普通煙草Aux/IAA轉錄因子家族全基因組鑒定分析
2020-01-15 00:59:46王國平周東潔牛永志鄭昀曄
中國煙草學報 2019年6期
關鍵詞:煙草
王國平,周東潔,牛永志,鄭昀曄,2*
1 玉溪中煙種子有限責任公司,玉溪市南祥路14號 653100;
2 云南省煙草農業科學研究院,玉溪市南祥路14號 653100
生長素(auxin)是一類重要的植物激素,在細胞分裂與伸長、器官的發生與形態建成、頂端優勢、植物向性、逆境脅迫等諸多方面都起著非常重要的調控作用[1-3],還可以與油菜素內酯、赤霉素、乙烯等激素信號通路相互作用[4-6],影響植物種子萌發、植株生長、花果形成、植株衰老的整個生長發育過程。
生長素水平的時空動態變化可以精確而快速地激活下游一系列信號通路。其中生長素早期響應因子在信號傳導中起重要作用,如生長素/吲哚-3-乙酸(Aux/IAA)家族、生長素響應因子(ARF)家族、生長素上調小RNA(SAUR)、氨基環丙基-1-羧酸合成酶(ACS)、谷胱甘肽-S-轉移酶(GST)、生長素響應基因GH3[7-8]。Aux/IAA是一種生長素響應基因的轉錄抑制因子,在生長素濃度較低時,ARF-Aux/IAA異質二聚體的形成抑制了ARF的轉錄活性;當生長素濃度較高時,Aux/IAA發生降解,ARF從Aux/IAA中釋放,調控下游靶基因的表達[9]。典型的Aux/IAA包含四個保守的結構域(結構域I、II、III和IV),在許多預測的蛋白質中可能會缺少一個或幾個結構域[10]。結構域I包含保守的亮氨酸重復基序LxLxLx,抑制生長素下游調控基因。結構域II保守的GWPPV基序與F-box蛋白TIR1相互作用,導致Aux/IAA的不穩定性和快速降解[11]。位于C端的結構域III和IV能與其他Aux/IAA或ARF相互作用,形成同源二聚體和異源二聚體,從而調控下游基因表達[12]。
第一個被分離到的Aux/IAA基因是豌豆的PSIAA4/5和PS-IAA6基因[13]。隨著眾多植物基因組測序的完成,Aux/IAA基因家族已在30多種植物中得到鑒定分析[14],包括擬南芥29個[15]、油菜119個[16]、黃瓜27個[17]、大豆63個[18]、棉花44個[19]、番茄25個[20]、馬鈴薯26個[21]、玉米34個[22]、小麥84個[23]等。煙草作為模式植物,從中鑒定Aux/IAA家族基因對于研究生長素調控植物生長發育的機制具有重要作用。本研究利用生物信息學方法從煙草參考基因組中鑒定Aux/IAA家族基因,并對其序列特性、進化模式、表達模式和功能注釋等進行系統分析,為進一步解析生長素調控機制奠定基礎。
1 材料與方法
1.1 Aux/IAA基因家族成員鑒定及序列特征分析
最新的煙草基因組數據是由Edwards K D等人于2017年發布,測序品種為K326[24]。從茄科數據庫(https://solgenomics.net/)下載該基因組數據,下載的文件包括GFF文件、蛋白序列文件和核酸序列文件。從Pfam數據庫(http://pfam.xfam.org/)下載Aux/IAA基因家族的HMM模型文件(PF02309),利用HMMER3.0軟件進行Aux/IAA蛋白序列的初步檢索,E值設為1e-5。將初步檢索得到的序列上傳至pfam網站進行結構域預測,E值設為1e-5,剔除不含Aux/IAA蛋白典型結構域的序列,剩余的即為煙草Aux/IAA基因家族成員,同時根據Pfam結果統計各蛋白序列所包含的結構域類型和個數。利用ExPaSy網站的ProtParam工具(http://web.expasy.org/protparam/)分析蛋白的氨基酸含量、分子量、等電點、脂肪族氨基酸指數和疏水性指數等理化性質。
1.2 連鎖群定位和亞細胞定位
從GFF文件提取各蛋白序列的連鎖群位置信息。利用在線網站Softberry(http://linux1.softberry.com/)的ProtComp工具進行亞細胞定位預測。
1.3 系統發育樹構建
從Uniprot數據庫分別下載擬南芥的29個Aux/IAA家族成員[14]和番茄的25個Aux/IAA家族成員[19]。利用MEGA軟件進行多序列比對和進化樹構建,采用ClustalW比對方式,構建NJ樹,參數設置為:檢驗方法為Bootstrap法,檢驗次數設為1000次,gap處理選擇Pairwise Deletion方式。利用在線工具iTOL(https://itol.embl.de/)對進化樹進行可視化。
1.4 序列結構及蛋白結構域分析
將Aux/IAA家族成員的GFF文件提交到在線工具GSDS2.0(http://gsds.cbi.pku.edu.cn/)進行基因結構分析,并進行可視化。將Aux/IAA家族成員的蛋白序列文件提交到在線工具MEME(http://meme-suite.org/tools/meme)進行保守結構域的預測,參數設置為:Motif長度范圍為6~100,檢索的最大Motif數為10,其他選擇默認參數。利用在線工具EvolView(http://www.evolgenius.info/evolview/)對蛋白結構域進行可視化。利用NCBI的CDD工具(https://www.ncbi.nlm.nih.gov/cdd/?term=)鑒定結構域的類型。
1.5 表達模式分析
基因表達數據從EMBL-EBI網站下載,登錄號為 E-MTAB-176(http://www.ebi.ac.uk/arrayexpress/experiments/E-MTAB-176/)。煙草表達圖譜(TobEA)是由Edwards K D等人于2010年發布,該圖譜包含了早期衰老葉、中早期衰老葉、中后期衰老葉、后期衰老葉、成熟葉、莖生葉、幼葉、上部莖、下部莖、成熟根、幼根、種子、未開花蕾、開放花蕾、花朵、幼芽、莖頂端、子葉和花頂端19個樣品組織,貫穿從種子到衰老整個生命周期[25]。在SGN Unigenes數據庫,通過blastn比對檢索Aux/IAA家族成員匹配的Unigene ID,通過檢索得到的ID獲取對應的基因表達譜數據。將數據整理后利用Cluster3.0軟件進行聚類,利用Java TreeView工具可視化熱圖。
1.6 GO注釋分析
利用Blast2GO軟件對Aux/IAA蛋白序列進行功能注釋,利用在線工具WEGO2.0(http://wego.genomics.org.cn/)對結果進行可視化。
2 結果與分析
2.1 Aux/IAA家族成員鑒定及序列特征分析
基于隱馬爾科夫模型序列譜,從煙草基因組數據庫中共檢索得到84條候選基因序列,通過Pfam數據庫對候選序列進行結構域分析,剔除不含Aux/IAA同源異型結構域的序列,最終得到77個煙草Aux/IAA家族基因,按照NtIAA1~NtIAA77的順序對每個基因依次進行編號(見表1)。通過Pfam數據庫進行結構域分析結果顯示,在77個基因中,只包含Aux/IAA結構域的有56個基因;同時包含Aux/IAA和ARF結構域的有21個基因,這21個基因同樣也屬于ARF家族;同時包含Aux/IAA、ARF和B3結構域的有15個基因,這15個基因同樣也屬于B3超家族;分析Aux/IAA家族成員的理化性質發現,平均氨基酸長度為379,范圍在72~1104之間;平均分子量為42191.17 kDa,范圍在 8119.66~121828.27 之間;平均等電點為6.74,范圍在4.31~9.65之間,其中酸性蛋白(等電點小于7)有51個,堿性蛋白(等電點大于7)有26個;平均脂肪族氨基酸指數為71.95,范圍在59.5~92.08之間;平均疏水性指數為-0.559,范圍在-0.106~-1.244之間,該家族蛋白全部表現為親水性。
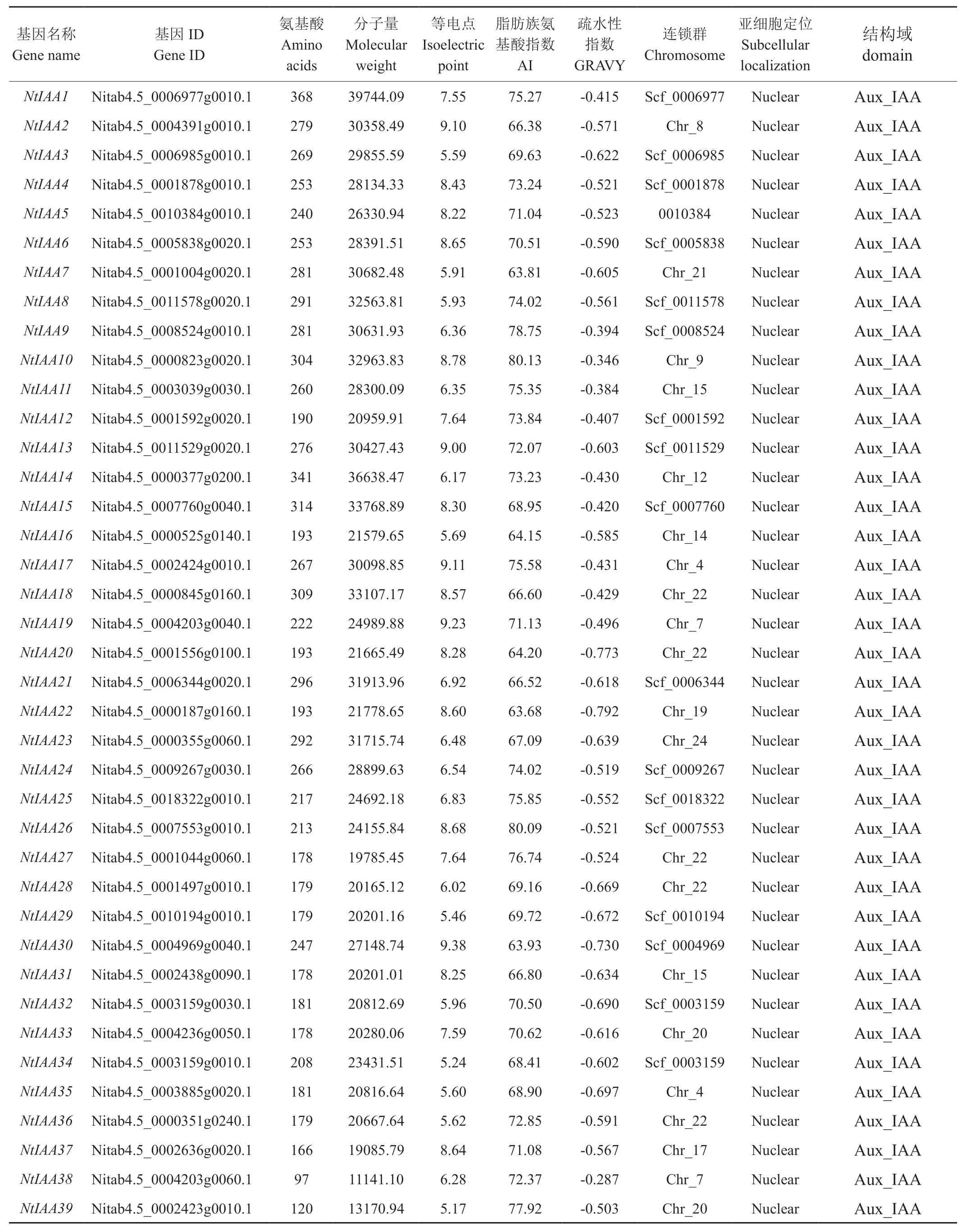
表1 Aux/IAA 家族蛋白特征Tab.1 Protein characteristics of Aux/IAA gene family

續表1
2.2 Aux/IAA家族連鎖群定位和亞細胞定位分析
連鎖群定位結果顯示(見表1),其中有46個基因定位到連鎖群上,有31個基因位于Scafford上。在定位到連鎖群上的46個Aux/IAA家族基因中,除了連鎖群2、3、6、11和16號外,其他連鎖群上均有分布,其中22號連鎖群上分布最多,達6個基因,9、17、19和20號連鎖群上各分布有4個基因,4和15號連鎖群各分布有3個基因,7、8、12、14、18、23號連鎖群上各分布有2個基因,1、5、10、21和24號連鎖群各分布有1個基因。在定位到Scafford上的31個Aux/IAA家族基因中,其中Scf_0007553和Scf_0003159上各分布有2個基因,其余Scafford上各分布有1個基因。亞細胞定位分析結果顯示(見表1),Aux/IAA家族所有基因都定位于細胞核內,這與以往研究結果一致[26]。
2.3 Aux/IAA家族進化分析
選取模式植物擬南芥的Aux/IAA基因29個,選取茄科作物番茄的Aux/IAA基因25個,同本研究獲得的77個煙草Aux/IAA基因一同構建系統進化樹。參照以往研究結果[27-28],將Aux/IAA家族分為A和B兩個類群(圖1),A類群又分為4個亞群(A1、A2、A3和A4),B類群又分為6個亞群(B1、B2、B3、B4、B5和B6)。A類群包括65個基因(煙草35個,擬南芥14個,番茄15個),B類群包括66個基因(煙草42個,擬南芥15個,番茄10個)。除了B6外,每個亞群中都分布有擬南芥、番茄和煙草的Aux/IAA基因,說明Aux/IAA分化時間早于物種分化時間。整個系統發育樹中共包括46對姐妹對,其中大部分為旁系同源蛋白,有40對(煙草30對,擬南芥9對,番茄1對),直系同源蛋白有6對(5對煙草-番茄,1對擬南芥-番茄),這可能是因為普通煙草是由絨毛煙草和林煙草經過染色體加倍進化而來,因此存在大量相似序列。另外,通過分枝的聚集也可以看出,煙草Aux/IAA基因與番茄更為接近,這是因為煙草和番茄同屬于茄科作物,親緣關系更近。令人驚奇的是,在B6亞群中,21個煙草Aux/IAA基因聚到了一起,而且與其他基因距離較遠,進一步對序列結構分析發現,這是因為這21個基因中,除了包含Aux/IAA結構域,還包含了ARF結構域,其中15個基因還包含B3結構域,這一點與其他作物差異較大。

圖1 Aux/IAA 家族系統進化樹Fig.1 Phylogenetic tree of Aux/IAA gene family
2.4 Aux/IAA家族序列結構及蛋白結構域分析
典型的Aux/IAA基因包含四個結構域(Ⅰ、Ⅱ、Ⅲ和Ⅳ)。本研究結構域分析結果顯示(圖2),所有煙草Aux/IAA基因都缺失了結構域Ⅰ,有40個基因同時包含了結構域Ⅱ(Motif3)、Ⅲ(Motif2)和Ⅳ(Motif1和Motif6),但其中有10個基因的結構域Ⅳ缺失了一部分(Motif6);有27個基因只包含結構域Ⅲ和Ⅳ,但其中有18個基因的結構域Ⅳ缺失了一部分(Motif6),這27個基因中的大部分屬于B6亞群成員(21個);基因NtIAA24、NtIAA30和NtIAA40只包含結構域Ⅱ和Ⅳ,但NtIAA40的結構域Ⅳ缺失了一部分(Motif6);基因NtIAA37和NtIAA41只包含結構域Ⅱ和Ⅲ;基因NtIAA53、NtIAA52、NtIAA45和NtIAA48分別只包含結構域Ⅱ、Ⅲ、Ⅳ;基因NtIAA74沒有檢測到任何結構域。綜合來看,大部分煙草Aux/IAA家族基因(有68個)都包含了結構域Ⅲ和Ⅳ,結構域Ⅲ和Ⅳ可與ARF相互作用,形成同源或異源二聚體,從而抑制生長素響應基因的表達,在功能行使中發揮重要作用。很明顯,B6亞群基因(21個)同時也包含了B3結構域(Motif4和Motif5)、ARF結構域(Motif7)以及類型未知的Motif8、Motif9和Motif10,B3基因家族是一個超基因家族,ARF基因家族也是一個生長素早期響應因子,Aux/IAA、B3和ARF家族成員之間通過互作,將眾多的信號通路聯系在一起。Aux/IAA內含子-外顯子結構分析顯示,除了B6亞群基因序列較長、外顯子數目較多外,其余基因序列相對較短,外顯子數目較少,結構較為簡單。
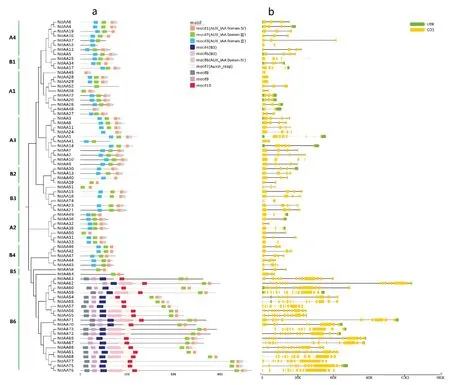
圖2 Aux/IAA家族蛋白結構域與序列結構Fig.2 Protein domain and sequence structure of Aux/IAA gene family
2.5 Aux/IAA家族表達模式分析
煙草Aux/IAA家族的77個成員中有50個基因,在煙草表達圖譜(TobEA)檢索到表達數據。基因聚類結果將這50個基因分為X和Y兩個類群,其中X類群又分為X1和X2兩個亞群,Y類群又分為Y1和Y2兩個亞群,X類群基因與系統發育樹中的A類群基因基本相同,Y類群基因與系統發育樹中的B類群基因基本相同,進一步說明同源性較高的基因之間有著相似的表達模式和生物學功能(圖3)。組織聚類結果將這19個組織分為兩個類群,其中一個類群包含根、莖、葉和種子等成熟的組織,另一個類群包含花蕾、子葉、幼葉、莖端等幼嫩的組織。
整體上看,不同亞群基因在不同的組織中的表達水平有所差異,同一亞群基因在不同的組織中的表達水平類似。X1亞群基因在不同發育時期的葉片中的表達量較高,X2亞群基因在花尖、幼芽、幼葉、未開放花蕾和開放花蕾等幼嫩組織中的表達量較高,Y1亞群基因在花朵和種子中的表達量較高,Y2亞群基因在根、莖和種子中的表達量較高。雖然不同Aux/IAA家族基因表達存在一定的組織特異性,但在植株生長發育的每個階段,都有Aux/IAA家族基因的參與。
2.6 Aux/IAA家族GO注釋分析
GO注釋結果顯示(圖4),Aux/IAA家族基因在細胞組成、分子功能和生物進程中都發揮了重要作用。在細胞組成方面,主要參與細胞、胞內和細胞器的組成,這關系到生長素可以直接作用于細胞膜或胞內組分,協同調節分裂、伸長和分化等胞內過程,而且作為非細胞自主信號,與其他信號轉導途徑相互作用,從而影響一些細胞反應;在分子功能方面,主要參與蛋白結合,這間接調控下游基因的表達,從而影響生長素對植物生長發育的調節;在生物進程方面,主要參與了胞內、代謝等生物進程調節,信號轉導,內源、胞內、化學等刺激反應,氮、細胞代謝過程等功能,這關系到生長素對植物向性生長、組織分化、器官發生、頂端優勢等的調節作用。
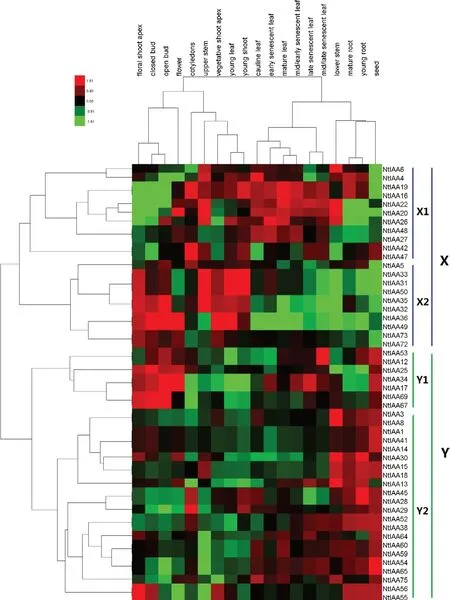
圖3 Aux/IAA家族基因表達熱圖Fig.3 Heatmap of gene expression of Aux/IAA gene family
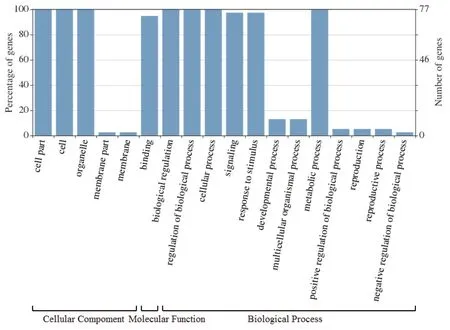
圖4 Aux/IAA家族GO注釋結果Fig.4 GO annotation results of Aux/IAA gene family
3 討論
隨著越來越多的植物基因組測序的完成,全基因組基因鑒定為基因功能研究提供了一種更高效的方法。Aux/IAA家族基因已在許多植物中得到鑒定[13],以往研究表明,很大一部分Aux/IAA家族基因是由基因復制事件(片段復制、串聯復制)進化產生的,而且Aux/IAA家族基因的復制事件似乎與全基因組復制(WGD)相一致。例如:大豆發生了兩次全基因組復制事件和一次全基因組三倍化事件,其中90%的Aux/IAA基因以片段復制的形式產生[18]。本研究共得77個普通煙草Aux/IAA家族基因,在已鑒定的30多種植物中,數量僅次于甘藍型油菜的119個[16]和普通小麥的84個[23]。而普通煙草是異源四倍體,普通小麥是異源六倍體,甘藍型春油菜是由自然種間雜交后雙二倍化進化而來的一種復合種,這三種作物都發生了基因擴張,可能是在進化過程中發生了全基因組復制使連鎖群加倍導致的,Aux/IAA基因擴張產生的功能冗余可能是為了適應復雜的自然環境。
Aux/IAA基因一般包括四個典型的結構域(I、II、III和IV),但在大部分植物中都存在一部分Aux/IAA基因缺失了其中某個或數個結構域[10]。例如:擬南芥中有11個(占比38%)[15],木瓜中有8個(占比44%)[29],馬鈴薯中有5個(占比15%)[21]等。在普通煙草中檢測到的77個Aux/IAA基因中,全部缺失了結構域I,有一半多(40個)包含了其余所有結構域,大部分(68個)都包含了結構域Ⅲ和Ⅳ。這些結構域的多樣性可能在生長素信號通路中行使不同的功能以適應復雜多變的自然環境。有趣的是,在B6亞群中的21個基因不僅含有Aux/IAA的結構域III和IV,也包含了ARF結構域,其中15個基因還包含了B3結構域。這些基因也屬于ARF基因家族或B3超基因家族。B3是一個廣泛存在于高等植物基因組中,并能與DNA結合的高度保守結構域,ARF基因家族也是一個生長素早期響應因子。以往研究表明,ARF基因的C端包含一個CTD結構域,該結構域與Aux/IAA的結構域III和IV高度相似。因此,Aux/IAA的結構域III和IV可以與ARF的CTD結構域相互作用,從而調控ARF蛋白的活性[30-31]。
一般認為普通煙草是由S基因組供體林煙草和T基因組供體絨毛煙草兩個祖先種,大約在20萬年前經過一次種間雜交產生[32],其存在兩套異源基因組。因此,本研究進化分析顯示,在鑒定的77個普通煙草Aux/IAA家族中存在大量的旁系同源基因。通過系統發育樹可以看出,煙草與番茄親緣關系更近,這是因為他們同屬于雙子葉綱茄科植物。以往研究表明,SiIAA3連接生長素和乙烯代謝途徑,RNA干擾該基因會導致幼苗葉片下垂和葉尖卷曲[33];RNA干擾SiIAA9該基因會導致葉片不規則和單性果實[34];抑制SiIAA15的活性會導致側根增多、腋芽異常發育、絨毛數量減少、坐果率降低、葉片厚度增大[35]。SiIAA17控制果實的重量[36];SiIAA27與葉綠素合成和獨腳金內酯合成有關[37-38]。SiIAA3與煙草的NtIAA20、NtIAA22、NtIAA27和NtIAA48聚到一起;SiIAA9與煙草的NtIAA1、NtIAA14和NtIAA41聚到一起;SiIAA15與煙草的NtIAA17和NtIAA34聚到一起;SiIAA17與煙草的NtIAA37和NtIAA53聚到一起;SiIAA27與煙草的NtIAA3和NtIAA8聚到一起。這些同源基因可能也具有類似的生物學功能。
Aux/IAA蛋白可以與ARF蛋白相互作用形成二聚體,從而影響生長素的調控作用。Aux/IAA蛋白是一種核蛋白,這與本研究的結果一致。理論上講ARF蛋白也屬于核蛋白,但是最新的研究發現大部分ARF蛋白被定位到細胞核,個別ARF蛋白卻被定位到葉綠體或線粒體[39]。這可能是由于ARF除了可以與Aux/IAA結合發揮作用,還有其他結合方式,例如:可以與含有TGTCTC AuxREs元件的生長素響應基因結合,可以通過它們的CTDs形成二聚體,再與靶基因結合等[40]。
Aux/IAA參與了胚胎發育、下胚軸生長、向性、花器官發育、側根伸長等生物過程,大部分擬南芥基因突變都會表現出相似的功能缺陷[41-42]。本研究中雖然不同Aux/IAA家族基因表達存在一定的組織特異性,但在植株生長發育的每個階段,都有Aux/IAA家族基因的參與。另外,Aux/IAA在植物抗逆性方面也具有很大作用。高粱在鹽和干旱脅迫下,SbIAA1受到誘導,其表達量在葉和根部組織中不同,該基因可能對高粱抗鹽抗干旱有重要作用[43]。本研究所分析的77種Aux/IAA蛋白均為親水性蛋白,蛋白質的高親水性使蛋白質能夠代替細胞中的水分子,從而很好的抵御干旱脅迫對植物造成的損傷[44]。
在過去十年里,在生長素感知和信號轉導機制方面取得了顯著成就。但在生長素信號通路中,Aux/IAA的介導途徑仍存在很多空白,Aux/IAA在植物抗逆方面的相關研究也相對較少。相信隨著基因組的不斷解析以及計算模型的不斷發展,越來越多的Aux/IAA信號網絡將被發現。煙草作為一種模式植物,通過研究煙草Aux/IAA基因,對于解析Aux/IAA功能及信號通路具有重要意義。
4 結論
本研究從普通煙草基因組中共鑒定了77個Aux/IAA家族基因,均定位于細胞核內,全部為親水性蛋白,大部分為酸性蛋白。Aux/IAA家族劃分為2個類群,10個亞群,Aux/IAA基因分化時間早于物種分化時間;煙草是異源四倍體,Aux/IAA家族存在大量旁系同源姐妹對(40對)。所有的煙草Aux/IAA基因均缺失了結構域Ⅰ,大部分基因都同時包含了結構域Ⅲ和Ⅳ;B6亞組基因同時包含了B3、ARF和Aux/IAA結構域,與其他基因差別較大。不同亞群的基因組織表達特異性不同,同一亞群的基因組織表達特異性一致。Aux/IAA家族基因主要參與細胞組成、蛋白結合、生物進程調節、信號轉導、刺激反應、代謝過程等功能。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
