5-氟尿嘧啶對(duì)結(jié)腸癌干細(xì)胞干性和增殖的影響及相關(guān)機(jī)制
2019-12-20 09:17:32蔡尚黨蔡滿地婁寧郭艷萍殷濤
中國(guó)老年學(xué)雜志 2019年24期
蔡尚黨 蔡滿地 婁寧 郭艷萍 殷濤
(1河南省中醫(yī)院肛腸科,河南 鄭州 450002;2河南大學(xué)醫(yī)學(xué)院臨床醫(yī)學(xué)系;3中山大學(xué)附屬腫瘤醫(yī)院ICU;4河南省人民醫(yī)院病理科;5華中科技大學(xué)同濟(jì)醫(yī)學(xué)院附屬協(xié)和醫(yī)院胰腺病研究所)
結(jié)腸癌是世界范圍內(nèi)第三常見(jiàn)的癌癥,每年造成約70萬(wàn)人死亡〔1~3〕。在過(guò)去的二十年中,該病在發(fā)展中國(guó)家的發(fā)病率增加了2~4倍〔4,5〕,手術(shù)切除和輔助化療是治療結(jié)腸癌的主要方法,然而原發(fā)性或獲得性藥物抗性是該病成功治愈的主要挑戰(zhàn)〔6〕。因此,亟待開(kāi)發(fā)新的治療策略或新的輔助藥物以增強(qiáng)該病的治療效果。5-氟尿嘧啶(FU)是一種細(xì)胞生長(zhǎng)抑制劑,是最常用的治療結(jié)腸癌的藥物,然而,對(duì)晚期結(jié)腸癌5-FU化療的有效率僅有20%左右,5-FU和如伊立替康及奧沙利鉑等新的化療藥物的聯(lián)合使用提高了5-FU對(duì)結(jié)腸癌的治療效果〔7〕,在5-FU的代謝途徑中,包括胸苷酸合成酶、甲基四氫葉酸還原酶、二氫嘧啶脫氫酶和胸苷磷酸化酶在內(nèi)的四種酶被認(rèn)為與5-FU的治療敏感性和抗性密切相關(guān),然而,許多研究結(jié)果相互矛盾〔8〕。最近,胰島素樣生長(zhǎng)因子信號(hào)通路被證明對(duì)于結(jié)腸癌的發(fā)育至關(guān)重要,其能通過(guò)激活如PI3K/Akt等存活通路而有利于結(jié)腸癌細(xì)胞的存活、侵染、轉(zhuǎn)移和化療抗性〔9~12〕。本研究探討5-FU對(duì)結(jié)腸癌干細(xì)胞(CCSCs)干性和增殖的影響及可能機(jī)制。
1 材料與方法
1.1CCSCs 標(biāo)記CD133、CD44和ALDH1b1陽(yáng)性的CCSCs購(gòu)自美國(guó)Celprogen公司。為維持上述細(xì)胞的未分化狀態(tài),培養(yǎng)過(guò)程中使用CCSCs生長(zhǎng)培養(yǎng)基和特異性包被的細(xì)胞培養(yǎng)皿,細(xì)胞培養(yǎng)于37℃,5%CO2濕潤(rùn)培養(yǎng)箱中,當(dāng)細(xì)胞達(dá)到80%匯合度時(shí),用于實(shí)驗(yàn)。所有實(shí)驗(yàn)所用細(xì)胞代次低于10代。細(xì)胞分為陰性對(duì)照組、DMSO組、舒林酸組、5-FU組。
1.2主要儀器及試劑 5-FU(Sigma,美國(guó)),兔抗人Bax、Bcl-2、cyclinD1和c-Myc抗體,鼠抗人α-tubulin抗體(Sigma,美國(guó)),兔抗人細(xì)胞色素c抗體(Sigma,美國(guó)),兔抗人Histone抗體(Cell Signaling Technology,美國(guó)),辣根過(guò)氧化物酶(HRP)標(biāo)記羊抗兔/鼠IgG(Abcam,美國(guó)),嘌呤霉素(Sigma,美國(guó)),p53特異性shRNA質(zhì)粒(Santa Cruz,美國(guó)),5-溴-2'-脫氧尿苷(BrdU)檢測(cè)試劑盒(Cell Signaling Technology,美國(guó)),細(xì)胞死亡檢測(cè)試劑盒(Roche,德國(guó)),電化學(xué)發(fā)光(ECL)顯色試劑盒(碧云天生物技術(shù)研究所,中國(guó)),CO2培養(yǎng)箱(SANYO,日本),相差顯微鏡(Olympus,日本)。
1.3介導(dǎo)p53敲低表達(dá)的重組慢病毒短發(fā)夾RNA(shRNA)載體 CCSCs接種含靶向p53基因特異性shRNA的慢病毒顆粒,在含有5 μg/ml凝聚胺的CCSC培養(yǎng)基中培養(yǎng)的CCSCs接種10 MOI的慢病毒,慢病毒感染后24 h,更換新鮮培養(yǎng)基,細(xì)胞繼續(xù)培養(yǎng)2 d后,在含有7.5 μg/ml嘌呤霉素的選擇性培養(yǎng)基中加壓篩選5 d。
1.4細(xì)胞增殖檢測(cè) BrdU檢測(cè)試劑盒檢測(cè)細(xì)胞活性,正常CCSCs或shRNA-p53 CSCs以1×105個(gè)/孔的密度鋪板于12孔板中,培養(yǎng)24 h后,更換為不含血清的CCSCs培養(yǎng)基,并經(jīng)10 μg/ml舒林酸或2.5 μg/ml 5-FU處理,處理24 h后,檢測(cè)BrdU含量,實(shí)驗(yàn)重復(fù)3次,結(jié)果以均值表示。
1.5末端轉(zhuǎn)移酶dUTP切口末端標(biāo)記檢測(cè) 使用熒光標(biāo)記的核苷酸和末端脫氧核苷酸轉(zhuǎn)移酶(TdT)鑒定細(xì)胞凋亡產(chǎn)生的DNA片段。6×104個(gè)正常CCSCs或shRNA-p53-CCSCs接種于玻璃載玻片上,經(jīng)10 μg/ml舒林酸或2.5 μg/ml 5-FU處理12 h后,根據(jù)說(shuō)明書(shū)的要求,使用原位細(xì)胞死亡檢測(cè)試劑盒定量檢測(cè)細(xì)胞凋亡情況,同時(shí)用不含TdT的孵育液孵育的樣品作為陰性對(duì)照,計(jì)數(shù)5個(gè)視野內(nèi)的染色細(xì)胞數(shù)目,計(jì)算凋亡細(xì)胞比例,每次至少計(jì)數(shù)50個(gè)細(xì)胞,實(shí)驗(yàn)重復(fù)3次,結(jié)果以均值表示。
1.6球體形成實(shí)驗(yàn) 1×104個(gè)CCSCs用干細(xì)胞特異性的不含血清的培養(yǎng)基培養(yǎng)于超低吸附性的6孔培養(yǎng)板中,然后將細(xì)胞置于正常細(xì)胞培養(yǎng)箱中,細(xì)胞接種于6孔板后6 h,加入10 μg/ml舒林酸或2.5 μg/ml 5-FU,處理10 d后,使用相差顯微鏡計(jì)數(shù)球體數(shù)量,實(shí)驗(yàn)重復(fù)3次,結(jié)果以均值表示。
1.7Western印跡檢測(cè) 正常CCSCs或shRNA-p53-CCSCs以3.0×105個(gè)/孔的密度鋪于6孔板中,用CCSCs培養(yǎng)基培養(yǎng),24 h后,細(xì)胞轉(zhuǎn)移至無(wú)血清培養(yǎng)基中孵育18 h,提取細(xì)胞樣品的總蛋白,并將總蛋白轉(zhuǎn)移至PVDF膜上,孵育一抗,一抗稀釋比例為:兔抗人Bax、Bcl-2、cyclinD1和c-Myc抗體1∶1 000稀釋,鼠抗人α-tubulin抗體1∶5 000稀釋,兔抗人細(xì)胞色素c抗體1∶1 000稀釋,兔抗人Histone抗體1∶2 000稀釋。一抗4℃孵育過(guò)夜,然后,HRP標(biāo)記的羊抗兔/鼠二抗以1∶2 000的比例稀釋,室溫孵育2 h,目的蛋白用增強(qiáng)型ECL化學(xué)發(fā)光液顯色,并經(jīng)BandScan軟件進(jìn)行灰度掃描分析,α-tubulin作為總蛋白檢測(cè)時(shí)上樣的內(nèi)參蛋白,而Histone作為細(xì)胞核蛋白檢測(cè)時(shí)上樣的內(nèi)參蛋白。
1.8統(tǒng)計(jì)學(xué)方法 采用GraphPad Prism5軟件進(jìn)行t檢驗(yàn)或單因素方差分析。
2 結(jié) 果
2.15-FU抑制CCSCs增殖并誘導(dǎo)凋亡 對(duì)陰性照組相比,5-FU的濃度為2.5 μg/ml時(shí),正常CCSCs的增殖速率下降了52%,而shRNA-p53-CCSCs增殖速率下降了73%,10 μg/ml的舒林酸處理導(dǎo)致正常CCSCs增殖速率下降了47%,然而,敲低CCSCs中p53表達(dá)后,舒林酸對(duì)CCSCs增殖的抑制效應(yīng)較未敲低時(shí)強(qiáng)(51%),表明舒林酸對(duì)CCSCs的增殖抑制效應(yīng)具有p53依賴性。使用TUNEL法檢測(cè)細(xì)胞凋亡情況結(jié)果表明,5-FU處理后,正常CCSCs凋亡率為31%,shRNA-p53-CCSCs凋亡率為42%,表明5-FU能以p53非依賴性方式抑制結(jié)腸CSCs增殖,見(jiàn)表1,表2,圖1。
2.25-FU抑制CCSCs球體形成能力 CCSCs經(jīng)2.5 μg/ml 5-FU或10 μg/ml舒林酸處理后,5-FU和舒林酸均顯著抑制CCSCs球體形成,與陰性對(duì)照組相比,5-FU和舒林酸處理組球體體積顯著下降,結(jié)果表明,5-FU除了能抑制CCSCs增殖并促進(jìn)凋亡外,其還能抑制CCSCs的自我更新能力,見(jiàn)圖2。

表1 正常CCSCs與轉(zhuǎn)染shRNA-p53對(duì)細(xì)胞增殖的影響
與陰性對(duì)照組比較:1)P<0.05,下表同

表2 正常CCSCs與轉(zhuǎn)染shRNA-p53對(duì)細(xì)胞凋亡率的影響

圖1 5-FU以p53非依賴性方式抑制CCSCs增殖并誘導(dǎo)細(xì)胞凋亡(×300)

圖2 5-FU抑制CCSCs球體形成(×300)
2.35-FU促進(jìn)線粒體介導(dǎo)的凋亡途徑相關(guān)蛋白Bax/Bcl-2和細(xì)胞色素c表達(dá) 正常CCSCs和shRNA-p53-CCSCs經(jīng)5-FU處理后,細(xì)胞裂解產(chǎn)物用于Western印跡檢測(cè),5-FU處理后正常CCSCs中Bax/Bcl-2比例顯著升高,同時(shí),5-FU處理也促進(jìn)了正常CCSCs和shRNA-p53-CCSCs中細(xì)胞色素c表達(dá),表明5-FU可能通過(guò)線粒體介導(dǎo)的凋亡途徑誘導(dǎo)CCSCs凋亡,盡管舒林酸誘導(dǎo)CCSCs凋亡,但未使細(xì)胞中Bax/Bcl-2比例,明顯改變但可以促進(jìn)細(xì)胞色素c表達(dá)。見(jiàn)圖3、表3。

1~4:陰性對(duì)照組,DMSO組,舒林酸組,5-FU組,下圖同圖3 5-FU上調(diào)線粒體介導(dǎo)的細(xì)胞凋亡途徑蛋白表達(dá)
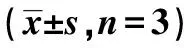
表3 5-FU上調(diào)線粒體介導(dǎo)的細(xì)胞凋亡途徑蛋白表達(dá)
與陰性對(duì)照組比較:1)P<0.01,2)P<0.001
2.45-FU抑制Wnt信號(hào)通路活化 正常CCSCs和shRNA-p53-CCSCs中,5-FU抑制胞質(zhì)和胞核中β-連環(huán)蛋白表達(dá),且其抑制效應(yīng)顯著強(qiáng)于舒林酸的抑制效應(yīng)(圖4,表4)。5-FU能夠抑制β-連環(huán)蛋白通路下游靶蛋白c-Myc和cyclinD1的表達(dá)(圖5,表5),上述結(jié)果表明5-FU能抑制β-連環(huán)蛋白的核轉(zhuǎn)位,進(jìn)而導(dǎo)致CCSCs生長(zhǎng)受到抑制。

圖4 5-FU抑制正常CCSCs和shRNA-p53-CCSCs胞質(zhì)和胞核內(nèi)β-連環(huán)蛋白表達(dá)

表4 5-FU抑制正常CCSCs和shRNA-p53-CCSCs胞質(zhì)和胞核內(nèi)β-連環(huán)蛋白表達(dá)
與陰性對(duì)照組比較:1)P<0.05,2)P<0.01,3)P<0.001,下表同

圖5 5-FU抑制正常CCSCs和shRNA-p53-CCSCs中β-連環(huán)蛋白的靶蛋白c-Myc和cyclinD1表達(dá)

表5 5-FU抑制正常CCSCs和shRNA-p53-CCSCs中β-連環(huán)蛋白的靶蛋白c-Myc和cyclinD1表達(dá)
3 討 論
本研究表明5-FU可能以非p53依賴性方式發(fā)揮作用,5-FU也能上調(diào)與線粒體介導(dǎo)的凋亡途徑相關(guān)的蛋白表達(dá),同時(shí)下調(diào)與Wnt/β-連環(huán)蛋白信號(hào)通路有關(guān)的蛋白表達(dá),5-FU的濃度為2.5 μg/ml時(shí)抑制CCSCs增殖并誘導(dǎo)細(xì)胞凋亡;然而舒林酸誘導(dǎo)的上述效應(yīng)與p53有關(guān)。以往研究表明,在AOM誘導(dǎo)的p53功能缺陷型鼠模型中,舒林酸的功能具有p53依賴性〔14~16〕,與p53+/+小鼠相比,舒林酸在p53-/-缺陷型小鼠中不能恢復(fù)急性凋亡反應(yīng),上述現(xiàn)象是至關(guān)重要的,因?yàn)樵诮Y(jié)腸癌晚期或轉(zhuǎn)移性階段,p53會(huì)發(fā)生突變〔17〕。5-FU誘導(dǎo)CCSCs發(fā)生凋亡的同時(shí),Bax/Bcl-2比例和細(xì)胞色素c的表達(dá)也升高。Bax是一種促凋亡蛋白,其能結(jié)合于Bcl-2并促進(jìn)細(xì)胞色素c釋放,細(xì)胞色素c是線粒體介導(dǎo)的凋亡途徑的重要分子〔18,19〕。這些結(jié)果表明,5-FU通過(guò)線粒體介導(dǎo)的凋亡途徑誘導(dǎo)CCSCs凋亡,本研究結(jié)果也證實(shí)5-FU抑制球體形成,因?yàn)榭寺∏蝮w的形成是干細(xì)胞干性的重要特征,本研究表明5-FU具有潛在的靶向CCSCs自我更新的潛能。本文中5-FU處理導(dǎo)致正常CCSCs和shRNA-p53-CCSCs中β-連環(huán)蛋白的表達(dá)受到抑制,β-連環(huán)蛋白的穩(wěn)定表達(dá)及其在胞核中的積累伴隨著c-Myc和cyclinD1的轉(zhuǎn)錄激活,這些基因的轉(zhuǎn)錄激活能促進(jìn)細(xì)胞增殖而增加致癌性〔20~22〕,5-FU處理后的CCSCs能抑制c-Myc和cyclinD1表達(dá),且這種抑制效應(yīng)不依賴于p53。
CCSCs的一些特征可能解釋5-FU抑制干細(xì)胞干性的原因,干細(xì)胞表達(dá)高水平的干性因子,其中包括腫瘤蛋白c-Myc〔23〕,該蛋白在CCSCs中過(guò)度表達(dá)〔24〕。本研究表明,體外5-FU能抑制CCSCs中Wnt效應(yīng)因子β-連環(huán)蛋白和下游靶蛋白c-Myc和cyclinD1表達(dá)。因此,相對(duì)于可分化細(xì)胞的致癌性變化,干細(xì)胞致癌性變化,如β-連環(huán)蛋白積累,可能對(duì)5-FU誘導(dǎo)的凋亡更敏感。
猜你喜歡
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
西南國(guó)防醫(yī)藥(2016年7期)2016-12-01 06:01:15
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年1期)2016-01-14 03:41:26
河南醫(yī)學(xué)研究(2014年3期)2014-02-27 14:51:48
沈陽(yáng)醫(yī)學(xué)院學(xué)報(bào)(2014年1期)2014-02-16 06:19:24
