Lrp家族轉錄調控因子的研究進展
2019-12-19 01:51:04柯美蘭張部昌
生物學雜志 2019年6期
關鍵詞:生物
柯美蘭, 吳 杭, 張部昌
(1. 安徽大學 生命科學學院; 2. 安徽大學 物質科學與信息技術研究院, 合肥 230601)
亮氨酸應答調控蛋白(Leucine responsive regulatory protein, Lrp)廣泛分布于細菌和古細菌中,如常見的大腸桿菌、惡臭假單胞菌和一些致病性病原菌(創傷弧菌)等;放線菌中的谷氨酸棒狀桿菌、結核分枝桿菌和抗生素產生菌等;古生菌中的嗜熱菌、硫化葉菌、甲烷球菌和嗜鹽菌等。截至目前,已報道的Lrp情況見表1。在現有真核生物基因組中,沒有發現Lrp同源蛋白[1]。
大腸桿菌Lrp(EcoLrp)能夠調控體內大約400多個基因的轉錄和表達,既能正調控涉及氨基酸生物合成、菌毛生物合成和氨同化的操縱子,也可以負調控參與氨基酸分解代謝和肽轉運[2]。大腸桿菌的天冬酰胺合成酶C(Asparagine synthetase C, AsnC)蛋白通過激活臨近天冬酰胺合成酶A基因(asnA)的轉錄和自我調節調控體內天冬酰胺的生物合成。天冬酰胺的存在可以消除這種激活作用,但并不影響AsnC的自我調節[3]。因兩者是同源的調控蛋白,且已有文獻將Lrp和AsnC蛋白放在一起描述,故又稱為Lrp/AsnC家族調控因子[4]。Lrp既可作為全局轉錄調控因子[2],也可發揮局部調控作用[3]。Lrp還能作為富足/饑餓調控蛋白[5-6],根據環境中營養的豐度調節氨基酸的代謝。當環境中營養豐富時,Lrp能降低氨基酸的合成;當處于營養貧乏的環境時,能夠刺激氨基酸的合成[2]。
隨著研究者對各族轉錄調控因子研究的進一步加深,Lrp家族得到了更多關注。本文綜述了 Lrp家族轉錄調控因子的研究進展,以期能夠為后續研究Lrp提供參考。

表1 已報道Lrp的分布情況
1 基本結構特征
Lrp蛋白單體分子量在15~18 ku之間,主要由N末端DNA結合結構域和C末端配體結合結構域組成(如圖1)[30]。DNA結合結構域能夠特異性識別并結合靶DNA,包含保守的螺旋-轉角-螺旋(HTH)結構。通過基因組序列比對發現古生菌中通過結合DNA來影響基因表達的調控蛋白大多都具有HTH結構,而且類似細菌中HTH的結構基序[31]。配體結合結構域,也稱為氨基酸代謝調控(Regulation of amino acid metabolism,RAM)結構域,可響應配體氨基酸。RAM結構域是典型的αβ-三明治結構,可以促進配體結合和蛋白寡聚化[5-6, 22, 32]。Lrp蛋白最小的功能結構單元是二聚體。兩個單體蛋白C末端結構域的β鏈相互作用,形成具有一個穩定疏水中心的二聚體蛋白,進而形成四聚體、六聚體、八聚體或十六聚體(如圖1)[33]。盡管Lrp同源蛋白之間的氨基酸序列相似性很低,但在已知Lrp晶體結構中,存在保守性的空間結構[13, 34]。
2 配體以及應答方式
2.1 Lrp的配體
最早發現EcoLrp的配體是Leu[7, 35]和應答程度較小的Ala[36-37]。隨后發現Lrp還能應答Arg、Gln、His、Lys、Met、Phe、Pro、Thr、Trp、Tyr、Val、Asp、Ile、Glu等氨基酸[19](見表2)。結核分枝桿菌RV3291c可響應Leu、Met、Ile、His和Thr[38]。腦膜炎奈瑟球菌NMB0573可應答Leu和Met[11]。嗜鹽桿菌LrpA1僅應答Asp[27]。古細菌中只有RAM結構域的DM1蛋白被證實除了對氨基酸響應外還能與2-酮戊二酸結合[5]。本實驗室研究發現Lys和Arg可以降低糖多孢紅霉菌Lrp(SACE_Lrp)對靶DNA的結合親和力,而His則增強。這表明SACE_Lrp 可以通過響應氨基酸調節蛋白對DNA的結合活性[18],這是首次被解析可以雙向響應配體的Lrp。最近發現小分子維生素B1、B3、B6、VC 等和四氫喹啉衍生物也可以作為Lrp的配體[14, 39]。
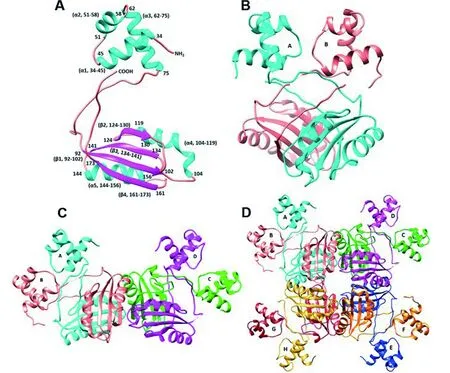
A:單體 Monomer; B:二聚體 Dimer; C:四聚體 Tetramer; D:八聚體Octamer
圖1 Lrp蛋白的結構[14]
Figure 1 Structure of Lrp protein
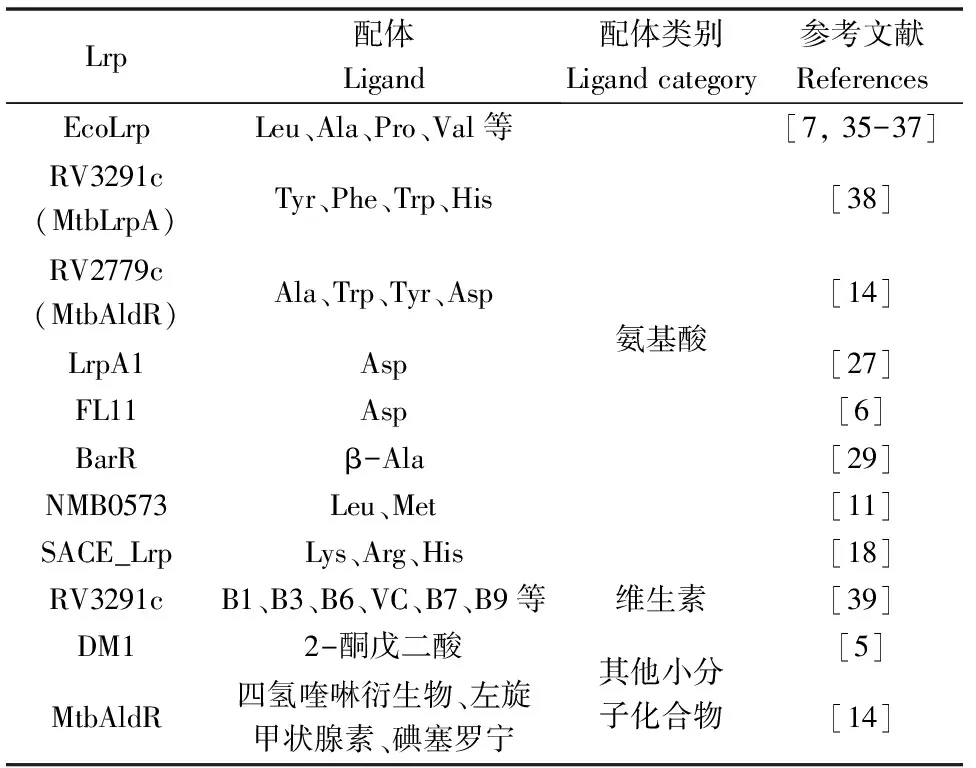
表2 Lrp應答不同種類的配體小分子
2.2 Lrp應答配體的方式
配體結合會導致Lrp蛋白構象變化[40]。Lrp在存在效應分子時,會通過結構微調來調節其DNA結合活性,從而將信號轉化成應答[38]。目前有3種變化方式:
1)DNA結合結構域附近微妙變化[41]。氨基酸結合改變了DNA結合域的相對空間分布,從而破壞了蛋白質-DNA復合物[14]。
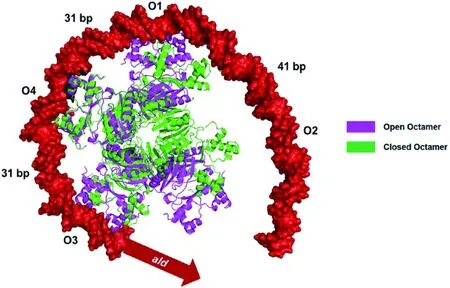
O1-O4為MtbAldR靶位點;兩個相鄰靶位點之間的數字表示位點序列中心T核苷酸(序列未列出)之間的距離
圖2“開放”和“封閉”的八聚體MtbAldR結合ald基因上游的多個靶位點[14]
Figure 2 “Open”and “closed” octamer MtbAldR binding to multiple target sites upstream ofaldgene
2)蛋白構象變化[14, 20]。“封閉”的四級結構對DNA結合域的調整或改變較小,而“開放”的結構可以使它們能夠結合到更多(20-40)堿基分隔的靶DNA位點上。如FL11與Lys結合時,形成一個“封閉”的八聚體,而與Arg結合,能使八聚體變成開環構象[20];Dey等發現MtbAldR的洗脫液中存在Ala、Asp和Phe時,蛋白質的洗脫峰會出現輕微左移,推測是因為效應分子使蛋白出現“開放”或者“閉合”的結構[14]。在透射電子顯微鏡下觀察到MtbAldR在Ala存在的情況下采用“開放”的四級結構,推測可能是因為ald基因上游存在多個靶位點需要蛋白結合[14](如圖2)。
3)蛋白寡聚體形式變化[5-6, 22, 32]。如EcoLrp與Leu結合使其從十六聚體轉變成八聚體[35];Dey等還發現MtbAldR的蛋白洗脫液中含有不同濃度的Ala時,MtbAldR以六聚體、七聚體、八聚體和十聚體等形式被洗脫下來[14]。
3 調控機制
Lrp蛋白既可以正調控靶基因的轉錄,也可以負調控靶基因的轉錄,甚至可以發揮雙重地激活或者抑制作用[2, 30, 42]。如硫磺礦硫化葉菌Ss-LrpB依據蛋白濃度雙重調節自身基因的轉錄,低濃度時促進基因的表達,高蛋白濃度則會抑制自身基因的轉錄[25, 43]。
根據已有的研究報道,Lrp調控機制總結為兩種。1)通過DNA結合結構域直接結合并調控靶基因;2)與其他調控因子或蛋白協同發揮調控作用。
大多數細菌Lrp通過直接與靶基因啟動子區結合調控靶基因的轉錄表達[11]。古細菌Lrp家族轉錄調控子的DNA結合結構域雖與細菌相似,但其轉錄機制卻類似真核生物[31],基因的轉錄起始需要轉錄起始復合物(TATA結合蛋白和其他轉錄因子)協同RNA聚合酶[44],所以古生菌中Lrp蛋白發揮調控作用不是直接由蛋白結合靶基因啟動子區而實現的。嗜熱古生菌的LrpA可以與TATA盒的下游序列結合,通過空間位阻阻斷RNA聚合酶結合轉錄起始位點,導致其無法正常地起始轉錄[25, 45]。硫磺礦硫化葉菌中的Lrs14也是通過空間位阻效應阻斷TATA結合蛋白(TATA-binding protein, TBP)、轉錄因子B(Transcription factor B, TFB)與TATA盒(TATA box)、B因子識別元件(Factor B recognition element, BRE)結合,從而發揮負向調節作用[25-26]。進一步研究還發現EcoLrp可以與其他全局性調控因子,如cAMP受體蛋白、整合宿主因子、組蛋白樣蛋白H-NS等,通過蛋白互作共同發揮調控作用[1]。有趣的是,嗜鹽桿菌中兩個Lrp可能選擇類似細菌中不同的sigma因子輔助調控不同的啟動子[27],如Lrp 激活tfbF,LrpA1 抑制tfbB。
4 調控功能
早期研究發現Lrp參與調控氨基酸的代謝,隨著研究進一步深入,發現Lrp還可調控菌毛生成、抗生素的生物合成、重金屬和多肽的轉運、ATP代謝等其他生理過程,具有多樣化的調控功能。新的研究發現Lrp還可調控致病菌的毒性強弱。以下主要介紹細菌與古生菌中Lrp的多種生物學功能。
4.1 Lrp在細菌中的調控功能
起初,Anderson等發現EcoLrp蛋白能夠影響分支氨基酸轉運[46]。隨著研究深入,發現EcoLrp不僅能影響氨基酸代謝,還能影響菌毛生成[1]。惡臭假單胞菌Lrp家族同源蛋白BkdR(與EcoLrp有36.5%的氨基酸同源性)通過正向調控bkd操縱子的轉錄表達增強BkdR對胰蛋白酶的敏感性[47]。將EcoLrp導入bkdR缺失突變株時,BkdR功能恢復,這暗示著BkdR與EcoLrp調控功能的互補性[8]。本實驗室發現SACE_Lrp與天藍色鏈霉菌中Lrp蛋白SCO3361也可以蛋白互作[16, 18]。近期發現創傷弧菌的Lrp作為全局調控因子正向調控細菌細胞毒力、趨化性和鐵離子獲取能力。Lrp可以結合在7個毒力相關基因的啟動子區并發揮正調控作用,但本身進行負向自我調節。當lrp突變后,突變株中與鐵轉運、蜂毒肽合成相關基因的轉錄水平被下調,與細胞毒素分泌、趨化性相關的基因水平都顯著降低[9]。嗜線蟲致病桿菌中全局性調控子Lrp蛋白的豐度會導致細菌的毒力發生變化[10]。
近年來,放線菌Lrp也得到較多的關注。谷氨酸棒狀桿菌Lrp通過激活轉運蛋白BrnFE的表達提高L-異亮氨酸的產量[12, 48]。結核分枝桿菌LrpA(RV3291c) 可以幫助細菌抵抗營養貧乏等不利的環境。當營養不足時,lrpA的轉錄水平升高,且LrpA會直接結合其上游編碼賴氨酸氨基轉移酶基因lat的啟動子區,促進lat轉錄[13]。MtbAldR(RV2779c)也具有同樣的調控作用,直接結合其上游編碼丙氨酸脫氫酶的靶基因(ald)。在缺乏營養的條件下,ald的轉錄水平會高度上調[14]。天藍色鏈霉菌中Lrp蛋白BkdR可以調控相鄰bkd操縱子的表達。當bkd突變后,菌株會存在產孢缺陷并且失去有關色素的表達能力,致使孢子數量減少、活力降低并且放線紫紅素的產量明顯下降[49]。Yu等還發現天藍色鏈霉菌中Lrp同源蛋白SCO2140可以正向調控放線紫紅素和鈣依賴抗生素的生物合成。SCO2140缺失突變后,孢子形成時間會延遲,而SCO2140蛋白缺乏DNA結合結構域,其作用靶點并不清楚[15]。
本實驗室在Lrp對抗生素生物合成的基因調控研究領域取得重要進展,發現SACE_Lrp通過調控分支氨基酸(Leu,Ile和Val)的轉運和代謝,負向影響紅霉素生物合成。這是首次在產抗生素放線菌中解析Lrp對抗生素生物合成調控機制的研究[18]。此后我們還發現天藍色鏈霉菌SCO3361通過直接調控actII-ORF4(編碼放線紫紅素生物合成基因簇內調控蛋白)和amfC(編碼氣生菌絲形成蛋白)的轉錄水平,正向影響放線紫紅素的生物合成和菌株的形態分化[16]。最近,Lu等又發現螺旋霉素鏈霉菌中的Orf2(SSP_Lrp)是一個全局性調控因子,通過控制(雙)螺旋霉素生物合成基因簇內正調控基因bsm23、bsm42和acyB2的轉錄水平,負調控(雙)螺旋霉素的生物合成[17]。
4.2 Lrp在古生菌中的調控功能
Lrp/AsnC家族是古生菌中4個廣泛存在并且數量最多的轉錄調控蛋白家族之一[31],其余3個調控家族分別為:HTH_3(假定激活蛋白家族)、TrmB(麥芽糖特異性調節相關蛋白)和ArsR(解毒過程相關蛋白)。目前古生菌中Lrp的研究主要集中在4個模式菌株:嗜熱菌、硫化葉菌、甲烷球菌和嗜鹽菌[19]。
古生菌Lrp不僅能調節氨基酸代謝,還可影響中心代謝、ATP代謝、重金屬多肽等物質運輸。堀崎嗜熱球菌Lrp家族蛋白FL11可應答Lys[20],還可以作為“富足或饑餓”調節因子調控細胞的生長狀態[6]。當營養富足時,氨基酸分解代謝并且ATP合成處于激活去阻遏狀態,細胞生長;當營養貧乏時,氨基酸生物合成基因被激活,細胞停止生長[6]。嗜鹽桿菌Lrp通過激活korAB(編碼氧戊二酸鐵氧還蛋白氧化還原酶)影響氧戊二酸和谷氨酸的平衡控制TCA循環的方向和速率,進而參與調控中心代謝[27];可正調控編碼谷氨酰胺合成酶的glnA、轉錄調節因子sirR和負調控甘油脫氫酶基因gldA1及轉導基因car;還可以刺激磷酸鹽轉運操縱子pstC2和phnC的表達[27]。另一Lrp蛋白(LrpA1)依賴L-天冬氨酸特異性地負向調節編碼天冬氨酸轉氨酶的aspb3基因,但正向調節自身基因的轉錄水平。詹氏甲烷球菌Ptr2通過二級上游激活位點(UAS)向TATA盒募集TBP增強紅素氧還蛋白Rb2和鐵氧還蛋白FdxA的表達[25, 28]。來自硫磺礦硫化葉菌的Ss-LrpB可正向激活丙酮酸鐵氧化還原酶操縱子porDAB和兩個滲透酶基因表達,這初步鑒定了Ss-LrpB蛋白的體內生理作用[25, 32]。嗜酸熱硫化葉菌BarR激活參與β-Ala降解的β-丙氨酸轉氨酶基因表達,也激活編碼谷氨酰胺合成酶基因的轉錄[29]。
古生菌中一些Lrp蛋白不僅可以調節本家族轉錄調控因子的表達,還可以調控其他家族調控子的表達,暗示著其中可能存在分級調控網絡[19, 27]。如堀崎嗜熱球菌的全局性調控因子FL11可以影響至少其他4個Lrp蛋白的表達[19-20]。
5 展望
近年來,Lrp受到越來越多地關注,但相對其他轉錄調控蛋白家族的研究,如TetR、LuxR和MarR等[50],其研究還是較少。大腸桿菌Lrp的結構和功能已研究得較為透徹,這對理解來自不同種屬Lrp的調控功能具有指導意義。然而,仍有許多問題亟待解決。例如:尚未有文章詳細介紹Lrp N末端的保守氨基酸如何識別不同的結合位點,這對研究Lrp如何發揮調控作用具有重要意義;古生菌中Lrp靶基因和效應分子仍缺乏認識,其分級調控網絡仍不清楚;古菌中DM1蛋白和天藍色鏈霉菌中SCO2140蛋白缺乏DNA結合結構域,其是怎樣發揮調控作用并不清楚;目前僅有一篇報道從結構上解釋了氨基酸(僅Ala)如何影響Lrp(MtbAldR)的DNA結合活性。這為后續研究其他Lrp如何應答氨基酸提供依據。文章還提出,不同配體的結合是否會引起蛋白質四級結構細微的變化而形成不同程度“開放”,不同四級結構的蛋白與靶位點結合是否引發不同的應答,都需要進一步研究。此外,本實驗室解析了Lrp參與調控抗生素生物合成的分子機制[16, 18],這為研究其他產抗生素放線菌中的Lrp指明了方向[17]。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
