飼料中蘇氨酸含量對建鯉肉質及組織蛋白酶B、L的影響
2020-01-13 01:43:32白稚子劉明宇李樹紅林昊林靈李冉陳治光陳秀華
食品與發酵工業 2019年22期
關鍵詞:水平
白稚子,劉明宇,李樹紅,林昊,林靈,李冉,陳治光,陳秀華
(四川農業大學 食品學院,四川 雅安,625014)
蘇氨酸作為動物生長所需的必需氨基酸,已被廣泛應用于畜禽飼料中,成為提高畜禽肉產率的關鍵營養素之一。大量研究也表明,適宜營養水平的蘇氨酸對促進陸生動物生長性能[1-2]、機體免疫[3-4]、腸道發育[5]及抗氧化特性[6]等方面發揮了重要作用。然而作為魚飼料中的限制性氨基酸,其對不同種經濟魚類的適宜水平目前研究尚不透徹,進行全面系統的探究十分有必要。
鯉魚是我國最重要的淡水經濟魚種之一,年養殖產量約為280萬t[7]。其中,建鯉(CyprinuscarpiovarJian)由于生長快、適應性強、抗病能力強且肉質鮮嫩,已成為我國鯉魚養殖的主導品種[8]。團隊前期針對飼料蘇氨酸水平對幼建鯉生長性能、蛋白質合成、消化吸收能力、免疫功能的影響作用進行了較系統的研究。設計7.4、9.1、12.2、15.7、18.6、22.3、25.2 g/kg 7個飼料蘇氨酸水平,發現相對于缺乏組和過量組,僅適宜水平(15.7 g/kg)蘇氨酸組,建鯉存活率、免疫性能、生長性能顯著增強,尤其是其蛋白質沉積率加快[9-11]。鑒于上述作用,推測飼料蘇氨酸水平可能與建鯉魚肉肉質特性也存在有一定的潛在聯系,然而關于飼料蘇氨酸水平對建鯉魚肉品質的影響尚未見報道。
肉質是一個綜合性狀,肉質好壞不僅與肌肉的物理、化學特性密切相關,某種程度上更是直接取決于肌纖維特性或者狀態[12]。原料魚肉硬度是影響魚肉片品質的重要因素。魚肉本身質地柔軟,并且大量研究表明魚類死后,溶酶體半胱氨酸組織蛋白酶B、L參與了肌纖維自溶,這些具有高內肽酶活性的酶類破壞死后魚肉肌肉肌球蛋白完整性,從而加速了宰后魚肉片的軟化[13],導致魚肉硬度品質劣變。
對于陸生動物,組織蛋白酶B可能與肉質,如嫩度[14-15]、背膘厚度[16]之間存在某些潛在關聯,被視為影響肉品品質的候選因子;值得注意的是,魚類營養攝入情況的改變或飼料中某些營養素成分和含量的改變,可進一步導致該類酶活或表達量的改變。飼料中脂肪酸的種類與含量,可影響大西洋鮭魚(Salmosalar)體內組織蛋白酶B的活性或其基因表達水平,同時使其魚肉質構發生改變[17]。由此推測,組織蛋白酶B、L作為魚肉品質的潛在影響因子,其可能受到營養水平的調節。
然而對飼料蘇氨酸水平對魚肉品質影響的研究尤其是與組織蛋白酶B、L的潛在關系未見報道,為此,本實驗在前期工作基礎上,以蘇氨酸缺乏組(7.4 g/kg)為對照,通過測定、比較及分析建鯉魚肉品質指標(肌纖維直徑、密度、肌纖維耐折力、肌原纖維斷裂指數、剪切力、肌肉pH值、失水率)及組織蛋白酶B、L活性和表達,初步探討了飼料中適宜水平(15.7 g/kg)和過量水平(25.2 g/kg)蘇氨酸對建鯉魚肉品質的影響及對組織蛋白酶B、L的影響,為進一步研究魚類蘇氨酸營養水平與魚肉品質關系及其可能的作用機制奠定實驗前提,并以期達到通過合理調節飼料蘇氨酸營養素水平進一步提高建鯉魚肉品質的應用目標。
1 材料與方法
1.1 材料與試劑
1.1.1 動物性實驗材料
選擇450尾健康幼魚((初始體重13.60±0.02)g),由四川農業大學動物營養所水生動物飼料與營養實驗室飼養并提供。飼養實驗設計蘇氨酸缺乏組(7.4 g/kg)、適宜組(15.7 g/kg)、過量組(25.2 g/kg)3個處理組,每組3個重復,每個重復50尾魚,每天飼喂6次,試驗期90 d。期間定期檢查水質,保證每天日光燈光照12 h,水溫控制在(25±10)℃,pH值保持在7.10±0.3。
采樣前進行24 h饑餓處理,分別從3個處理組中隨機選擇30尾魚,稱取體重。缺乏組平均體重為(61.22±3.86)g,適宜組平均體重為(100.10±10.37)g,過量組平均體重為(63.00±5.81)g。
1.1.2 建鯉組織蛋白酶B、L多克隆抗體及背肌石蠟組織切片
建鯉組織蛋白酶B、L多克隆抗體及其背肌石蠟組織切片,均由四川農業大學食品學院水產品加工與理論實驗室制備保存,其制備過程參見陳秀華[18]的方法。其中組織蛋白酶B多克隆抗體效價為1∶256 000,組織蛋白酶L多克隆抗體效價為1∶512 000。
1.1.3 主要試劑
Z-Arg-Arg-AMC/Z-Phe-Arg-AMC/8-苯胺基-1-奈磺(ANS)/三磷酸腺苷酸(ATP):美國Sigma公司;蛋白定量試劑盒:南京建成生物工程研究所;Ultra-Sensitive TM SP免疫組化試劑盒:福州邁新生物技術開發有限公司;DAB辣根過氧化物酶顯色試劑盒:上海碧云天生物技術有限公司。
1.2 儀器與設備
Varioskan全波長熒光酶標儀,賽默飛世爾科技公司;CX21光學顯微鏡,奧林巴斯;V-1100可見分光光度計,上海美譜達儀器有限公司;DY89-Ⅱ電動玻璃勻漿機,寧波新芝生物科技股份有限公司;Scientz-IID超聲波細胞粉碎機,蘇州江東精密儀器有限公司。
1.3 實驗方法
1.3.1 建鯉肌纖維特性測定
1.3.1.1 肌纖維直徑測定肌纖維直徑頻率分布圖
各組隨機取5尾建鯉,3%烏來糖麻醉并去皮,在脊椎骨上方、背鰭第一硬棘下方靠左位置取肉樣,每個樣品1cm3,入刀時使切面與肌纖維走向垂直,4%多聚甲醛固定4 h,58~60℃石蠟包埋,切片,脫蠟至水,HE染色[19]。
每張切片選取5個不重復視野,于(10×40)倍放大條件下完成圖像采集,Image pro-Plus 6.0軟件分析肌纖維直徑。測定方式參照JOHNSTON等[19]的方法。并且將測得各組肌纖維直徑按照0~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90 μm歸類,繪制肌纖維直徑頻率分布圖[20]。
1.3.1.2 肌纖維密度測定
采集處理及切片同1.3.4,對切片中單位面積(mm2)內的肌纖維進行計數,從而計算肌纖維密度。
1.3.2 建鯉魚肉品質指標測定
1.3.2.1 肌原纖維耐折力測定
參考鄭小淼等[21]的試驗方法。各組隨機取10尾建鯉,剪取背鰭第一硬棘下方、脊椎骨上方部位2.0 g背肌,加100 mL A液(KCl 14.9 g、EDTA-2Na 3.44 g、硼酸4.78 g,定容至2 L,用四硼酸鈉調解pH=7.0),22 000 r/min勻漿15 s,取樣于(10×40)倍顯微鏡下測量肌原纖維長度,每個樣品隨機記錄50個觀測數據,10次重復。根據肌原纖維長度判斷耐折力強弱,肌原纖維越長其耐折力越強,耐折力計算如公式(1)所示:
(1)
式中:Li表示每次測定肌原纖維長度,cm;A表示顯微鏡放大倍數為400;B表示為擴印倍數為3.7;n表示測定次數。
1.3.2.2 肌原纖維斷裂指數測定
斷裂指數測定參考HOPKINS等[22]的方法。各組隨機取10尾建鯉,剪取背鰭第一硬棘下方、脊椎骨上方部位1 g肉,加30 mL緩沖液(100 mmol/L KCl,20 mmol/L K2HPO4,1 mmol/L EDTA,1 mmol/L MgCl2,1 mmol/L疊氮鈉),均質機均質15 s后,用4層紗布過濾,10 mL緩沖液沖洗濾渣,收集濾液在2 ℃下離心(1 000×g,10 min),沉淀用10 mL緩沖液懸浮,再次離心,重復2次。最后將沉淀用10 mL緩沖液懸浮,稀釋20倍后,測定A540吸光值。實驗10次重復,取平均值后乘以200即為肌原纖維斷裂指數(myofibril fragmentation index,MFI)。
1.3.2.3 剪切力測定
各組隨機取10尾建鯉,順背部肌肉肌纖維方向,在脊椎骨上方、背鰭第一硬棘下方用鋒利的刀片等間隔剪取3個3 cm長厚度一致的肉樣,去除肌間刺,取樣時全程冰上操作。測定時,肉樣擺放方向應使剪切方向與肌纖維方向垂直,剪切力值用公斤力值(kg·f)表示,每尾魚3個肉樣,每個肉樣測定3次,取平均值。
1.3.2.4 pH值測定
取0.5 g建鯉背肌白肉,將肉樣充分剪碎研磨,放入勻漿管,加入10 mL蒸餾水,勻漿,4 ℃ 3 000 ×g離心10 min,取上清,用酸度計測pH值,重復3次,取平均值作為樣品pH值。
1.3.2.5 失水率測定
參考冷向軍[23]試驗方法。各組隨機取10尾建鯉,在背鰭第一硬棘下方、脊椎骨上方部位取5 g左右背肌,記錄數據為W0,置于沸水中煮沸2.5 min后,冷卻,用濾紙吸干魚肉表面水分,再次稱重記錄為Wt。10次重復,取失水率的平均值如公式(2)所示。
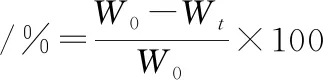
(2)
1.3.3 組織蛋白酶B、L活性測定
組織蛋白酶B、L活性的測定基本參照熒光合成肽底物法[24]。分別以熒光合成肽Z-Arg-Arg-AMC和Z-Phe-Arg-AMC為底物,用酶標儀于激發波長380 nm、發射波長460 nm下測定建鯉組織蛋白酶粗提液中CTS B、CTS L的活性。
一個酶活單位定義為在反應溫度(40℃)及pH值下,能夠在1 min內水解底物并釋放出1nmol AMC產物的酶活性量(1nmol AMC/min)。
1.3.4 組織蛋白酶B、L的免疫組化定量
在背鰭第一硬棘下方、脊椎骨上方靠左取1 cm×1 cm×1 cm魚背肌,58~60℃石蠟包埋,切片,脫蠟至水;選擇切片用乙醇、PBS洗脫后,滴加免疫組化試劑盒A液,PBS洗脫,滴加試劑盒B液孵育10 min后滴加1抗,37℃反應1 h,PBS洗脫后加入牛血清,室溫反應30 min,滴加試劑盒C液,室溫反應10 min,PBS洗脫,滴加試劑盒D液,室溫反應10 min后,PBS洗脫,除去PBS,滴加預先配制好的DAB顯色液(含85%超純水,5% DAB試劑盒A液,5% DAB試劑盒B液,5% DAB試劑盒C液),滴加過程中在顯微鏡下仔細觀察,陽性物為棕色。用Image pro-Plus 6.0軟件對所采集的圖片上組織染色部位進行面積測定和光密度測定,并計算單位面積內的平均光密度。
1.3.5 數據統計分析
采用SPSS 19.0數學軟件進行統計分析,計算各指標的平均值和標準差。采One-Way ANOVA法進行差異顯著性分析。P>0.05表示差異不顯著,0.01 用不同水平蘇氨酸添加的飼料飼喂建鯉90 d后,各組背肌HE染色結果見圖1。由圖1經形態分析可知,不同直徑的肌纖維組成一種馬賽克鑲嵌模式,大肌纖維中鑲嵌著小肌纖維,這些小肌纖維是新募集的肌纖維形成的。馬賽克增生使幼魚在生長期間肌纖維數量大量增加,這對商業水產養殖物種非常重要[21]。 飼料中添加不同水平蘇氨酸飼喂的建鯉肌纖維直徑、密度,經統計計算后列于表1。肌纖維直徑分布頻率如圖2所示。由表可知,適宜組建鯉平均肌纖維直徑最小,肌纖維密度最大,并分別顯著(0.01 ◆-大肌纖維;●-小肌纖維;A-缺乏組;B-適宜組;C-過兩組圖1 各處理組建鯉背肌的蘇木精-伊紅染色結果Fig.1 Hematoxylin-eosin staining results of the Jian carp dorsal muscles of each treatment group 表1 飼料中不同水平蘇氨酸對建鯉肌纖維特性的影響Table 1 Effects of different levels of threonine in feed on the characteristics of Jian carp muscle fibers 注:數據表示為平均值±標準差,同列數據肩標為不同小寫字母表示差異顯著(P<0.05),肩標為不同大寫字母表示差異極顯著(P<0.01)。下同。 圖2 肌纖維直徑頻率分布圖Fig.2 Muscle fiber diameter frequency distribution 飼料中蘇氨酸添加水平對建鯉魚肉品質的影響見表2。由表2可知,3個處理組建鯉肌肉的肌原纖維耐折力無顯著差異;適宜組建鯉肌肉肌原纖維斷裂指數極顯著(P<0.01)低于其他2組,剪切力值最高,并顯著(0.01 表2 飼料中不同水平蘇氨酸對建鯉魚肉品質指標的影響Table 2 Effects of different levels of threonine in feed on quality index of jian carp meat 飼料中不同水平蘇氨酸對建鯉肌肉中組織蛋白酶B(CTS B)、組織蛋白酶L(CTS L)的活性影響見圖3。由圖3可知,當飼料中蘇氨酸水平為7.4 g/kg(缺乏組)時,建鯉肌肉中CTS B、CTS L活性均顯著高于其他水平組(P<0.05),而適宜組與過量組之間CTS B、CTS L活性沒有顯著性差異(P>0.05),但適宜組稍低。 圖3 各處理組建鯉肌肉中的組織蛋白酶B、L活性Fig.3 CTS B and CTS L activities in Jian carp muscles of each treatment group注:缺乏組、適宜組、過量組進行方差分析,各數據柱狀圖上小寫和大寫字母分別代表0.05和0.01水平上的差異顯著性。 采用免疫組織化學方法,利用團隊前期制備的建鯉CTS B、CTS L多克隆抗體,對各處理組建鯉肌肉中CTS B、CTS L進行定位及相對定量。由圖4可知,經抗體標記、DAB顯色后的各處理組建鯉肌肉均呈現不同程度陽性染色,棕色染色部位即為真核表達的CTS B、CTS L。 進一步測定各組染色組織切片光密度,對建鯉肌肉中CTS B、CTS L蛋白表達水平進行相對定量,定量結果見圖5。由圖5可知,與CTS B、CTS L在肌肉中的活性模式相似,即當蘇氨酸水平7.4 g/kg (缺乏組)時,肌肉中CTS B、CTS L的蛋白表達量均顯著高于其他水平組(P<0.05),適宜組與過量組之間CTS B、CTS L活性均沒有顯著性差異(P>0.05)。 1~3-缺乏組、適宜組、過量組CTS B免疫組化染色;4~6-缺乏組、適宜組、過量組CTS L免疫組化染色圖4 各處理組建鯉背肌中組織蛋白酶B、L蛋白表達量的免疫組化測定Fig.4 Immunohistochemical determination of CTS B and CTS L protein expression in Jian carp dorsal muscle of each treatment group 圖5 各處理組建鯉肌肉中的組織蛋白酶B、L蛋白表達量Fig.5 Expression of CTS B and CTS L protein in Jian carp muscles of each treatment group 魚類肌肉生長方式與畜禽類不同,肌纖維募集(肌纖維數目增加)和肥大(肌纖維面積增大)是魚類肌纖維兩種主要的生長方式,可持續到魚類性成熟后,且在不同發育階段,募集與肥大的比重不同[25]。在幼魚期的肌纖維募集促成了肌纖維數量的顯著增加,這對于經濟魚種而言是十分重要的。也有研究表明,魚類直徑低于20 μm的肌纖維數可體現其肌纖維的募集能力,其比例越高說明其生長速度越快[26]。根據本研究得出的結果,與缺乏組、過量組相比,正常組建鯉肌肉不僅肌纖維平均直徑最小、肌纖維密度最高,且直徑小于20 μm的肌纖維數比例最高,說明在正常水平蘇氨酸的調節下,建鯉肌肉肌纖維募集能力最強,暗示其生長速度較快,生長能力提升。陳擁軍等[27]的研究表明,飼料中添加16.2 g/kg的蘇氨酸能有效提高草魚的生長能力,該最適蘇氨酸水平及其得出的研究結果與本文相似。 此外,肌纖維直徑分布和肌纖維密度還決定了肌纖維的多孔性,其與魚肉硬度質構特性密切相關,肌纖維密度越高,直徑越小,硬度越高[28]。在另一方面,肌纖維的物理特性(肌原纖維耐折力和肌原纖維斷裂指數)也可以間接反映肌肉的硬度品質,而剪切力直接反映了肌肉的硬度值,剪切力越大,硬度越強。研究表明肌原纖維耐折力與肌肉組織的剪切力成正相關[29],肌原纖維斷裂指數與剪切力成負相關[30],本研究中,經添加適宜水平蘇氨酸的飼料飼喂后,建鯉肌肉肌原纖維耐折力略有提高,肌原纖維斷裂指數則顯著降低,剪切力則顯著增加,說明肉質硬度提高。而適宜組的肌纖維密度、直徑數據也符合建鯉肌肉剪切力增加,硬度提高的實驗結果。綜上所述,飼料中適宜水平的蘇氨酸可改善建鯉魚肉的硬度品質。 魚肌肉中大量的水分與蛋白質的極性基團結合,在蛋白質的空間結構中形成水合離子,說明肌肉系水力與肌肉蛋白空間結構存在密切關系。研究表明,蘇氨酸能有效提高草魚蛋白質效率以及蛋白質沉積率,且當蘇氨酸水平(16.2 g/kg)達到最高,之后隨蘇氨酸水平升高而降低[27]。本研究中,當適宜組(蘇氨酸水平為15.7 g/kg)建鯉肌肉失水率最低,系水力最高。因此,推測蘇氨酸可能通過改善建鯉肌纖維生長、提高蛋白質沉積效率使肌肉系水力提高,對改善魚肉多汁性品質有一定的促進作用。 蘇氨酸缺乏將引起動物厭食,BAHUAUDA等[31]發現,大西洋鮭魚在宰殺前先饑餓4 d,導致其組織蛋白酶B、L基因表達量升高。長期處于饑餓狀態時,魚類通過提高蛋白水解酶表達量以水解蛋白維持其生理需求。研究表明蘇氨酸缺乏時,蛋白沉積率下降,蛋白合成速度下降[27],團隊前期研究也發現,蘇氨酸缺乏將導致建鯉蛋白沉積率下降[9-10]。本研究中,當建鯉處于蘇氨酸缺乏狀態時,可導致組織蛋白酶B、L活性及表達量均顯著升高,可能與此機制相關。研究表明組織蛋白酶B、L可水解肌原纖維蛋白[32],因此內源組織蛋白酶B、L也是引起其魚糜凝膠軟化的主要蛋白酶[33]。組織蛋白酶B、L活性更高的魚類,不僅死后魚肉蛋白的降解速度加快,高酶活亦影響魚肉片的硬度品質,同時還可能進一步影響加工制品如魚糜制品的質構品質。本研究中,相對于缺乏組、過量組,飼喂正常水平蘇氨酸的建鯉肌肉中組織蛋白酶B、L活性及蛋白表達量均降低,推測蘇氨酸可能通過某種途徑調控了組織蛋白酶B、L的活性及蛋白表達。這也進一步說明了正常水平的蘇氨酸對維持機體正常組織蛋白酶活性和表達水平起到促進作用,對保持魚肉質品質也具有積極的意義。由此可見,通過在飼料中添加營養物,對魚肌肉組織中組織蛋白酶的活性和表達量進行調節,從而調控魚肉肉質,對從飼養環節控制魚肉及加工制品品質,具有深遠的研究意義。 飼料中過量添加蘇氨酸對幼魚期建鯉肌纖維募集和魚肉品質的影響無顯著促進作用,而添加適宜水平蘇氨酸則能夠改善魚肉肌纖維特性及硬度品質,可能與其促進建鯉幼魚期肌纖維募集并降低內源組織蛋白酶B、L活性、蛋白表達量,具有潛在關聯性。2 結果與分析
2.1 飼料中蘇氨酸水平對建鯉肌纖維特性的影響



2.2 飼料中蘇氨酸水平對建鯉魚肉品質的影響

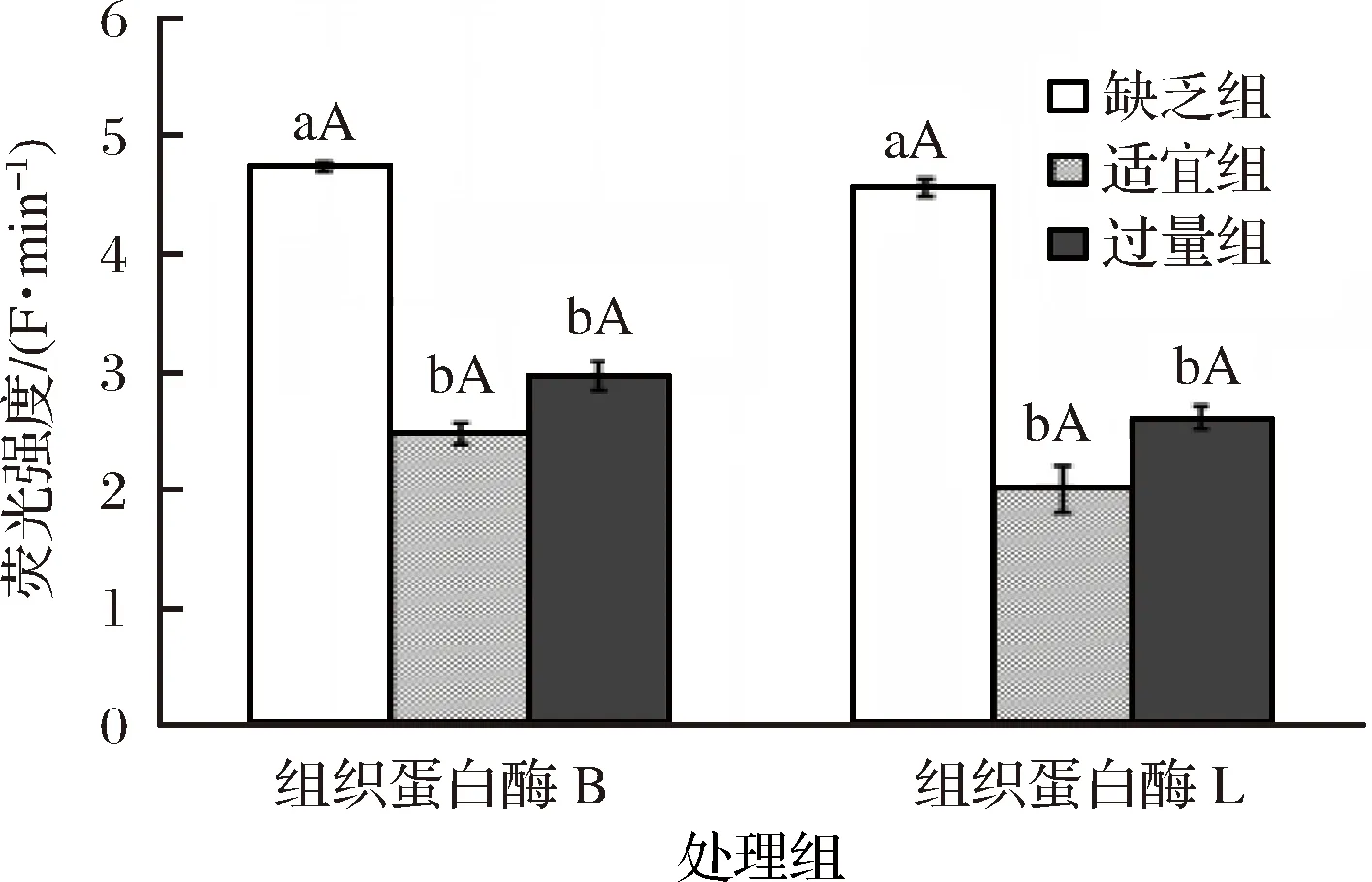
2.3 飼料中添加不同水平蘇氨酸對建鯉肌肉中CTS B/L活性的影響
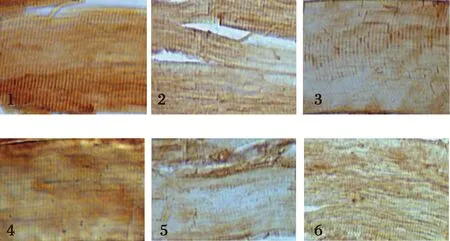
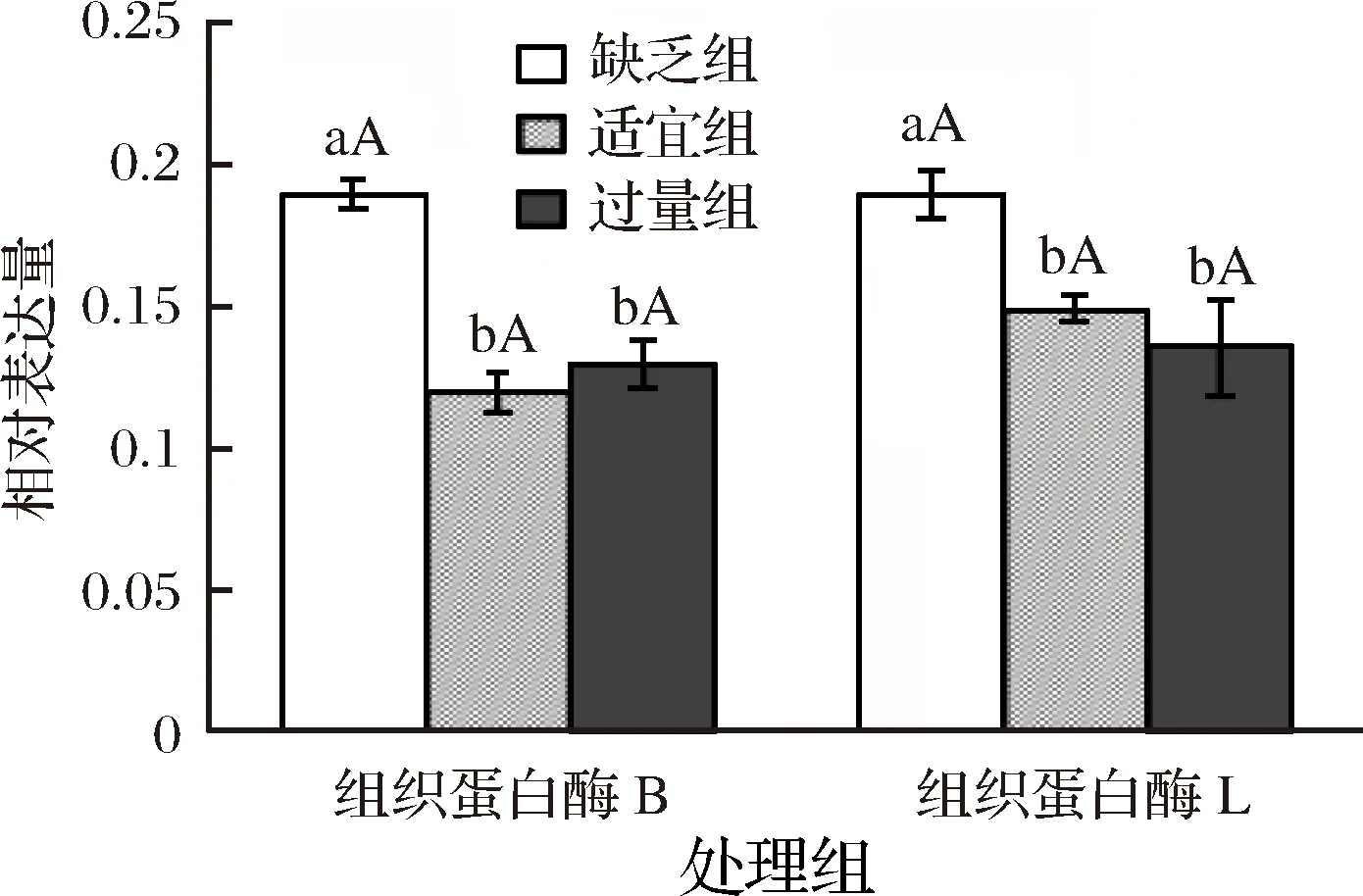
2.4 飼料中添加不同水平蘇氨酸對建鯉肌肉中CTS B/L蛋白表達量的影響
3 討論
4 結論
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15火花(2019年12期)2019-12-26 01:00:28人大建設(2019年6期)2019-10-08 08:55:48人大建設(2019年12期)2019-05-21 02:55:32雜文月刊(2018年21期)2019-01-05 05:55:28人大建設(2017年6期)2017-09-26 11:50:44學苑創造·A版(2015年11期)2016-01-14 09:03:27俄羅斯問題研究(2012年1期)2012-03-25 09:54:45中國火炬(2010年12期)2010-07-25 13:26:22中國火炬(2010年8期)2010-07-25 11:34:30
