模式原生動物浮萍棘尾蟲純培養體系的建立與優化
2019-11-26 02:17:30吳禹岐李世寬范金鳳
水生生物學報 2019年6期
王 影 姚 琳 吳禹岐 李世寬 范金鳳 陳 瑛
(哈爾濱師范大學黑龍江省水生生物多樣性研究重點實驗室, 哈爾濱 150025)
浮萍棘尾蟲(Stylonychia lemnae)是一種營自由生的淡水纖毛蟲, 廣泛分布于池塘、河流和沼澤中。隸屬于原生動物界纖毛門多膜綱下毛目尖毛科[1], 是棘尾蟲屬的模式種。作為單細胞模式生物, 棘尾蟲在研究核組織和發育、染色體和染色質、端粒生物學以及基因組重排等領域有著重要的地位[2—4]。早在1876年, 第一份記錄染色體和有絲分裂紡錘體的報道就來自于對棘尾蟲小核結構的觀察[5]。另外一份研究發現, 在浮萍棘尾蟲大核多倍體化后, 基因組中有大量DNA(約90%)存在丟失現象, 這一發現引起了學者們對于纖毛蟲中基因組減少和重排現象的廣泛研究。隨后, 浮萍棘尾蟲即作為模式單細胞生物, 廣泛用于端粒結構調控[6]和染色質動力學的研究中[7—9]。在2013年,浮萍棘尾蟲大核基因組完成測序, 為研究染色質介導、RNA引導的發育基因組重排等現象提供了分子生物學基礎[10]。基于基因組測序結果, 本實驗室研究了浮萍棘尾蟲在表皮生長因子誘導下的轉錄組響應, 從轉錄水平初步揭示了其分裂和增殖調控的分子機制[11]。棘尾蟲主要營動物性營養[12],傳統的棘尾蟲培養方式是喂飼藻類(Algae)、細菌(Bacteria)及唇鞭蟲(Chilomonas)等。在利用該物種作為研究對象開展相關實驗前, 通常需進行饑餓處理, 以減少細菌和微藻對研究結果的干擾, 但是依然難以完全消化蟲體內的食物[13], 非常不利于開展棘尾蟲的基礎生物學研究。因此, 建立浮萍棘尾蟲純培養體系, 成為開展棘尾蟲生物學研究的重要前提。
原生動物的培養方式主要分為自由生和寄生型兩類: 自由生活的模式原生動物以草履蟲(Paramecium)和四膜蟲(Tetrahymena)為代表, 其專用的無菌培養基已經建立[14—16], 其他種屬的培養方式均為混合培養[17—19]。因此, 本研究以原生動物嗜熱四膜蟲純培養基為主要參考[20], 采用響應面分析方法,通過單因素實驗和正交實驗篩選調整各個主要組成分配比, 以獲得適合浮萍棘尾蟲細胞的無菌純培養基, 并進一步優化了培養條件。
1 材料與方法
1.1 供試材料
實驗所用的浮萍棘尾蟲(Stylonychia lemnae)由中國科學院水生生物研究所饋贈。其基因組數據庫網址: http://stylo.ciliate.org/index.php/home/welcom。
1.2 培養基及實驗方法
嗜熱四膜蟲培養基的成分主要包括碳源、氮源、無機鹽、葡萄糖和檸檬酸等成分達39種, 用于探究不同成分對蟲體生長繁殖的影響, 具體成分參考唐任寰等[21]; 培養唇鞭蟲的Pringsheim’s液成分為Na3PO4、KCl、Ca(NO3)2、MgSO4和FeSO4; 綠梭藻(Chlorohonium)培養基成分為NaCl、KCl、CaCl2、MgCl2、NaH2PO4、NH4NO3、MgSO4、FeCl3和MnCl2等無機成分, 具體成分參考Ammermann等[22], Pringsheim’s液、綠梭藻培養基只用來比較和檢驗優化培養基的優化效果。
優化培養基的方法主要分為兩步: 第一步, 利用單因素實驗設計, 探索主要營養元素的濃度, 為優化培養基成分提供合理的數據范圍。第二步, 為克服單因素實驗的局限性, 通過正交實驗和統計學數據分析后, 確定主要影響因素。將最佳培養基成分和最適培養條件相組合, 篩選出適合于浮萍棘尾蟲的培養體系。
1.3 浮萍棘尾蟲培養方法
取100只蟲體放入10 mL的培養液中, 在設定的pH和溫度條件下靜置培養, 每24h測定一次細胞密度, 每次計數進行3個技術重復。
1.4 氮源濃度對浮萍棘尾蟲生長的影響
在嗜熱四膜蟲培養基其他組分不變的條件下:分別添加甲硫氨酸、蘇氨酸、纈氨酸、異亮氨酸、組氨酸、丙氨酸、谷氨酸、谷氨酰胺、甘氨酸、絲氨酸(以5%的單位為一個濃度梯度, 共稀釋7個濃度), 研究10因素不同濃度組合條件下氮源濃度對細胞密度的影響。每個處理做3個重復。培養方法同1.3。
1.5 碳源濃度對浮萍棘尾蟲生長的影響
在嗜熱四膜蟲培養基其他組分不變的條件下:分別添加葡萄糖和檸檬酸(以10 mg/L為一個濃度梯度, 共稀釋6個濃度), 研究在不同濃度條件下碳源濃度對細胞密度的影響。每個處理做3個重復。培養方法同1.3。
1.6 磷酸氫二鉀濃度對浮萍棘尾蟲生長的影響
在嗜熱四膜蟲培養基其他組分不變的條件下:分別添加磷酸氫二鉀(以0.1%的單位為一個濃度梯度, 共稀釋5個濃度), 研究磷酸氫二鉀濃度對細胞密度的影響。每個處理做3個重復。培養方法同1.3。
1.7 優化試驗
響應面優化實驗響應面優化實驗是由Box等[23]提出的一種試驗設計方法, 是一種綜合試驗設計和數學建模的優化方法, 通過對具有代表性的局部各點進行試驗, 回歸擬合在全局范圍內因素與結果之間的函數關系, 并且取得各因素的最優水平值[24]。氮源采用central composite design設計了10因素的組合實驗, 研究不同組合條件對于棘尾蟲細胞數目的影響, 篩選出最適的氮源元素含量的配比。
正交實驗在單因素實驗結果的基礎上, 以氮源、碳源的母液(葡萄糖和檸檬酸按照1:1配制)和磷酸氫二鉀作為考察因素, 按正交實驗設計,統計實驗結果, 比較各組合培養條件對細胞密度的影響, 篩選出最適于棘尾蟲生長的培養基組分。每個處理做3個重復。培養方法同1.3。
1.8 最佳培養條件的篩選
在優化后的培養基下培養, 研究培養溫度(10、15、20、25和30℃)、培養基初始pH (6.0、6.5、7.0、7.5和8.0)和初始接種密度(5、50、100、200和300 cells/mL)對最終獲得細胞密度的影響。每個處理做3個重復。培養方法同1.3。
1.9 細胞密度與代時計算方法
生長曲線與代時和細胞密度的換算公式如下:

其中, A為初始體系中微生物數量常用對數值;B為最大生長速率; C為微生物穩定期數量常用對數值與初始數量常用對數值之差; M為最大生長速率時所對應的時刻;Y(Nt)為微生物在時間t時的常用對數值lg。根據上述公式用Origin8.0即可直接得出參數A、C、B和M, 進而算出: MCC=eA+C;GT=(2.7182/B)/(log2MCC/N0); 其中, e為自然對數,N0為接種初始密度, MCC為細胞最大密度, GT為對數期的代時[25]。通過生長曲線可算出最大細胞密度(Maximum cell concentration, MCC)和代時(Generation times, GT)。
1.10 數據統計
實驗數據利用SPSS(20.0)進行統計學分析。
2 結果
2.1 氮源濃度對細胞密度的影響
單因素實驗發現, 嗜熱四膜蟲培養基中18種氨基酸里有10種氨基酸的濃度變化對細胞密度具有顯著影響。利用central composite design設計了10因素的組合實驗, 共300組不同濃度氨基酸的組合實驗。經過方差分析和各因素的貢獻度分析得出: 呈現正效應的異亮氨酸貢獻度最大為48.182%、丙氨酸的貢獻度是22.4506%, 呈現負效應的組氨酸的貢獻度是14.6905%, 說明在一定的范圍內, 增加異亮氨酸、丙氨酸的濃度含量并減少組氨酸的濃度含量有助于棘尾蟲細胞密度的增加。氮源中心組合的細胞密度的回歸方程為:
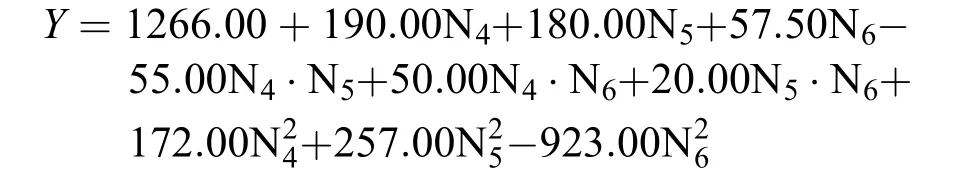
N4為異亮氨酸, N5為組氨酸, N6為丙氨酸。回歸方程用決定系數R2檢測, 其值為0.9037, 說明實際值與預測值擬合良好。變異系數(CV)=5.18%, 值稍大, 但在合理范圍內, 說明模型可信度高。通過響應面的貢獻度分析發現N4-6的貢獻程度最大, 需要進一步調整比例, 在實際統計數據結果時發現當組氨酸的添加量為零時, 細胞生長密度最高, 所以最終表 1中只包含9種氨基酸, 配成氮源母液。氮源的配比見表 1。
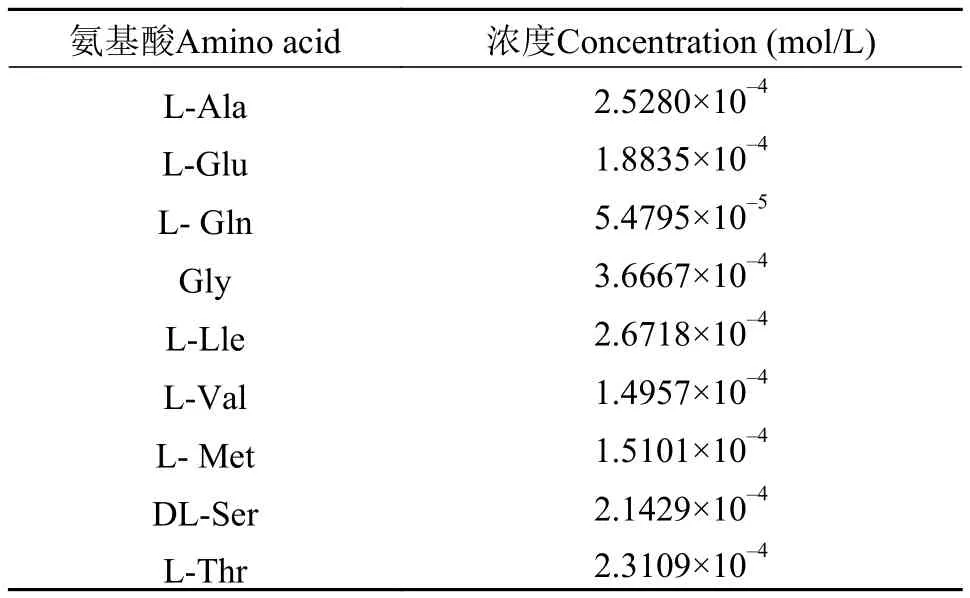
氨基酸Amino acid 濃度Concentration (mol/L)L-Ala 2.5280×10-4 L-Glu 1.8835×10-4 L- Gln 5.4795×10-5 Gly 3.6667×10-4 L-Lle 2.6718×10-4 L-Val 1.4957×10-4 L- Met 1.5101×10-4 DL-Ser 2.1429×10-4 L-Thr 2.3109×10-4
2.2 碳源濃度對細胞密度的影響
培養基中的碳源是由葡萄糖和檸檬酸提供, 因此對碳源進行單因素實驗, 結果如圖 1所示, 當葡萄糖濃度為30 mg/L、檸檬酸濃度為40 mg/L, 作為碳源的最佳濃度時, 細胞密度均存在最大值約為400 cells/mL。
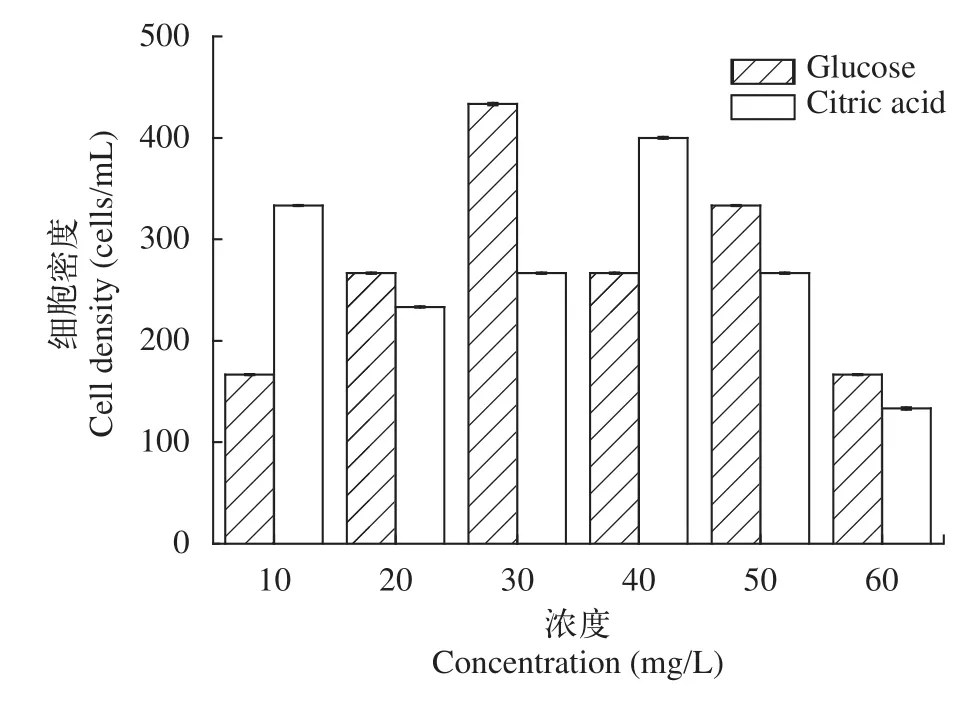
圖1 葡萄糖和檸檬酸的濃度對細胞密度的影響Fig. 1 The effect of glucose and citric acid on cell density
2.3 磷酸氫二鉀濃度對細胞密度的影響
如圖 2所示, 細胞密度隨著K2HPO4·3H2O濃度的增加而增加。當K2HPO4·3H2O的濃度達到0.5%時, 細胞密度達到最大300 cells/mL左右, 而K2HPO4·3H2O濃度繼續提高細胞密度則隨之遞減。在一般情況下, K2HPO4·3H2O除了為細胞的生長提供磷源外, 還具有穩定培養液pH的作用, 偏高或偏低的pH都不利于細胞生長。本實驗驗證了K2HPO4·3H2O的最適添加量為0.5%。
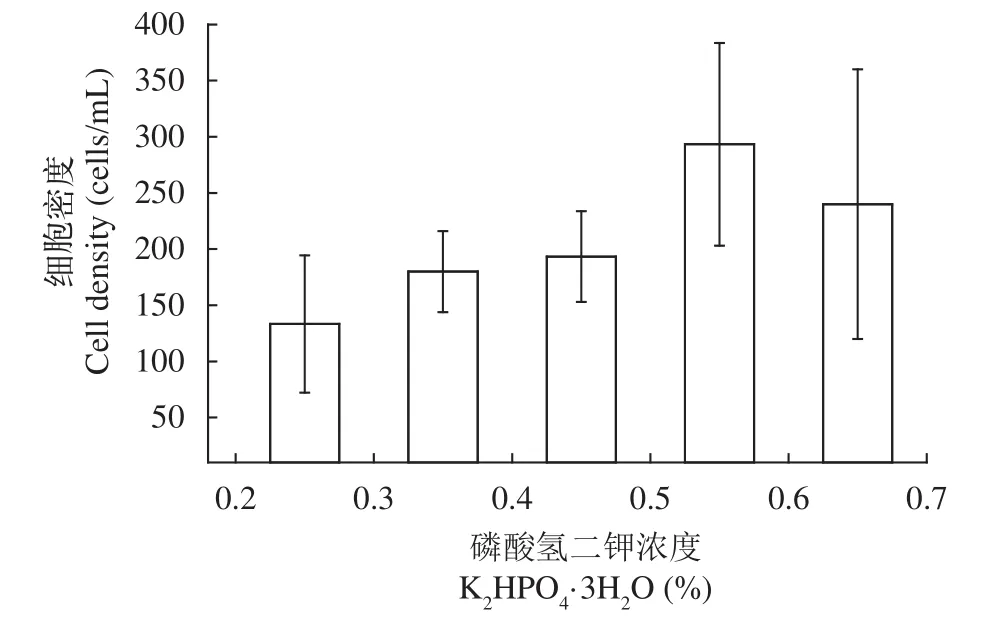
圖2 磷酸氫二鉀的濃度對于細胞密度的影響Fig. 2 The Effect of K2HPO4 on cell density
2.4 氮源、碳源和磷酸氫二鉀三因素的正交實驗及結果分析
為克服單因素實驗結果的局限性, 探究幾大營養元素之間相互作用的關系, 分別以氮源母液、碳源母液及磷酸氫二鉀作為三大類營養元素, 采用正交實驗設計方法, 設計了3因素3水平的優化實驗,正交實驗設計見表 2, 結果分析見表 3。
三因素中心組合的細胞密度的回歸方程為:
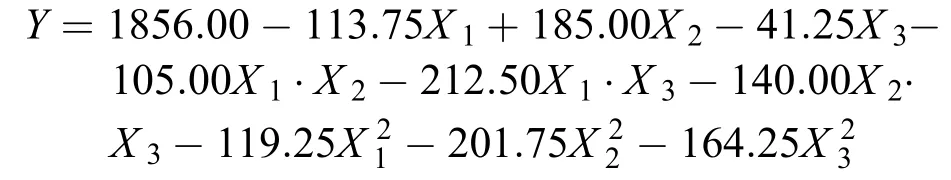
X1:氮源,X2:碳源,X3:磷酸氫二鉀, 回歸方程用決定系數R2檢測, 其值為0.8221, 說明實際值與預測值擬合良好; 變異系數(CV)=11.21%, 值稍大, 但在合理范圍內, 說明模型可信度高, 因素和水平的顯著性可有效地檢測棘尾蟲的生長情況, 反映因素之間相互影響的關系。實驗數據經過方差分析, 從表中可以觀察到氮源、碳源和磷酸氫二鉀對于棘尾蟲細胞生長的影響都顯著, 氮源的影響最大, 其次是磷酸氫二鉀。最佳的配比為氮源的濃度80 mg/mL, K2HPO4·3H2O的濃度達到0.65% mg/mL, 碳源的終濃度為20 mg/mL時, 細胞密度最大可達868 cells/mL左右。
2.5 優化純培養基與常用培養基對浮萍棘尾蟲培養效果的比較
在相同的培養條件下, 初始接種量均為10 cells/mL, 分別添加等量的綠梭藻培養液、Pringsheim’s唇鞭蟲培養液和純培養基, 每組設置3個重復, 對其進行長達144h的計數觀察, 統計數據通過差異顯著性分析, 得到3種培養基對于浮萍棘尾蟲的影響, 得到在72h、96h和120h時均差異性顯著,P<0.05, 說明優化后的培養基, 相對于其他兩種培養條件, 具有顯著影響。
在相同培養條件下, 浮萍棘尾蟲在Pringsheim’s液、綠梭藻培養液和優化培養基(Medium)中的生長曲線見圖 3。可以看出在3種培養基中, 細胞密度都在120h左右達到最大, 120h后隨之減少。在優化培養基中最大細胞密度(Maximum cell concentration)MCC約為6.0×102cells/mL、代時(Generation times)GT為10h, 而在常用的Pringsheim’s液和綠梭藻使用的混合培養基中MCC為4.8×102和4.0×102cells/mL、GT為13h。在優化后培養基相對于傳統的培養方式, 在保證細胞數量有所增加的情況下, 成功的縮短了細胞分裂代時。同時, 活體和蛋白銀染色觀察培養蟲體效果(圖 4), 發現無菌純培養基中生長的棘尾蟲體內不再有大量藻和細菌, 細胞質呈現均勻潔凈的狀態。
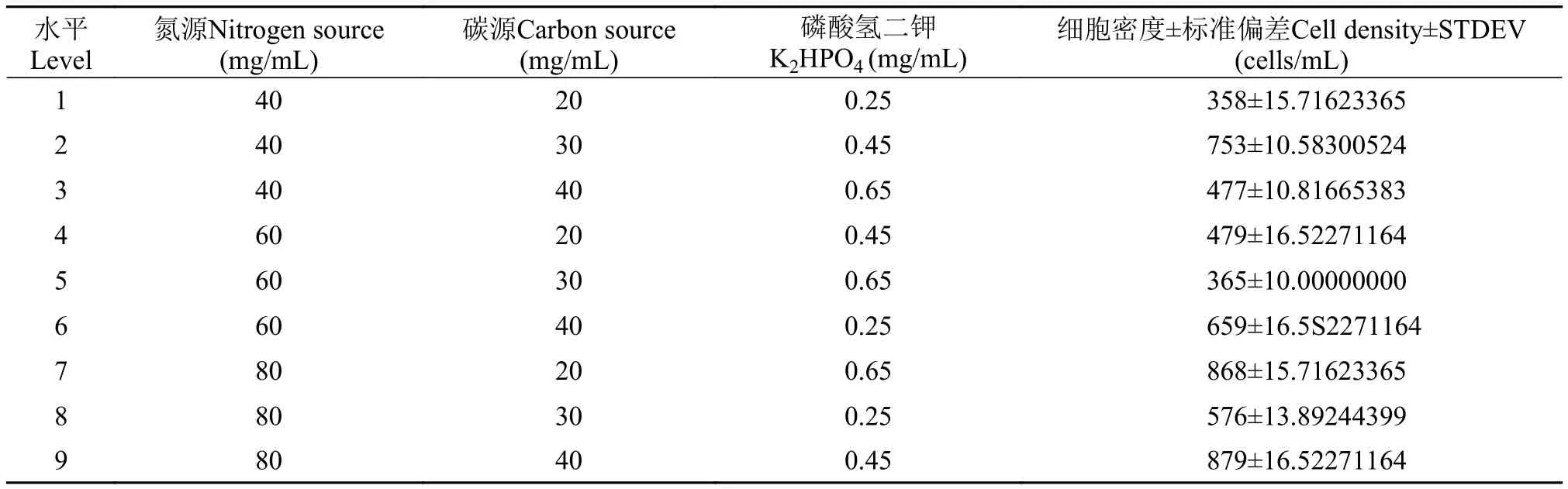
表2 三因素的正交實驗設計Tab. 2 Three-factor orthogonal experimental design
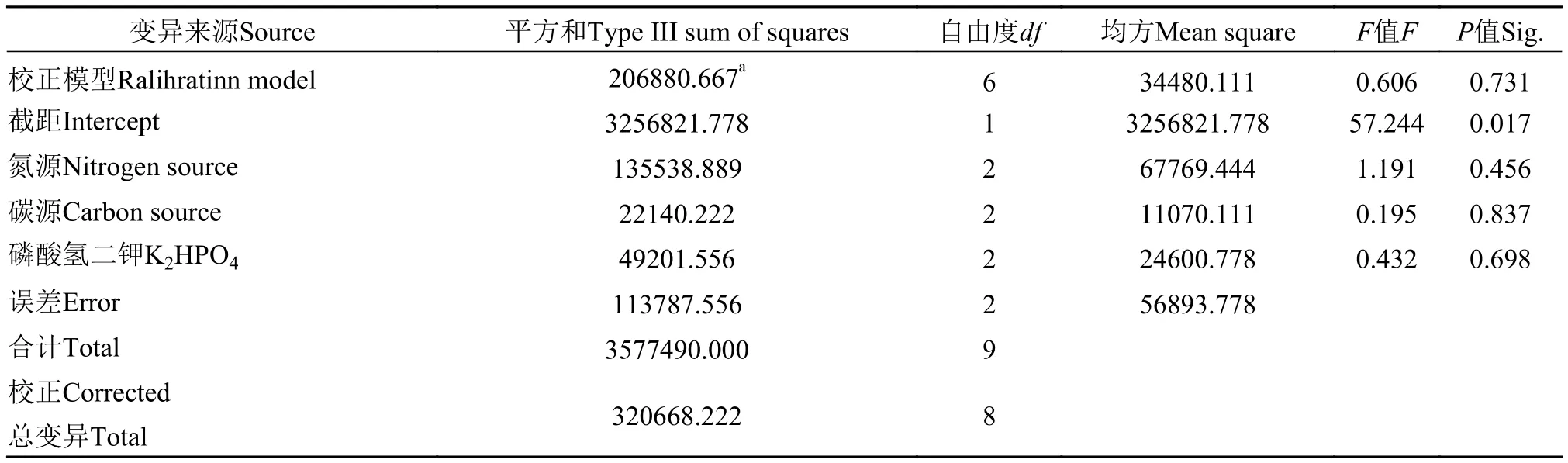
表3 三因素的方差分析Tab. 3 ANOVA for content of three components
2.6 浮萍棘尾蟲培養條件的優化
分別以培養溫度、初始pH、接種量做單因素實驗, 根據培養結果, 分析可知: (1)溫度對于棘尾蟲生長的影響是顯著的, 隨著溫度的升高, 細胞密度逐漸增大, 到25℃時, 細胞密度可達到850 cells/mL左右; 但溫度進一步升高, 蟲體密度急劇減少(圖 5);(2)不同的初始pH對棘尾蟲的生長具有一定的影響,隨著初始pH的增加, 細胞密度也逐漸增加, 當初始pH為7.0時, 細胞密度達到1100 cells/mL左右, 當初始pH繼續增加時, 細胞密度反而下降, 利用單因素實驗得出, 棘尾蟲生長的最佳初始pH為7.0 (圖 6);(3)不同的接種量對棘尾蟲數量的影響是顯著的, 利用單因素實驗得出, 當接種量為100 cells/mL時, 細胞密度可達到3500 cells/mL, 在實驗過程中發現過高或過低的接種量均無法滿足分子實驗的需求, 確定最佳接種量為100 cells/mL (圖 7)。本實驗中采用的是容量為50 mL的無菌培養瓶, 加入10 mL培養液后水平放置, 既能夠保證培養瓶內有充足的氧氣, 又能給棘尾蟲類喜貼壁生活的原生動物提供足夠的表面積, 因此選擇的裝液量為10 mL。當溫度為25℃, 初始pH為7.0, 接種量為100 cells/mL時, 此為各因素的最佳培養條件。
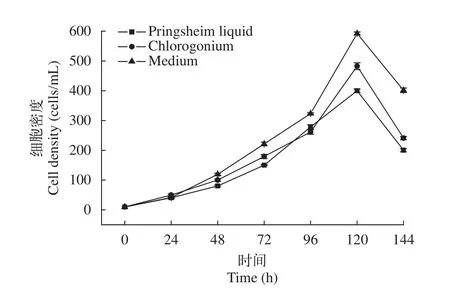
圖3 浮萍棘尾蟲在三種培養條件下的生長曲線Fig. 3 The growth curve of Stylonychia lemnae under three culture conditions
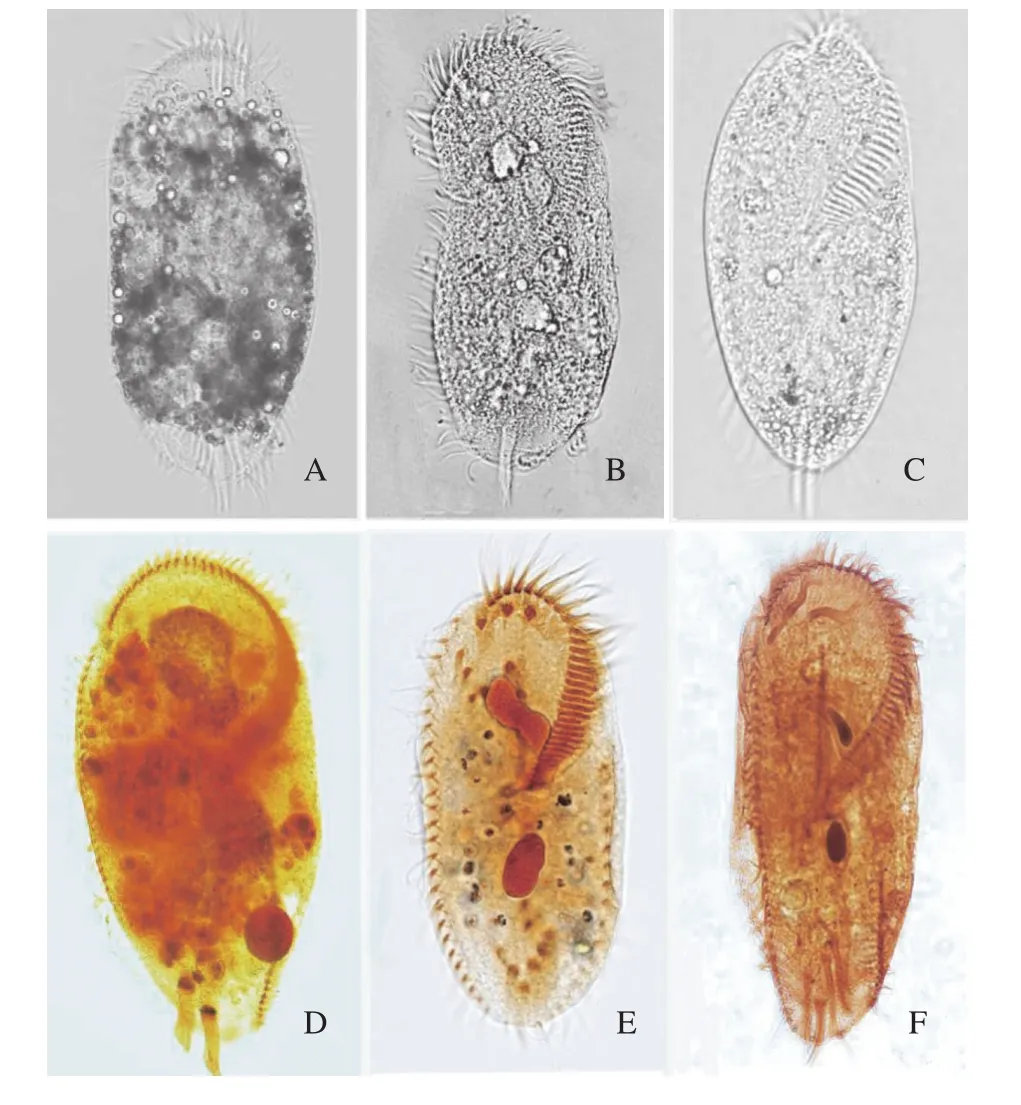
圖4 浮萍棘尾蟲在不同培養條件下的活體和染色圖片Fig. 4 Living and staining images of Stylonychia lemnae under different culture conditions
2.7 培養基配制方法及浮萍棘尾蟲的馴化
本實驗將無菌培養浮萍棘尾蟲的純培養基命名為SLPP, 具體成分配比如下(表 4), 具體配置方法和步驟: (1)按配方稱取相應藥品, 溶于100 mL滅菌ddH2O, 攪拌溶解均勻后, 調節pH等于7.0, 配制為母液; (2)將母液高壓蒸汽滅菌鍋, 115℃, 滅菌15min, 冷卻后4℃冰箱儲存待用; (3)培養前, 在超凈工作臺中, 用滅菌ddH2O將母液稀釋20倍后, 用于培養蟲體。
為了避免藻類的干擾, 浮萍棘尾蟲可先選用普通麥粒水培養一段時間, 待達到對數生長期后, 再將浮萍棘尾蟲轉接到滅菌的SLPP培養基中, 在25℃條件下, 無菌培養至對數期生長期, 再連續轉接3次, 可獲得能夠穩定的在SLPP培養基中生長繁殖的無菌蟲體, 用于后續的擴大培養和分子實驗。

圖5 浮萍棘尾蟲在不同溫度下的細胞密度Fig. 5 The cell density of Stylonychia lemnae under different temperatures
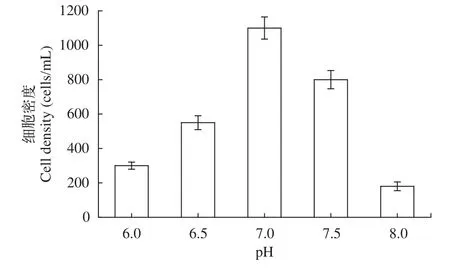
圖6 浮萍棘尾蟲在不同初始pH下的細胞密度Fig. 6 The cell density of Stylonychia lemnae under different initial pH
3 討論
原生動物大都生活在有機質豐富、水質腐敗且不大流動的污水中。不同的水質, 生活著不同種類的原生動物。通常我們在野外采集回的樣品, 均需經過原位水的培養, 逐漸替換為蒸餾水進行馴化,培養過程中通過添加麥粒、米粒等物質產生的菌類, 供給營養[26]。研究發現原生動物和水細菌關系密切, Sanders等[27]指出無論在淡水或海洋生態系統中, 細菌和鞭毛蟲的數量有著較為穩定的比例(約1000:1)。原生動物對細菌的掠食速度是驚人的,每時每刻都在進食, 所以食用的菌類、藻類等物質長期殘留在蟲體內。在顯微鏡下分別拍攝在菌類、藻類喂飼下的棘尾蟲活體和蛋白銀染色圖片時, 可明顯的觀察到存在于蟲體內部未消化完全的食物(圖 4)。因此, 為了減少細菌和藻類對分子測序分析的干擾, 往往在提取浮萍棘尾蟲遺傳物質前,先進行饑餓處理, 但是細菌和微藻在蟲體內依然難以被完全消化[28], 對分子測序結果產生干擾。而使用已有的自由生纖毛蟲無菌純培養基時, 發現棘尾蟲不能獲得穩定的生長效果。經過本實驗的研究和檢驗, 初步建立了適合浮萍棘尾蟲無菌純培養的培養基和培養條件。利用該純培養體系, 最大細胞密度可比傳統培養方式提高1.2%左右, 有效避免其他物種干擾, 同時, 該培養基成分與嗜熱四膜蟲純培養基相比更加簡潔廉價。
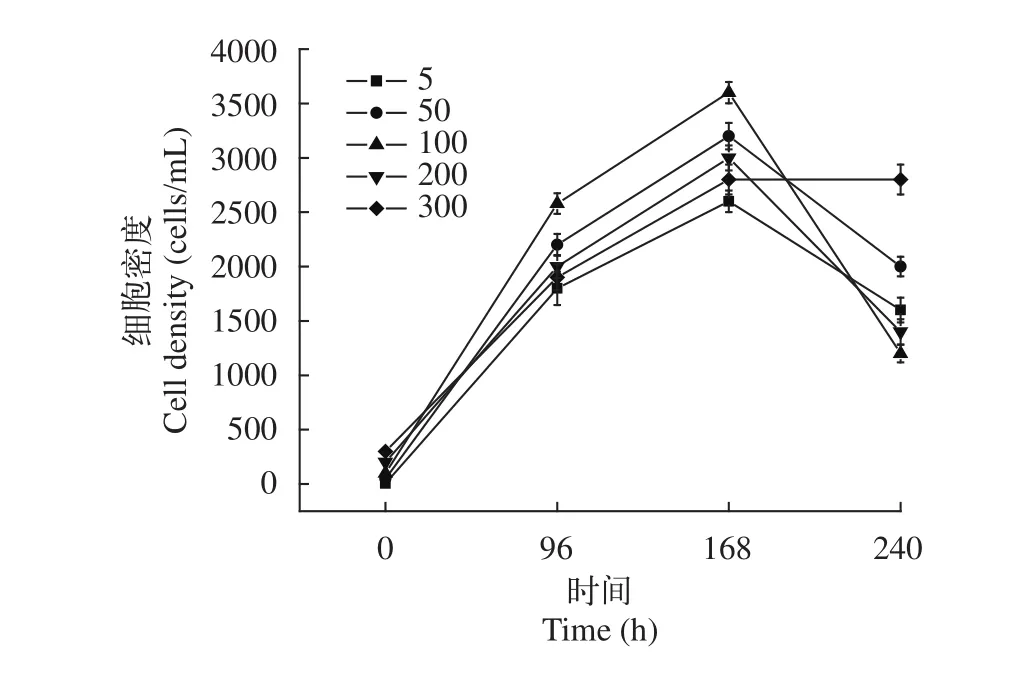
圖7 浮萍棘尾蟲在不同接種量下的細胞密度Fig. 7 The cell density of Stylonychia lemnae under different inoculation quantity
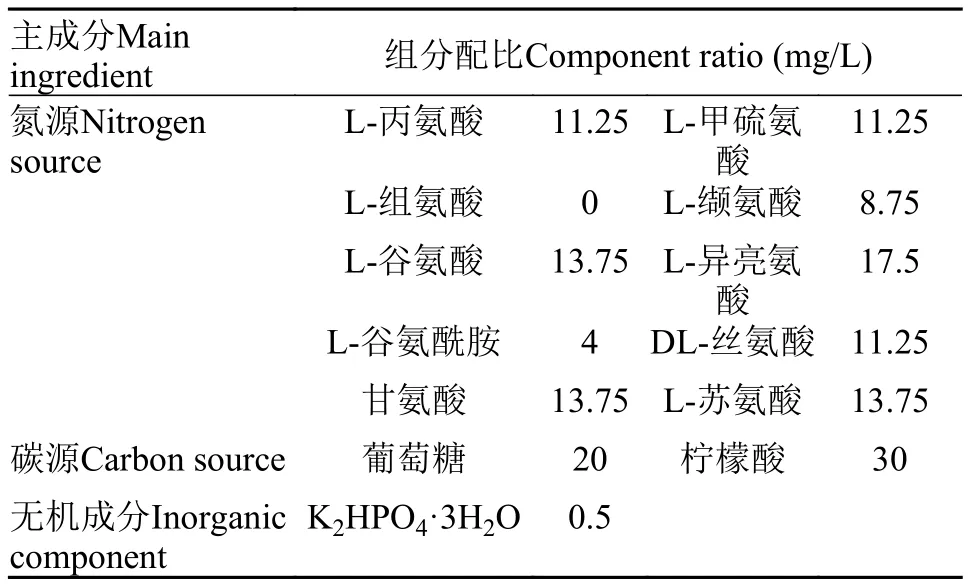
表4 SLPP培養基組分Tab. 4 SLPP medium component
經過優化的培養基從原來的“少量原種, 純營養液, 長周期培養”的方式轉變為“大量原種, 配合型營養液, 短周期培養”的方式[29], 在培養基中所添加的碳源、氮源和無機鹽均是原生動物培養中常用的營養成分。經過篩選營養成分和優化培養條件, 建立起了適用于腹毛類棘尾蟲屬的培養基, 既可以獲得純凈的生物樣品, 又可以縮短實驗材料培養的周期, 有利于后續實驗的開展。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
房地產導刊(2022年5期)2022-06-01 06:20:14
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
中學生數理化(高中版.高考數學)(2021年12期)2021-03-08 01:28:50
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
