CRTC2/miR-451軸調節非小細胞肺癌患者糖異生途徑的機制研究
2019-12-23 05:31:14石少卿
實用癌癥雜志 2019年12期
汪 矗 李 科 石少卿 李 敏
肺癌是世界上最常見的惡性腫瘤之一,其中約85%是非小細胞肺癌(NSCLC)[1]。NSCLC常診斷在患癌晚期,且患者的5年總生存率非常低[2]。癌細胞的葡萄糖代謝方式主要依靠糖酵解而不是三羧酸循環,由于癌細胞的快速生長會消耗大量的葡萄糖[3-5],因此機體會通過糖酵解的逆反應-糖異生途徑為癌細胞不斷地提供葡萄糖。
CRTC2是CREB調節轉錄的共激活因子,能夠促進肝臟的糖異生反應[6-7]。Shi等[8]發現CRTC2能夠促進肺癌細胞A549的體外遷移和增殖能力。Li等[9]發現非小細胞肺癌患者的CRTC2及其突變后的蛋白表達水平明顯高于正常人群。
本研究發現CRTC2在非小細胞肺癌患者癌組織中的表達水平明顯高于癌旁,且與糖異生關鍵酶G6Pase和PEPCK的表達顯著正相關,能夠增加NSCLC大鼠的血糖水平、促進糖異生反應并減少細胞凋亡。進一步的機制驗證發現miR-451是參與CRTC2促糖異生發生的關鍵因子,CRTC2通過CREB結合在miR-451的啟動子區并抑制miR-451的轉錄活性,提高miR-451靶基因Gyk及其下游G6Pase和PEPCK的表達水平,從而促進NSCLC的糖異生途徑,這為NSCLC的發生提供新的理論依據和作用機制。
1 材料與方法
1.1 材料
20只非小細胞肺癌大鼠模型購自廣州賽業生物公司;葡萄糖、丙酮酸和胰島素購自美國sigma公司;CRTC2蛋白、參與糖異生途徑的G6Pase 、PEPCK和Gyk蛋白及內參β-actin抗體均購自美國Abcam公司;干擾CRTC2的慢病毒載體、miR-451 mimic、miR-451 inhibitor及pcDNA3.1-CRTC2質粒均購自上海吉瑪公司;Tunel凋亡試劑盒、RNA提取及逆轉錄試劑盒和熒光素酶活性檢測試劑盒均購自美國promega公司。染色質免疫共沉淀試劑盒購自美國milippore公司。
1.2 實驗方法
1.2.1 標本采集 選取本院2015年至2018年收治住院的70例非小細胞肺癌患者,其中男性42例、女性28例;年齡40~70歲,中位年齡57歲;肺鱗癌30例、肺腺癌32例、腺鱗混合癌8例。納入標準:經病理學和細胞學證實為非小細胞肺癌;肝腎功能、血象正常。
1.2.2 非小細胞肺癌大鼠的處理 將0.2 ml滴度為1×109的陰性對照(sh-Ctr)、干擾CRTC2(sh-CRTC2)慢病毒載體及miR-451抑制劑通過尾靜脈注射至非小細胞肺癌大鼠,注射后第五天檢測糖異生途徑相關指標變化。
1.2.3 葡萄糖代謝檢測 葡萄糖代謝檢測包括葡萄糖耐量實驗(GTT)、丙酮酸耐受實驗(PTT)和胰島素耐受實驗(ITT)。對于GTT和PTT實驗,大鼠禁食6小時后腹腔注射2 g/kg體重的葡萄糖和丙酮酸;對于ITT實驗,大鼠禁食4 h后腹腔注射1單位/kg體重的胰島素,采用血糖測量儀檢測大鼠的血糖濃度。
1.2.4 實時定量PCR檢測 采用貼壁細胞RNA提取試劑盒提取非小細胞肺癌細胞株A549的總RNA。將提取的總RNA逆轉錄成cDNA后,熒光定量PCR檢測CRTC2、miR-451、G6Pase、PEPCK和Gyk的表達水平。各檢測基因的QPCR引物見表1。QPCR結果采用2-△△Ct法計算相對表達量。
1.2.5 western blot檢測 向A549細胞中加入RIpA蛋白裂解液,至4 ℃離心機以12 000 rpm離心10 min,吸取上清加入4倍體積的SDS loading buffer,混勻后放入100 ℃水浴5 min。 取30 μg的蛋白裂解液加入8% SDS-PAGE凝膠中分別以60 V和100 V電壓進行蛋白濃縮和分離,然后轉至PVDF膜,以10%脫脂奶粉室溫封閉2 h后,加入兔抗大鼠CRTC2、G6Pase、PEPCK和Gyk抗體,4 ℃過夜孵育后,ECL發光液顯色。
1.2.6 miR-451 mimic及pcDNA3.1-CRTC2轉染A549細胞 A549細胞接種至6孔板,當細胞密度達到80%,每孔轉染5微升的miR-451 mimic和陰性對照NC,轉染24 h后,再轉染入1 μg的pcDNA3.1-CRTC2質粒。轉染48 h后,檢測A549細胞中CRTC2、miR-451、G6Pase、PEPCK和Gyk糖異生途徑相關指標的變化。

表1 QPCR檢測引物
1.2.7 TUNEL染色檢測A549細胞凋亡 A549細胞經miR-451 mimic和pcDNA3.1-CRTC2質粒轉染48 h后,采用4%多聚甲醛固定。再利用TUNEL試劑盒檢測A549細胞的凋亡個數,定量結果采用TUNEL陽性細胞核占總細胞核的百分比表示。
1.2.8 熒光素酶報告載體的構建及轉染 采用MEME在線預測miR-451啟動子上游2000 bp的啟動子序列,發現-1121 bp處存在CREB的結合位點。從人基因組中釣取miR-451啟動子片段,并構建入pGL3-basic載體中。帶酶切位點的引物為Kpn I-miR-451-F:GGTACCTTTAGCTGAAACCGTTGA,Nhe I-miR-451-R:GCTAGCGTAGCGTTGTGATGGATC,構建成功后提取質粒,命名為pGL3-Wt。miR-451啟動子上CREB位點的缺失突變由上海生工公司合成,采用DNA合成技術合成不含有CREB結合位點的miR-451啟動子,并將此序列克隆至pGL3-basic載體,命名為pGL3-Mut。實驗組將pGL3-Wt或pGL3-Mut與pRL-TK載體共轉染入A549細胞中,對照組轉染pGL3-basic與pRL-TK,每孔轉染劑量為2 μg。轉染24 h后,裂解細胞并采用promega熒光素酶分析儀檢測螢火蟲熒光素酶和海腎熒光素酶的活性,兩者比值為熒光素酶的相對活性。
1.2.9 染色質免疫共沉淀 取出生長至90%以上的細胞,加入37%多聚甲醛,室溫交聯10 min后,加入甘氨酸終止交聯。將細胞培養基吸出,加入預冷的PBS潤洗兩次后,將細胞刮下并4 ℃下以2 000 rpm離2 min,收集細胞沉淀加入SDS Lysis Buffer后進行超聲破碎。將超聲后的細胞分為3組:input組、IgG陰性對照組和加anti-CRTC2抗體組。4 ℃過夜孵育后,于每組加入proteinA磁珠,孵育2 h后洗滌沉淀復合物。將洗滌后的DNA片段用于QPCR分析,設計miR-451啟動子-1121位點的QPCR引物,上游:5'-AGCATGTAGACTGCTGGGGCAA-3';下游:5'-CCTGCAGTAGGTTTCTGCTGCCTTG-3'。以input為內參,結果采用2-△△Ct法計算相對值。
1.3 統計學方法
采用Graphpadprism 6.05軟件統計分析實驗數據,單因素方差分析One-Way ANOVA和Student'st檢驗進行差異比較分析,P<0.05為差異有統計學意義。
2 結果
2.1 CRTC2、miR-451及糖異生途徑關鍵酶在非小細胞肺癌患者中的表達水平
CRTC2、G6Pase和PEPCK在NSCLC患者腫瘤組織中的表達水平明顯高于癌旁(P<0.01),與之相反,miR-451在NSCLC患者腫瘤組織中的表達水平明顯低于癌旁(P<0.01)。pearson相關性分析發現70例NSCLC患者的CRTC2與G6Pase和PEPCK的表達水平呈顯著正相關(P<0.01),相關系數分別為0.4815和0.2723;CRTC2與miR-451的表達水平則呈顯著負相關(P<0.01,γ=-0.608)。以上結果表明,糖異生途徑可能影響NSCLC的發生,CRTC2表達水平與糖異生關鍵酶G6Pase、PEPCK及miR-451存在明顯關聯。
2.2 CRTC2通過抑制miR-451表達影響NSCLC大鼠的葡萄糖代謝
設計和構建干擾CRTC2的慢病毒載體sh-CRTC2和miR-451 抑制劑,并通過尾靜脈注射NSCLC大鼠和葡萄糖耐量實驗(GTT)、丙酮酸耐受實驗(PTT)、胰島素耐受實驗(ITT)檢測CRTC2和miR-451是否參與NSCLC大鼠的葡萄糖代謝。如圖1A-1C所示,檢測結果發現三段CRTC2干擾片段中,sh-CRTC2-2的干擾效果最佳,且干擾CRTC2導致miR-451的表達水平顯著升高,而糖異生途徑關鍵酶G6Pase和PEPCK的表達明顯降低;注射miR-451 inhibitor挽救了被sh-CRTC2抑制的G6Pase和PEPCK表達水平,提示CRTC2上調糖異生途徑關鍵基因G6Pase和PEPCK的表達可能是通過抑制miR-451。與sh-Ctr對照組相比,干擾CRTC2導致NSCLC大鼠飼喂狀態和空腹狀態下的血糖水平顯著下降,而miR-451 inhibitor的注射緩解了血糖的下降程度,見圖1D。PTT實驗表明sh-CRTC2的NSCLC大鼠新生肝糖原水平低于sh-Ctr組,而miR-451 inhibitor使肝糖原生成水平明顯上升;ITT實驗顯示干擾CRTC2的表達導致血糖水平降低,而注射miR-451 inhibitor緩解了血糖下降趨勢,提示NSCLC大鼠對胰島素的敏感性與CRTC2和miR-451的表達有關,兩者分別發揮著促進和抑制的作用;GTT實驗表明干擾CRTC2導致外源葡萄糖的消耗更加迅速,明顯快于sh-Ctr組大鼠,因此導致血糖水平降低的更快,注射miR-451 inhibitor挽救了血糖水平的降低趨勢,使其回升的血糖水平明顯高于sh-CRTC2組,見圖1E-1G。
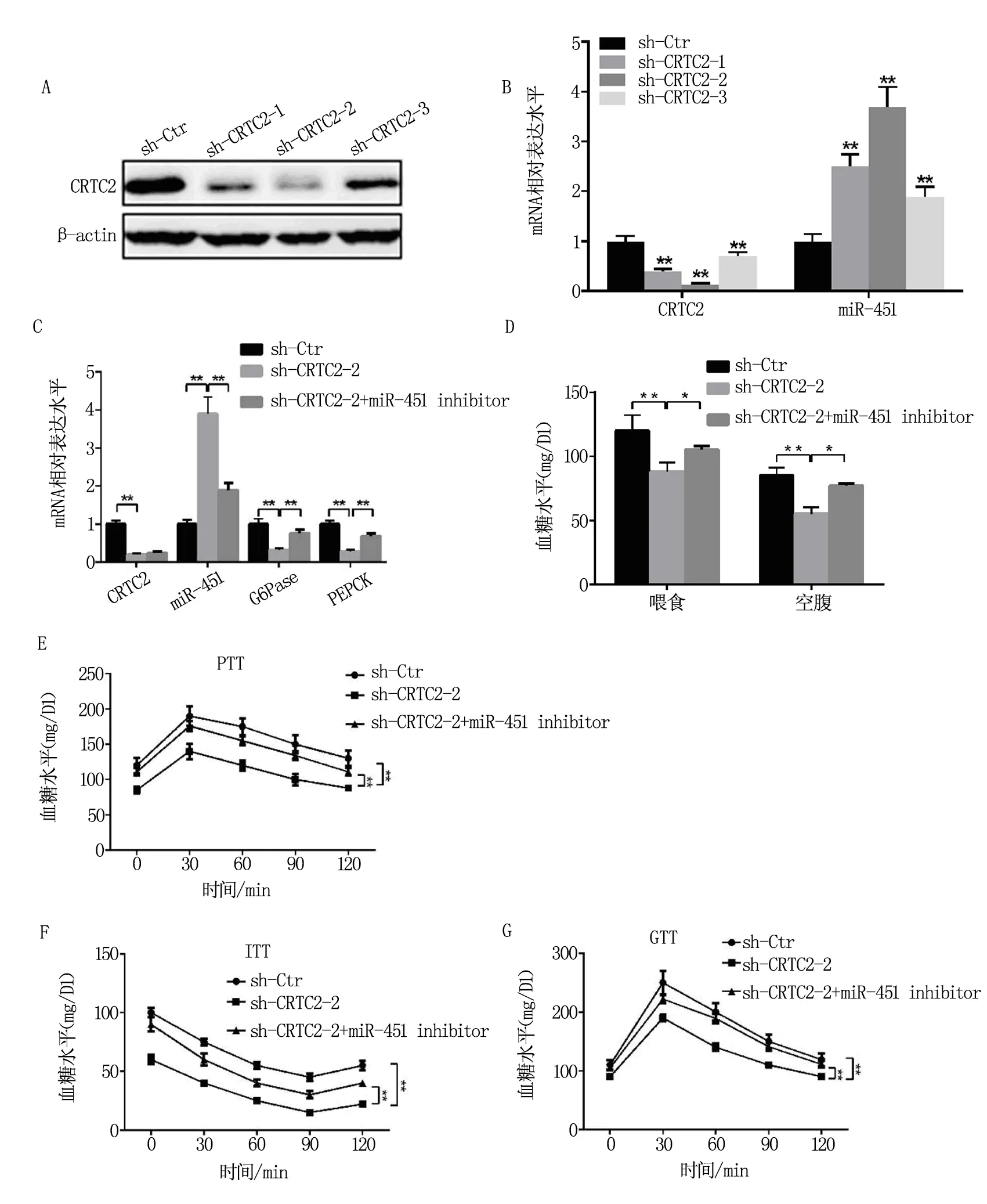
注:圖1A-1B表示CRTC2三段干擾序列在NSCLC肺癌組織中的干擾效果;圖1C表示CRTC2和miR-451 inhibitor對G6Pase和PEPCK RNA水平的影響;圖1D表示空腹和喂食狀態的NSCLC大鼠的血糖水平受到CRTC2干擾和miR-451 inhibitor的影響;圖1E-1G分別表示CRTC2干擾和miR-451 inhibitor影響PTT、ITT和GTT實驗下的NSCLC大鼠血糖水平。
2.3 CRTC2/miR-451軸通過促進糖異生途徑抑制細胞凋亡
本研究采用Tunel凋亡實驗檢測NSCLC細胞A549在經受miR-451 mimic和pcDNA3.1-CRTC2轉染后的凋亡情況,結果發現相比NC對照轉染組,轉染miR-451 mimic促進A549細胞的凋亡,而Gyk、PERCK和G6Pase的表達受到抑制;在轉染miR-451 mimic的基礎上再次轉染pcDNA3.1-CRTC2,結果發現與單獨轉染miR-451 mimic相比,A549的細胞凋亡個數減少,且受miR-451抑制的Gyk、PERCK和G6Pase表達水平明顯回升,見圖2A-2D,以上結果證明miR-451能夠抑制糖異生途徑并促進細胞凋亡,而CRTC2減弱了miR-451對糖異生的抑制作用,及其引起的A549細胞凋亡。
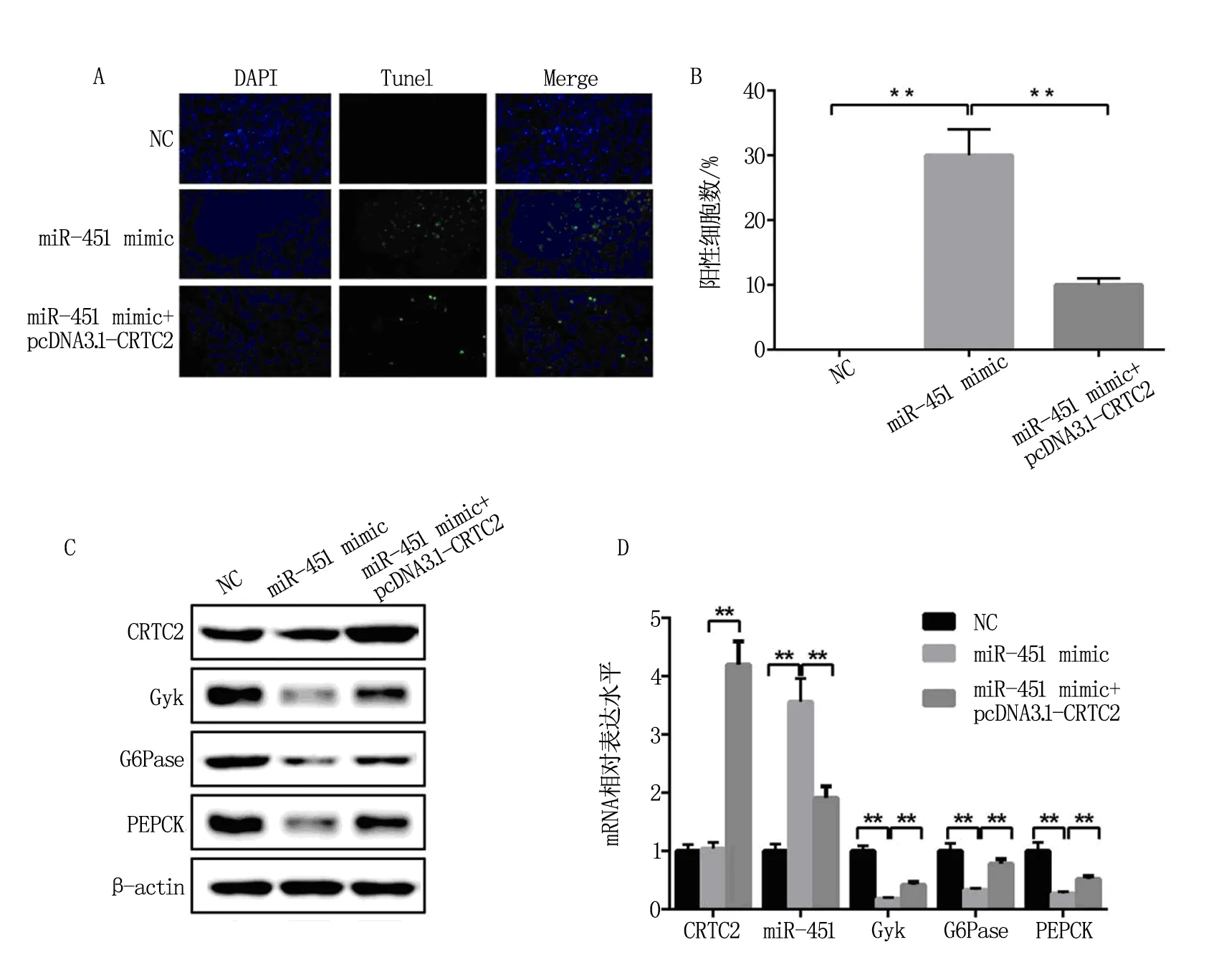
A和B表示A549細胞凋亡實驗的檢測結果;C和D表示western blot和QPCR檢測miR-451 mimic和pcDNA3.1-CRTC2轉染的A549細胞中miR-451靶基因Gyk及下游PERCK和G6Pase的表達水平。
2.4 CRTC2抑制miR-451的分子機制
我們采用hTTP://meme-suite.org/數據庫在線預測miR-451上游2 000 bp啟動子序列,發現-1121 bp處存在CREB的結合序列。染色質免疫共沉淀(ChIP)實驗發現,干擾CRTC2導致CREB與miR-451啟動子-1121 bp處的結合能力明顯下降,見圖3。將大小為2000 bp的miR-451啟動子克隆至pGL3-basic載體,重組載體命名為pGL3-Wt,CREB位點缺失突變的miR-451啟動子重組載體命名為pGL3-Mut。熒光素酶報告系統結果顯示pGL3-Mut的雙熒光素酶活性明顯高于pGL3-Wt,而干擾CRTC2導致pGL3-Wt和pGL3-Mut的雙熒光素酶活性明顯高于sh-Ctr對照組,表明miR-451啟動子-1121 bp處的CREB結合序列是CRTC2抑制miR-451轉錄活性的關鍵位點,突變此位點序列或干擾CRTC2都能夠增強miR-451啟動子的轉錄活性。
3 討論
腫瘤細胞的代謝重編程,被認為是癌癥發生的標志之一,在過去的十年中,與腫瘤發生、發展和轉移相關的代謝途徑逐漸被報道,如戊糖磷酸途徑、絲氨酸甘氨酸途徑和糖酵解途徑[10-12],但與糖酵解相逆的糖異生途徑卻鮮有報道。糖異生途徑是機體中的非碳水化合物碳源如乳酸、丙酮酸、氨基酸及甘油等物質轉變為糖(葡萄糖或糖原)的過程,在空腹或饑餓狀態下,機體主要依靠糖異生途徑維持正常血糖濃度。這致使糖酵解或糖異生反應會不可避免地造成細胞的代謝壓力,進而導致癌細胞的代謝重編程受損[13-16]。因此,糖酵解或糖異生途徑作為代謝重編程的治療靶點對癌細胞的生長和生存至關重要。
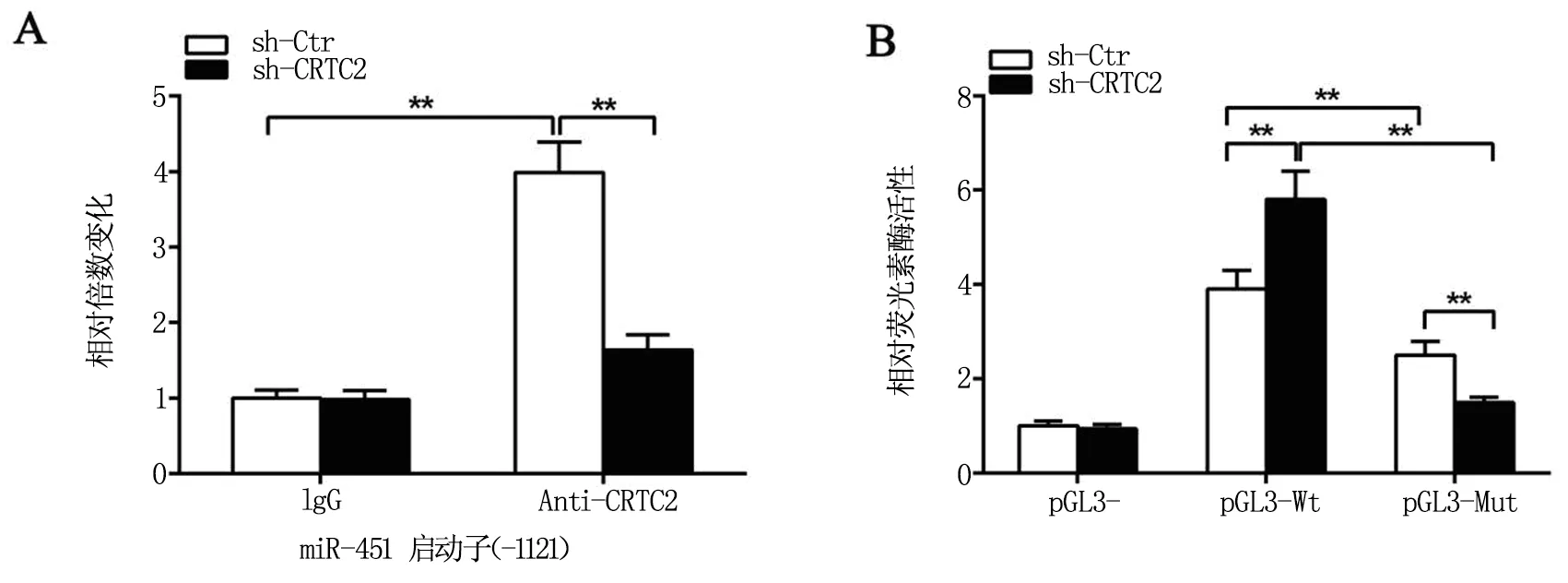
A表示ChIP實驗驗證-1121位置的結合位點與CRTC2和CREB轉錄復合物的結合能力;B表示熒光素酶實驗驗證干擾CRTC2對miR-451野生型及突變型的啟動子活性的影響。
CRTC2作為人體血糖調節的開關性蛋白,在正常情況下,以磷酸化形式存在于細胞質中,當接收活化信號,CRTC2迅速去磷酸化并轉位至細胞核中與CREB蛋白形成轉錄復合物,激活糖異生關鍵酶G6Pase、PEPCK的表達,從而調節糖異生途徑。CRTC2與癌癥發生的相關報道較少,Fang等[17]的研究發現CRTC2是淋巴瘤抑制因子,能通過轉錄DNA得錯配修復來保持基因組的完整性;Brown等[18]發現CRTC2通過調節芳香化酶,影響更年期和肥胖相關得荷爾蒙環境,并在肥胖和乳腺癌之間建立聯系。此外,Li等發現CRTC2及其突變體在非小細胞肺癌組織中的表達水平高于癌旁組織。以上報道提示CRTC2作為調節糖異生途徑的關鍵因子亦可能參與癌癥的發生。本研究通過對70例非小細胞肺癌患者癌和癌旁組織的定量分析,發現CRTC2在NSCLC癌組織中的表達明顯高于高旁,這與Li的報道一致。不僅如此,CRTC2與糖異生關鍵酶G6Pase和PEPCK的在腫瘤組織中的表達均高于癌旁,且與CRTC2呈顯著正相關,表明CRTC2在非小細胞肺癌腫瘤組織中的表達水平可能影響糖異生途徑。
腫瘤細胞中的miR-451處于失調狀態,能通過靶向抑制多個分子發揮促癌或抑癌的作用。研究發現miR-451在非小細胞肺癌組織中的表達降低,而上調miR-451能夠增強非小細胞肺癌對放療和順鉑化療的敏感性[19-20],表明miR-451對NSCLC的發展發揮著抑制作用。而在高脂飲食誘導的糖尿病小鼠和肥胖小鼠肝臟中,miR-451表達上升,表明其參與肝臟的代謝紊亂[21-22]。Shu等[23]發現miR-451是糖異生反應的上游抑制因子,能夠靶向抑制Gyk的表達從而進一步阻遏AKT-FOXO1-PEPCK/G6Pase 通路。本研究發現miR-451在NSCLC癌組織中的表達明顯高于癌旁組織,且與CRTC2的表達呈顯著負相關,推測miR-451可能與CRTC2共同參與NSCLC的發生。為證實此猜想,進一步的研究利用干擾CRTC2的慢病毒載體和miR-451抑制劑通過尾靜脈注射至NSCLC大鼠模型中,通過QPCR和葡萄糖耐量實驗(GTT)、丙酮酸耐受實驗(PTT)和胰島素耐受實驗(ITT)證實缺乏CRTC2導致miR-451表達上升,且降低空腹和飽腹狀態下NSCLC大鼠的血糖水平,而miR-451 inhibitor在降低miR-451表達的同時,亦挽回了CRTC2缺陷大鼠的血糖下降趨勢,使其血糖水平高于僅注射sh-CRTC2的NSCLC大鼠,表明CRTC2/miR-451軸具有穩定NSCLC大鼠血糖水平的功能。
已有的研究證實敲除肺癌細胞中的PERCK能夠增強葡萄糖耗竭誘導的細胞凋亡,miR-451能夠靶向調節Gyk控制PERCK的表達[23],提示miR-451不僅能夠抑制糖異生途徑,還能通過下調PERCK引起細胞凋亡。為證實CRTC2通過糖異生途徑影響非小細胞肺癌發生的進程,本研究通過向A549細胞轉染miR-451 mimic、過表達CRTC2(pcDNA3.1-CRTC2)的載體以及細胞凋亡實驗,發現miR-mimic導致A549凋亡細胞增加,而lv-CRTC2則抑制了miR-mimic的促凋亡作用。進一步,通過western blot和QPCR發現CRTC2阻遏了miR-451對Gyk的靶向抑制,表現為Gyk及其下游的G6Pase和PEPCK表達升高,由于PEKCK參與葡萄糖消耗引起的細胞凋亡,因此本研究結果表明CRTC2通過抑制miR-451的表達促進糖異生途徑,并參與NSCLC細胞的凋亡抑制。
CRTC2作為CREB的共激活因子共同參與調節細胞的內質網應激和糖異生途徑[24]。本研究采用在線分析hTTP://meme-suite.org/預測miR-451上游啟動子的-1121bp處存在CREB的結合序列。進一步,將miR-451啟動子及其CREB位點突變序列克隆至熒光素酶報告載體,并采用ChIP檢測CRTC2與CREB的轉錄復合物與-1121bp位點的結合能力和miR-451啟動子的轉錄活性,結果發現干擾CRTC2明顯阻遏了其與miR-451啟動子的結合能力,并導致啟動子的轉錄活性升高,表明-1121bp位點對miR-451啟動子的抑制作用;進一步的點突變實驗結果發現野生型的miR-451啟動子活性明顯低于突變型,而在CRTC2干擾的A549細胞中,突變型啟動子的活性升高的更為明顯,表明-1121bp處的CREB結合序列是抑制miR-451啟動子活性的關鍵位點,CRTC2通過此位點抑制miR-451的轉錄,下調其在NSCLC細胞中的表達。
綜上所述,CRTC2是參與非小細胞肺癌糖異生反應的上游關鍵因子,通過轉錄抑制miR-451,增強miR-451下游Gyk、G6Pase和PEPCK的表達,并進一步減少NSCLC細胞的凋亡,這為NSCLC的發生提供新的理論依據和作用機制。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
保健醫苑(2022年6期)2022-07-08 01:26:34
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
媽媽寶寶(2017年3期)2017-02-21 01:22:30
飼料與畜牧(規模養豬)(2016年5期)2016-12-01 03:48:40
醫學研究雜志(2015年12期)2015-06-10 06:57:46
