAS、FAD、BCCP、FABG、EAR等基因可能是油橄欖果油脂和脂肪酸生物合成關鍵基因,其后續(xù)通過表達載體構建和蛋白異源表達等對基因功能進行驗證,為進一步闡明油橄欖果油脂合成途徑及其關鍵酶的功能,揭示油橄欖果油脂合成的分子機理,進而為建立油橄欖果油脂異源合成系統(tǒng)奠定基礎,而不同時期油橄欖果差異基因表達的掌握對研究不同組織中生物活性成分的品質形成機制具有重要作用。
(3)脂肪酸合成、甘油酯代謝和甘油磷脂代謝等途徑是影響油橄欖油脂合成的關鍵途徑,在FA-vs-EZ的脂類代謝途徑中,僅有脂肪酸生物合成途徑中的差異基因表達呈顯著富集,表明油橄欖果油脂在從頭合成中即產(chǎn)生明顯差異,油橄欖果中與脂肪酸合成相關的關鍵基因的表達水平均隨果實成熟度的增加呈上升趨勢。
猜你喜歡
食品工業(yè)(2023年12期)2023-12-12 12:56:16
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
中學生數(shù)理化·七年級數(shù)學人教版(2019年10期)2019-11-25 07:33:58
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中學生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國糧油學報(2018年12期)2018-01-26 13:32:04
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
中國衛(wèi)生(2014年12期)2014-11-12 13:12:52
